
M. Saraswathi
Department of Applied Microbiology, Sri Padmavati Mahila VisvaVidyalayam, Tirupati, A.P., India
M.N. Reddy
Department of Applied Microbiology, Sri Padmavati Mahila VisvaVidyalayam, Tirupati, A.P., India
International Journal of Plant Pathology
Year: 2012 | Volume: 3 | Issue: 2 | Page No.: 82-88







ABSTRACT
Stem rot caused by Sclerotium rolfsii is a potential threat to groundnut yield. This disease causes severe damage during any stage of crop growth and yield losses over 25%. Isolation trails from diseased plants were carried out in some areas of Srikalahasti and Tirupati of Chittoor district. Oatmeal-sand culture of S. rolfsii was inoculated to 10 day old seedlings of TMV-2 variety of groundnut for disease development and divided into five fairly distinct stages, on the basis of lesion development. Phenolic extracts were prepared from the hypocotyl regions with lesions and healthy seedlings respectively. The extracts were subjected to 2-dimensional ascending paper chromatographic technique by employing the benzene: acetic acid: water as first solvent system and sodium formate:formic acid:water as the second solvent system. There is a tremendous increase in qualitative and quantitative production of phenolic acids in the diseased area of the groundnut hypocotyls compared to healthy ones. Chromatographic data revealed that there are 10 phenolic acids in healthy plants, whereas in the infected plants, in addition to the above 10 phenolic acids, 6 additional compounds were detected with the progression of the disease.
PDF Abstract XML References Citation
Received: November 05, 2011;
Accepted: March 21, 2012;
Published: June 05, 2012
How to cite this article
M. Saraswathi and M.N. Reddy, 2012. Phenolic Acids Associated with Sclerotium rolfsii in Groundnut
(Arachis hypogaea L.) During Pathogenesis. International Journal of Plant Pathology, 3: 82-88.
DOI: 10.3923/ijpp.2012.82.88
URL: https://scialert.net/abstract/?doi=ijpp.2012.82.88
DOI: 10.3923/ijpp.2012.82.88
URL: https://scialert.net/abstract/?doi=ijpp.2012.82.88
INTRODUCTION
Sclerotium rolfsii is a ubiquitous soil-borne fungal pathogen known to cause disease on worldwide range of agricultural and horticultural crops (Kaveriappa, 1979; Sennoi et al., 2010). Among plantae, more than 500 species belonging to 100 families are susceptible to this pathogen (Punja, 1985; Sarma et al., 2002; Adandonon et al., 2005; Pandey et al., 2005; Ganesan et al., 2007). It has been reported that most of S. rolfsii infections were identified in dicots and also in some monocots (Aycock, 1966). Patil and Rane (1983) reported that all the 35 hosts, including important cultivated crop plants, were susceptible to the pathogen, indicating the wide host range of parasitism of S. rolfsii. Numerous secondary hosts were identified to this pathogen, among which food crops and ornamental plants that are economically important are included. Secondary hosts also include a variety of hosts.
S. rolfsii devastates the plant severely at any stage of crop its growth and is detrimental to all parts of the plant, but stem damage is the most common and serious risk (Ganesan et al., 2007). As a protective structure, sclerotia possess viable hyphae which serves a primary inoculums for disease development (Aycock, 1966; Nalim et al., 1995; Cilliers et al., 2000) as well as its principle means of dispersal and sole organs by which the fungus survives adverse environmental condition, germinates and infects the susceptible hosts during favourable conditions. The disease manifests itself in the form of dark sunken necrotic lesions girdling the hypocotyls near soil level. The necrotic lesions formed due to infection with S. rolfsii are attributed by many to be due to some diffusible phytotoxic metabolites or toxins (or more precisely “the pathogen-produced determinants of disease”) produced by the fungus (Kerr, 1956; Wyllie, 1962; Sherwood and Lindberg, 1962; Aoki et al., 1963). The present study was undertaken to evaluate quantitative changes in the phenolics of groundnut hypocotyls brought about by S. rolfsii infection and to determine the nature of phenolic compounds produced by the pathogen in vitro.
MATERIALS AND METHODS
Isolation of pathogen: During the survey of groundnut fields around Srikalahasti and Tirupati areas of Chittoor District and also other Rayalaseema Districts of A.P., Sclerotium rolfsii was found to be associated with the infected hypocotyls region of groundnut variety TMV-2 at early stages of growth and development. The plants showing stem rot or southern blight symptoms were brought to the laboratory for making isolation according to the tissue segment method on PDA, pure culture was obtained by transferring the sclerotia to PDA plates. The stock culture was maintained on PDA slants in a refrigerator and subcultured for every two months.
