
S.K. Fathima
Post Graduate Department of Microbiology, Maharani�s Science College for Women, JLB Road, Mysore-570005, Karnataka, India
S. Shankara Bhat
Department of Studies in Microbiology, Manasagangotri, University of Mysore, Mysore-570006, Karnataka, India
K. Girish
Post Graduate Department of Microbiology, Maharani�s Science College for Women, JLB Road, Mysore-570005, Karnataka, India
International Journal of Plant Pathology
Year: 2012 | Volume: 3 | Issue: 2 | Page No.: 56-65







ABSTRACT
A study on cultural, morphological and biochemical variability among the isolates of Phomopsis azadirachtae Sateesh, Bhat and Devaki collected from sixteen districts of Karnataka state, South India was carried out. The isolates were compared for the cultural and biochemical variability. It was observed that the isolates varied considerably for the cultural characteristics such as mycelial growth pattern, colour of the colony, sporulation behaviour, texture of pycnidia, production of alpha and beta conidia and the dimension of conidia produced. Estimation of water soluble mycelial proteins and toxicity of culture filtrate of P. azadirachtae isolates revealed a remarkable difference in the mycelial protein content of the isolates and different isolates exhibited various extent of toxicity on the germination of neem seeds.
PDF Abstract XML References Citation
Received: November 01, 2011;
Accepted: March 05, 2012;
Published: June 05, 2012
How to cite this article
S.K. Fathima, S. Shankara Bhat and K. Girish, 2012. Cultural, Morphological and Biochemical Variability among the Isolates of Phomopsis azadirachtae from Karnataka. International Journal of Plant Pathology, 3: 56-65.
DOI: 10.3923/ijpp.2012.56.65
URL: https://scialert.net/abstract/?doi=ijpp.2012.56.65
DOI: 10.3923/ijpp.2012.56.65
URL: https://scialert.net/abstract/?doi=ijpp.2012.56.65
INTRODUCTION
Neem (Azadirachta indica A. Juss.) a member of mahogany family Meliaceae is well-known for its biomedical properties. It is an evergreen, ecofriendly, native tree of Indian sub-continent. Neem is a source of effective bio-pesticides and provides a cure for many ailments right from common cold to cancer and AIDS (Anonymous, 2009; Girish and Bhat, 2008a). The eco-friendly neem tree, in spite of having antimicrobial properties, is infected by various pathogens belonging to bacteria and fungi. The most destructive pathogen of neem at present is Phomopsis azadirachtae Sateesh, Bhat and Devaki which causes die-back disease (Sateesh et al., 1997). The chief symptoms of the disease are twig blight, inflorescence blight and fruit rot. The disease results in almost 100% loss of fruit production (Bhat et al., 1998; Girish and Bhat, 2008b), because of which, neem seeds used as a raw material in the preparation of bio-pesticides, medicines and various industrial products are not obtained.
Plant pathogens are important components of the biodiversity of all natural ecosystems. Remarkable differences in cultural, morphological and biochemical characteristics are observed among pathogen populations from different geographical locations (Thakur, 1999). Differences in cultural, morphological and biochemical characteristics have been reported among populations of various plant pathogens (Sharma et al., 2002; Basandrai et al., 2005; Khurana et al., 2005; Hosen et al., 2010). Phytopathogenic fungi exhibit intraspecific variability in toxin production (Nandakumar et al., 2007; Asran and Amal, 2011; Singh and Kumar, 2011).
A good number of Phomopsis species are reported to exhibit remarkable intraspecific variability (Brayford, 1990; Shivas et al., 1991). Die-back disease of neem caused by P. azadirachtae is widespread in different parts of Karnataka State, South India (Sateesh, 1998). Girish and Bhat (2010) reported the presence of morphological, cultural and biochemical variability among the P. azadirachtae collected from different regions of Tamil Nadu state, India. Significant differences were found in the protein banding patterns of P. azadirachtae isolates from Karnataka (Fathima et al., 2004) and Tamil Nadu (Girish et al., 2009), India. The present investigations were undertaken to study the morphological, cultural and biochemical variability among the isolates of P. azadirachtae collected from 16 districts of Karnataka State.
MATERIALS AND METHODS
Sites sampled: The collections of infected neem twig samples were made from Mandya, Gulbarga, Mysore, Bijapur, Hassan, Chikmagalur, Shimoga, Kolar, Raichur, Tumkur, Chamarajnagar, Chitradurga, Belgaum, Davanagere, Bengaluru and Bellary. The twigs were either used immediately after their collection or stored in brown paper bags or polythene covers in a refrigerator at 4°C until used for further study.