DISEASE DEVELOPMENT
Method of raising plants: High quality seed material (average 95% germination) of groundnut variety TMV-2 was obtained from the local Agricultural Research. Sound seeds were surface sterilized with 0.1% mercuric chloride for 2-3 min followed by repeated washings with sterile water and sown in sterilized soil contained in seed pans. The seeds germinated and emerged in 4-5 days. One week old seedlings were used for inoculation purpose.
Inoculation: Ten days old oatmeal-sand culture of S. rolfsii was thoroughly mixed with sterilized soil at 10%. This inoculum-soil mixture was then distributed in 12" diameter earthenware pots and left undistributed for two days. After this period, one week old seedlings grown in seed pans were lifted carefully without causing much damage to the root system and transplanted into the pots. They were watered on alternate days and kept in an open atmosphere.
Disease indexing: The plants were periodically examined for the progress of the disease. Samples were collected at random from four pots each time at ‘0’ h, 2 days, 5 days, 9 days and 11 days after inoculation. Almost all the seedlings collapsed by 11 days after transplantation. The progress of disease in the hypocotyls of the seedlings could be differentiated into the following five fairly distinct stages, on the basis of lesion development:
| Stage 1 | : | 0 hours, i.e. immediately after inoculation: Healthy seedlings |
| Stage 2 | : | 2 days after inoculation: The early or young phase, characterized by water-soaked appearance of invaded portions of hypocotyls which remained almost colourless or were light brown in colour |
| Stage 3 | : | 5 days after inoculation: The intermediate stage, in which the lesion surface become brown to dark brown colour |
| Stage 4 | : | 9 days after inoculation: Well developed, dark necrotic lesions often girdling hypocotyls. This marks the final stage in lesion maturation. The lesions also showed sunken appearance |
| Stage 5 | : | 11 days after inoculation: Characterized by dry appearance of the lesion surface. Downward destruction or rotting of the tap root occurred and then the seedlings wilted and died |
Symptoms characteristic of each of the above stages of lesion maturation are shown in Fig. 1.
Collection of host tissue: For extraction of phenolics, hypocotyl regions with lesions and the corresponding hypocotyl portion of healthy seedlings were collected at the five different stages of lesion development cited above. They were washed thoroughly with distilled water to remove adhering soil particles and used immediately for extraction.
Changes in phenolic acids: Phenolic acids were extracted according to the method of Bate-Smith (1954) adopted by Das and Rao (1964). About 10-15 g of fresh material was extracted in 2 N HCl and digested on a boiling water bath for 20 min. The digest was filtered and the filtrate shaken with diethyl ether to extract phenolic acids into it. It was heated for 20 min and then shaken several times with ether (Reddy et al., 1975). All the other extracts were combined and the phenolic acids were taken up from the combined extract into 5% sodium carbonate which was acidified and reextracted with ether. The final ether extract was subjected to paper chromatographic analysis using a 2-dimensional ascending technique on Whatman No.1 chromatography paper. The solvents employed were benzene-acetic acid-water (60:70:30 upper phase) in the first direction and the sodium formate-formic acid-water (10:1:200) in the second direction (Ibrahim and Towers, 1960). Papers were run twice in the same direction in the first solvent to ensure better resolution.
The dried chromatograms were observed under ultraviolet light, first without and then with ammonia vapors, all the fluorescent spots were marked. The sheets were then sprayed with diazotized p-nitraniline (Smith, 1960) or diazotized sulphanilic acid (Ames and Mitchell, 1952) or 1% ferric chloride in order to identify the phenolic acids present in the extracts. Authentic samples were also developed under identical conditions.
 | |
| Fig. 1: | Groundnut seedlings inoculated with S. rolfsii, 1: Immediately after inoculation, 2, 3, 4 and 5: Disease development on stems until the maturation of lesions |
Identity of some of the compounds was also confirmed by co-chromatography with authentic samples. The areas form unsprayed sheets, corresponding in Rf values to the detected phenolic acids, were eluted with 90% ethanol. The eluates were evaporated to dryness and dissolved in 3 mL of distilled water; 0.5 mL of Folin’s reagent was added and thoroughly shaken. After 3 min, 1 mL of saturated sodium carbonate solution was added and the mixture was made up to 10 mL. After 1 h, the blue colour developed was read at 725 nm using Spectronic-20 colorimeter. The phenolic acid content of the individual spots was calculated by making use of the standard curves prepared from their respective authentic samples. The quantities of unidentified compounds were calculated from the standard curves prepared with known quantities of chlorogenic acid.