Isolation of P. azadirachtae from infected neem shoot: The infected twigs collected from each district were cut into 2-3 cm pieces with middle transition region of healthy and infected portion. Healthy twig explants served as control. Both healthy and infected twig pieces were washed separately with running tap water for one hour. Later they were cut into 1-1.5 cm segments with transition zone at the middle portion. The bark was removed and the segments were washed thoroughly and were surface-sterilized using sodium hypochlorite solution (with 5% available chlorine) (Sauer and Burroughs, 1986). Then they were rinsed six times with sterile distilled water. The explants were plated on Potato Dextrose Agar (PDA) medium amended with 200 ppm of chloramphenicol in Petri plates at the rate of four segments per plate (Sateesh, 1998). The inoculated Petri plates were incubated for 3-7 days in dark and observed for the growth of the pathogen from the twig segments (Alexopoulos et al., 1996). Later the plates were kept near the window on the laboratory bench for natural photoperiod and the pathogen was allowed to sporulate.
Mycelial plugs (5 mm diam.) were removed from the advancing margins of 7 day old cultures of each isolate and transferred onto fresh PDA in Petri plates (90 mm diam.). Three replicates of each isolate were maintained and the plates were incubated for 10 days at 26±2°C for 12 h photoperiod. The isolates were identified as per Sateesh et al. (1997). The isolates were designated as follows: Mandya-Pa 01, Gulbarga-Pa 02, Mysore-Pa 03, Bijapur-Pa 04, Hassan-Pa 05, Chikmagalur-Pa 06, Shimoga-Pa 07, Kolar-Pa 08, Raichur-Pa 09, Tumkur-Pa 10, Chamarajnagar-Pa 11, Chitradurga-Pa 12, Belgaum-Pa 13, Davanagere-Pa 14, Bengaluru-Pa 15 and Bellary-Pa 16.
Cultural characteristics: Mycelial plugs were removed from the advancing margins of 7 day old cultures of each isolate and transferred onto fresh PDA plates. Three replicates of each isolate were maintained and incubated for 10 days at room temperature (26±2°C) with 12 h photoperiod. The single spore culture (pure culture) was obtained on PDA medium by inoculating a single spore (Tuite, 1969). The cultures thus obtained were compared for their cultural characteristics. The colony morphology was assessed for growth characteristics, mycelial type, colour of the colony, colony diameter and texture and growth pattern of pycnidia. To measure the conidial dimension, the spores were collected onto the microscope slides, by touching the spore ooze. They were spread by adding a drop of cotton blue in lactophenol. About 100 alpha (α) and 100 beta (β)-conidia were measured in case of each isolate (Uecker and Caruso, 1988).
Estimation of water soluble mycelial proteins of P. azadirachtae isolates: All the 16 isolates of P. azadirachtae were grown separately on Czapek Dox broth for seven days. Mycelial mat was taken out, washed thoroughly with distilled water and dried using filter paper. One gram of the dried mycelium of each isolate was homogenized in 10 mL of TCA and centrifuged at 3000 xg for 20 min. The residue was re-dissolved in 10 mL of phosphate buffer and centrifuged. The supernatant was used for protein estimation as per Lowry et al. (1951).
Effects of culture filtrate of P. azadirachtae isolates on germination of neem seeds: To determine the variability in toxicity of culture filtrate among the isolates of P. azadirachtae, the filtrate of each isolate was obtained from 25 day old culture grown on potato dextrose broth. About 100 healthy surface-sterilized neem seeds were treated with the culture filtrate of each isolate separately by incubating in a beaker for 24 h. The control treatment included medium alone and sterile distilled water. The seeds were plated on sterile moist blotter in petri plates or on standard paper towels (ISTA, 1993) and incubated at room temperature (26±2°C). Each treatment had four replications. After 10 days shoot length, root length and percentage germination were recorded. Vigour index was calculated using the formula given by Abdul-Baki and Anderson (1973).
RESULTS
Sites sampled and isolation of P. azadirachtae from infected neem shoots: The field survey of various districts of Karnataka state, South India revealed the occurrence of die-back disease in all the 16 districts of Karnataka state, South India. The infected neem segments collected from all the locations developed fungal colonies whose identification was confirmed as P. azadirachtae as per Sateesh et al. (1997). All the isolates produced characteristic α and β-conidia.