RESULTS
Phenolic acids occurring in healthy and S. rolfsii infected groundnut hypocotyls are presented in the Table 1. There is a tremendous increase in the number of phenolic acids in the diseased area of the groundnut hypocotyls. Chromatographic data revealed that there are altogether 10 phenolic acids in healthy plants, nine of which were identified as trans-caffeic acid, chlorogenic acid, cis-caffeic acid, cis-p- coumaric acid, p-hydroxybenzoic acid, trans-p-coumaric acid, trans-ferulic acid and vanillic acid and remaining one was unidentified. Infection resulted in a tremendous effect on the qualitative nature of phenolic compounds resulting also in the production of new or additional compounds. In the infected plants, in addition to the above mentioned ten phenolic compounds 6 additional compounds were detected with the progress of disease.
| Table 1: | Effect of S. rolfsii infection on phenolic acid content (mg g-1 fresh weight) of healthy and infected hypocotyls at various stages of disease development |
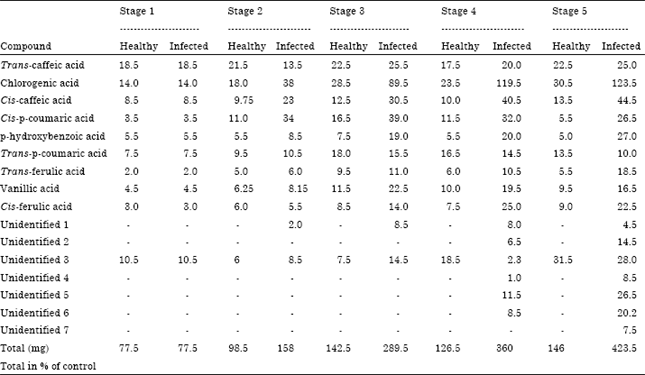 | |
| Each value is an average of 3 replicate samples | |
DISCUSSION
Many fungi produce metabolites in culture which are phototoxic and it is possible that such substances are released by fungi in the course of growth in their host tissues and they are involved in the production of disease symptoms (Reddy et al., 1975; Basha et al., 2006). The point of interest in this investigation is that significantly greater accumulation of phenolic compounds is observed in diseased plants than in healthy ones and that a correlation exists between the compounds produced by the pathogen in vitro and some of those that appeared a fresh in diseased plants. It is interesting to note that salicylic acid which also reported to induce resistance in host plants against pathogen attack was not detected in sclerotial exudates of Sclerotinia sclerotiorum (Basha et al., 2006). The occurrence of the new compounds in the S. rolfsii infected tissue may be due to the production by the fungus because the corresponding similar compounds were also observed in the culture filtrate. The production of several new compounds in the infected tissues indicates that they are apparently formed through the interaction of the fungus and the living host since no such compounds were observed in extracts of the healthy host and also neither in the fungus nor in culture filtrate of the fungus. It is also well established that the processes and mechanisms associated with disease development are a junction of both the host and the pathogen and that the disease determined abnormal physiological activity, strictly speaking, may not be characteristic of either the host or the pathogen individually but of the host-pathogen complex (Bateman, 1970). The fluorescent compounds may be involved in localization of the fungus and may limit lesion size (Deverall and Wood, 1961).
The chromatographic data, precipitation with lead acetate, fluorescence in UV radiation and reaction to different spray reagents used suggest that the fluorescent compounds are phenolic and acidic substances. Negative reactions of several unidentified compounds with diazotized sulphanilic acid, diazotized p-nitraniline and ferric chloride do not support this conclusion. It is possible however, that these compounds were not sufficiently concentrated to give visible reaction with these reagents.
Phenolics are well-known as antifungal, antibacterial and antiviral compounds occur naturally in plants (Basha et al., 2006; Leiss et al., 2009; Osman et al., 2011). According to Ali et al. (2009) the first step in defense mechanism in plants involves a rapid accumulation of phenols at the infection site, which restricts or slows the growth of the pathogen. The role of phenolic compounds in the host/pathogen interaction is well established (Sarma et al., 2002) and constitutive phenolics are known to confer resistance indirectly through activation of post-infection responses in the host (Harborne, 1988). Several studies have shown that some phenolics are inhibitors associated with non-host resistance (Nicholson and Hammerschmidt, 1992) whereas, others are formed or increased in response to pathogen infection and are considered to be an important component in the defence response of the pathogen (Harborne and Turner, 1984; Punja et al., 1985; Nicholson and Hammerschmidt, 1992).