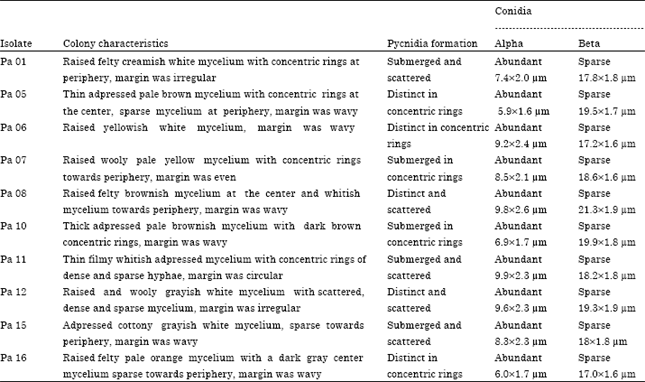
Cultural characteristics: The isolates of P. azadirachtae derived from naturally infected neem shoots exhibited variable colony characteristics on PDA plates at 26±2°C. The isolates varied considerably for mycelial growth, colour of the colony, sporulation behaviour, growth pattern, texture and formation of pycnidia, production of alpha and beta conidia and their length and breadth (Fig. 1). The cultural characteristics of ten of the isolates are presented in the Table 1. Isolate Pa 14 had the longest alpha conidia (11.2x3.0 μm) while Pa 09 had the shortest alpha conidia (4.2x1.0 μm). Isolate Pa 04 had the longest beta conidia (24.2x1.8 μm) while shortest beta conidia (15.3x1.4 μm) were observed in isolate Pa 13. The isolates also varied significantly for colony growth and sporulation rate. Isolate Pa 07 had the largest colony (74 mm diam.) while Pa 06 had the smallest (32 mm diam) (Fig. 2).
 | |
| Fig. 1: | Ten day old colonies of P. azadirachtae collected from different regions of Karnataka on potato dextrose agar |
Maximum sporulation was observed in Pa 08 (280 to 300 pycnidia/colony), while it was minimum in Pa 11 (25 to 40 pycnidia/colony) (Fig. 3). Further almost all the isolates produced more number of α conidia than β-conidia except the isolate Pa 04 which produced large number of beta conidia and very few alpha conidia with an alpha to beta conidia ratio of 8:42 unlike rest of the 15 isolates in which the ratio ranged between 35:15 to 44:6.
Estimation of water soluble mycelial proteins of P. azadirachtae isolates: The isolates of P. azadirachtae showed significant difference in their mycelial protein content.
| Table 1: | Colony characteristics of the 10 isolates of P. azadirachtae on potato dextrose agar plates after 10 days of incubation at 26±2°C |
 | |
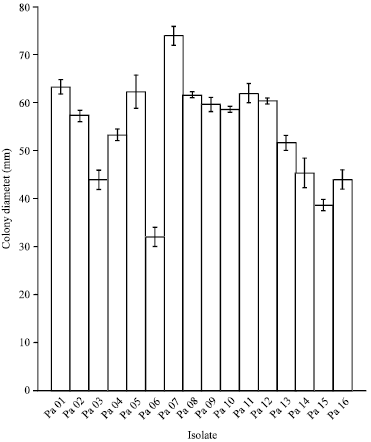 | |
| Fig. 2: | Colony diameter of 16 isolates of P. azadirachtae on potato dextrose agar medium after 10 days of incubation at 26°C, Bars indicate standard deviation |
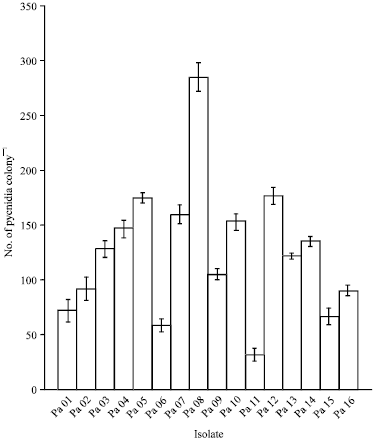 | |
| Fig. 3: | Sporulation rate of 16 isolates of P. azadirachtae on potato dextrose agar medium after 10 days of incubation at 26°C, Bars indicate standard deviation |
| Table 2: | Water soluble mycelial protein of 16 isolates of P. azadirachtae |
 | |
| Values given are mean of three replicates ±SE, Values followed by different superscript letters differ significantly when subjected to Tukey’s HSD (honestly significant difference) | |
The protein content of different P. azadirachtae isolates is presented in Table 2. Isolate Pa 04 had the highest mycelial protein content 150 μg g-1 whereas the isolate Pa 11 had the lowest mycelial protein content 85 μg g-1.
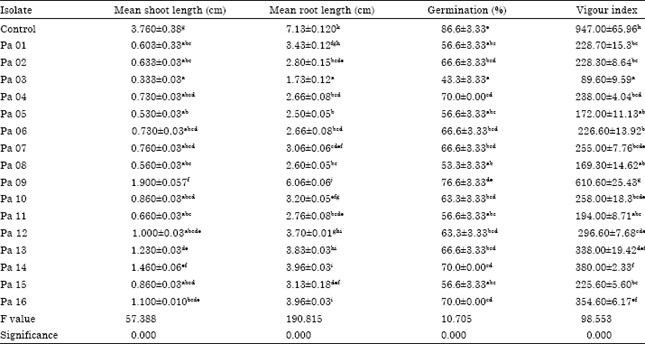
| Table 3: | Effect of culture filtrate of 16 isolates of P. azadirachtae on germination of neem seeds |
 | |
| Values given are means of three replicates ±SE, Values followed by different superscript letters differ significantly when subjected to Tukey’s HSD (honestly significant difference) | |
Effects of culture filtrate of P. azadirachtae isolates on germination of neem seeds: The neem seeds treated with culture filtrate of isolates of P. azadirachtae collected from various districts showed reduction in their germination and seed vigour when compared to control seeds in both Petri plates and paper towels. Further it was observed that the culture filtrate of different isolates exhibited different extent of toxicity on the germination of neem seeds. The difference in the level of toxicity is presented in the Table 3. The seeds treated with isolate Pa 03 culture filtrate showed lowest percentage of germination and the seedling quality was also significantly affected. Isolate Pa 09 culture filtrate was least toxic compared to all other isolates.
DISCUSSION
Fungi are a unique group of microorganisms with their wide range of adaptability in different ecological situations (Moore, 1996). The genus Phomopsis is important plant pathogen (Raeisi et al., 2011) and reported to be a highly variable pathogen (Brayford, 1990; Shivas et al., 1991; Akhtar and Chaube, 2002). The present study revealed remarkable differences in cultural, morphological and biochemical characteristics of various P. azadirachtae isolates collected from different agroclimatic regions of Karnataka State, South India. Similar differences were observed among the P. azadirachtae isolates of Tamil Nadu, in radial growth, colony morphology, colour, texture, pycnidial density and arrangement, conidial number and size (Girish and Bhat, 2010). Similar variation was reported in Phomopsis oblonga isolated from Ulmus species in the British Isles and Italy (Brayford, 1990). Intraspecific variation was demonstrated among the isolates of Phomopsis leptostromiformis the causal agent of stem canker on Lupinus angustifolius using cultural and biochemical techniques (Shivas et al., 1991). Similar variability among the isolates of other phytopathogenic fungi was also reported. Rhynchosporium secalis isolates from barley in different agroecological zones of Ethiopia differed markedly in several cultural characteristics like colony and conidial morphology, colony growth rate and sporulation (Meles et al., 2004). Isolates of Sclerotinia sclerotiorum collected from infected lentil plants from two agroecological zones of Syria showed considerable variation in cultural characteristics such as mycelia growth, mycelia pigmentation and sclerotial production (Akem et al., 2006). Two isolates of Pythium ultimum var. ultimum isolated from Egypt and Germany varied in the size of sexual organs, oospore production, growth rate and other morphological and physiological characteristics (Al-Sheikh and Abdelzaher, 2010).
The present investigations revealed that in one of the isolates of P. azadirachtae, designated as Pa 04 collected from Bijapur district, there was abundant production of beta conidia. Generally beta conidia production is influenced markedly by the substrate (Parmeter, 1958). The remarkable difference in alpha to beta conidia ratio in the isolate Pa 04 in the present study could be possibly due to the difference in the host growing in different geographical locations.
Studies on non-morphological characteristics such as wall composition, proteins and other hydrocarbons, of fungal isolates becomes important to differentiate among isolates when the strains no longer produce typical morphological structure (Jernejc and Cimerman, 2001). A remarkable variation in the mycelial protein content of various isolates of P. azadirachtae was observed in the present study. Similar observations were made by Girish and Bhat (2010) among the Tamil Nadu isolates of P. azadirachtae and Khurana et al. (2005) among the Alternaria brassicae isolates. The variations could be possibly due to the difference in the geographical locations from where the isolates are collected.