CONCLUSION
The role of phenolic compounds in the host-pathogen interaction is well established and constitutive phenolics are known to confer resistance indirectly through activation of post-infection responses in the host. Several studies have shown that some phenolics are inhibitors associated with non-host resistance whereas, others are formed or increased in response to the pathogen infection and are considered to be an important component in the defense response on the host to the pathogen.
ACKNOWLEDGMENT
The authors are very much thankful to the Sri Padmavati Mahila Visvavidyalam, Tirupati for providing the facilities.
REFERENCES
- Ali, A.A., K.M. Ghoneem, M.A. El-Metwally and K.M. Abd El-Hai, 2009. Induce systemic resistance in lupine against root rot diseases. Pak. J. Biol. Sci., 12: 213-221.
CrossRefPubMedDirect Link - Adandonon, A., T.A.S. Aveling, N. A. van der Merwe and G. Sanders, 2005. Genetic variation among Sclerotium isolates from Benin and South Africa, determined using mycelial compatibility and ITS rDNA sequence data. Aust. Plant Pathol., 34: 19-25.
Direct Link - Basha, S.A., B.K. Sarma, K.P. Singh and U.P. Singh, 2006. Variation in biochemical composition among Indian isolates of Sclerotinia sclerotiorum. Microbiology, 34: 114-119.
Direct Link - Aoki, H., T. Sassa and T. Tamura, 1963. Phytotoxic metabolites of Rhizoctonia solani. Nature, 200: 575-575.
Direct Link - Bate-Smith, E.C., 1954. Leuco-anthocyanins. 1. Detection and identification of anthocyanidins formed leuco-anthocyanins in plant tissues. Biochem. J., 58: 122-125.
PubMedDirect Link - Cilliers, A.J., L. Herselman and Z.A. Pretorius, 2000. Genetic viability within and among mycelial compatibility groups of Sclerotium rolfsii in South Africa. Phytopathology, 90: 1026-1031.
Direct Link - Deverall, B.J. and R.K.S. Wood, 1961. Chocolate spot of beans (Vicia faba L.)-interactions between phenolase of host and pectic enzymes of the pathogen. Ann. Applied Biol., 9: 473-487.
CrossRefDirect Link - Ganesan, S., K.R. Ganesh and R. Sekar, 2007. Integrated management of stem rot disease (Sclerotium rolfsii) of groundnut (Arachis hypogaea L.) using Rhizobium and Trichoderma harzianum (ITCC-4572). Turk. J. Agric. For., 31: 103-108.
Direct Link - Leiss, K.A., F. Maltese, Y.H. Choi, R. Verpoorte and P.G. Klinkhamer, 2009. Identification of chlorogenic acid as a resistance factor for thrips in Chrysanthemum. Plant Physiol., 150: 1567-1575.
PubMed - Osman, M.E.H., M.M. El-Sheekh, M.A. Metwally, A.E.A. Ismail and M.M. Ismail, 2011. Antagonistic activity of some fungi and cyanobacteria species against Rhizoctonia solani. Int. J. Plant Pathol., 2: 101-114.
CrossRefDirect Link - Nalim, F.A., J.L. Starr, K.E. Woodard, S. Segner and N.P. Keller, 1995. Mycelial compatibility groups in Texas peanut field populations of Sclerotium rolfsii. Phytopathology, 85: 1507-1512.
CrossRef - Nicholson, R.L. and R. Hammerschmidt, 1992. Phenolic compounds and their role in disease resistance. Annu. Rev. Phytopathol., 30: 369-389.
CrossRefDirect Link - Pandey, M.K., B.K. Sarma and U.P. Singh, 2005. Induction of sexual stage and colony morphology of some isolates of Sclerotium rolfsii causing spotted leaf rot in plants. Microbiology, 33: 7-11.
Direct Link - Punja, Z.K., 1985. The biology ecology and control of Sclerotium rolfsii. Ann. Rev. Phytopathol., 23: 97-127.
CrossRef - Punja, Z.R., V.L. Smith, C.L. Campbell and S.F. Jenkins, 1985. Sampling and extraction procedure to estimate numbers, spatial pattern and temporal distribution of sclerotia of Sclerotium rolfsii in soil. Plant Dis., 69: 469-474.
Direct Link - Sarma, B.K., U.P. Singh and K.P. Singh, 2002. Variability in Indian isolates of Sclerotium rolfsii. Mycologia, 94: 1051-1058.
Direct Link - Sennoi, R., S. Jogloy, W. Saksirirat and A. Patanothai, 2010. Pathogenicity test of Sclerotium rolfsii, a causal agent of jerusalem artichoke (Helianthus tuberosus L.) stem rot. Asian J. Plant Sci., 9: 281-284.
CrossRefDirect Link