A good majority of deuteromycetes are reported to release toxic secondary metabolites into the media (Maude, 1996; Agrios, 2004). Sateesh, (1998) indicated the release of a toxic metabolite by P. azadirachtae into the medium which reduces the seed vigour and seed quality, based on the study of effects of culture filtrate on germination of neem seeds. In the present investigations, a remarkable difference in toxicity of culture filtrate was observed among various isolates of P. azadirachtae on the germination of neem seeds. Similar observation was reported by Shivas et al. (1991) among Phomopsis leptostromiformis isolates. Variation in toxin production among the isolates of Sarocladium oryzae causing rice sheath blight was reported by Nandakumar et al. (2007). Isolates of Fusarium moniliforme from pepper plants produced different quantities of each of fumonisin, zearelenone in vitro (Abo-Elnaga and Ahmed, 2007). A remarkable difference in toxin production was observed among the isolates of Fusarium oxysporum f. sp. chrysanthemi pathogenic to Chrysanthemum (Singh and Kumar, 2011).
The results of present study suggest that P. azadirachtae is a highly variable pathogen. These variations can be attributed to the influence of host variety and prevailing environment. This variability in pathogen populations makes them capable of invading a wide variety of host plants, thereby reducing the possibility of evolving disease management methods (Moore, 1996). The primary objective of studies on variation in pathogenicity or virulence is breeding and exploitation of resistance for disease management. The study of variations among the fungal isolates will also be helpful in characterizing sub-specific taxa and thereby in fungal taxonomy (Jernejc and Cimerman, 2001).
ACKNOWLEDGMENTS
The authors thank University of Mysore for facilities. Syeda Kousar Fathima acknowledges UGC, New Delhi for granting FIP fellowship.
REFERENCES
- Abdul-Baki, A.A. and J.D. Anderson, 1973. Vigor determination in soybean seed by multiple criteria. Crop Sci., 13: 630-633.
CrossRefDirect Link - Akem, C., M. Bellar and B. Bayaa, 2006. Comparative growth and pathogenecity of geographical isolates of Sclerotinia sclerotiorum on lentil genotypes. Plant Pathol. J., 5: 67-71.
Direct Link - Al-Sheikh, H. and H.M.A. Abdelzaher, 2010. Differentiation between two isolates of Pythium ultimum var. ultimum isolated from diseased plants in two different continents. J. Biol. Sci., 10: 306-315.
CrossRefDirect Link - Asran, M.R. and M.I.E. Amal, 2011. Aggressiveness of certain Fusarium graminearum isolates on wheat seedlings and relation with their trichothecene production. Plant Pathol. J., 10: 36-41.
CrossRefDirect Link - Basandrai, A.K., S. Pande, G.K. Kishore, J.H. Crouch and D. Basandrai, 2005. Cultural, morphological and pathological variation in Indian isolates of Ascochyta rabiei, the chickpea blight pathogen. Plant Pathol. J., 21: 207-213.
Direct Link - Hosen, M.I., A.U. Ahmed and M.R. Islam, 2010. Variability and pathogenecity in Bangladeshi isolates of Botrytis cinerea causing botrytis gray mold in chickpea (Cicer arietinum). Plant Pathol. J., 3: 129-134.
Direct Link - Jernejc, K. and A. Cimerman, 2001. Morphological characteristics, extracellular and intracellular protein and enzyme patterns of five Aspergillus species. Food Technol. Biotechnol., 39: 333-340.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Meles, K., M. Hulluka and M.M. Abang, 2004. Phenotypic diversity in Rhynchosporium secalis from Ethiopia and host response to barley scald. Plant Pathol. J., 3: 26-34.
CrossRefDirect Link - Nandakumar, R., S. Babu, G. Amutha, T. Raguchander and R. Samiyappan, 2007. Variation in toxin production among isolates of Sarocladium oryzae the rice sheath rot pathogen. Plant Pathol. J., 6: 120-126.
CrossRefDirect Link - Raeisi, S., A. Puteh and K. Sijam, 2011. First report of Diaporthe/Phomopsis complex isolates in soybean from Malaysia and their longevity in stored seeds. Asian J. Plant Pathol., 5: 146-154.
CrossRefDirect Link - Sauer, D.B. and R. Burroughs, 1986. Disinfection of seed surfaces with sodium hypochlorite. Phytopathology, 76: 745-749.
CrossRefDirect Link - Sharma, S.K., D.V. Singh, R. Aggarwal and K.D. Srivastava, 2002. Pathogenic and biochemical variations in Neovossia indica. Indian Phytopathol., 56: 133-139.
Direct Link - Singh, P.K. and V. Kumar, 2011. Variability among isolates of Fusarium oxysporum f.sp.chrysanthemi pathogenicto chrysanthemum. Int. J. Plant Pathol., 2: 136-143.
CrossRef - Thakur, R.P., 1999. Pathogen diversity and plant disease management. Indian Phytopathol., 52: 1-9.
Direct Link