
Javan Omondi Were
Department of Seed, Crop and Horticultural Sciences, School of Agriculture and Biotechnology, University of Eldoret, P.O. Box 1125, 30100 Eldoret, Kenya
LiveDNA: 254.31155
Julius Onyango Ochuodho
Department of Seed, Crop and Horticultural Sciences, School of Agriculture and Biotechnology, University of Eldoret, P.O. Box 1125, 30100 Eldoret, Kenya
Nicholas Kipkemboi Rop
Department of Seed, Crop and Horticultural Sciences, School of Agriculture and Biotechnology, University of Eldoret, P.O. Box 1125, 30100 Eldoret, Kenya
Sanjaya Gyawali
International Center for Agricultural Research in the Dry Areas, Biodiversity and Integrated Gene Management Program, Morocco
International Journal of Plant Breeding and Genetics
Year: 2020 | Volume: 14 | Issue: 1 | Page No.: 1-8







ABSTRACT
Background and Objective: Aluminium cation toxicity remains to be one of the major abiotic factors limiting barley production not only in Kenya but also other parts of the world where this crop is grown. The study aimed at screening both commonly grown barley varieties as well as the improved barley lines to identify potential varieties that could be used in improving tolerance to aluminium toxicity. Materials and Methods: Thirty two winter and spring adapted genotypes were arranged in a completely randomized design consisting of two Al treatments (0 and 148 μM) and barley genotypes as experimental units replicated 5 times. Five-day old seedlings were subjected to hematoxylin staining with 0.2% hematoxylin solution containing 0.02% potassium iodide. Visual assessment on the intensity of root staining was done on a 1-5 scale with 1 as tolerant and 5 as sensitive to aluminium toxicity. Data on net root length, relative net root growth, percent response, hematoxylin staining intensity were subjected to analysis of variance (ANOVA) on Genstat statistical software release 16.0 VSN International Ltd., at 5% level of significance. Mean response were separated using duncan multiple range test. Results: Winter and spring adapted barley differed significantly in their response to aluminium cation toxicity (p<0.05). Net root length, relative net root growth, percentage response, degree of root staining for spring adapted genotypes expressed more tolerance to aluminium toxicity compared to the winter adapted barley. Irrespective of the adaptation to growth conditions, some genotypes expressed exclusion while others utilized fixation mechanisms of tolerance to aluminium toxicity. Conclusion: In conclusion, a number of barley grown in Kenya were tolerant to aluminium toxicity and the study recommends Fanaka, MALT 1, Aliciana and Grace genotypes for studies aiming at increasing tolerance to aluminium toxicity.
PDF Abstract XML References Citation
How to cite this article
Javan Omondi Were, Julius Onyango Ochuodho, Nicholas Kipkemboi Rop and Sanjaya Gyawali, 2020. Rapid Screening Method for Tolerance to Aluminium Toxicity in Barley: Case Study on Commonly Grown and Newly Bred Genotypes in Kenya. International Journal of Plant Breeding and Genetics, 14: 1-8.
DOI: 10.3923/ijpbg.2020.1.8
URL: https://scialert.net/abstract/?doi=ijpbg.2020.1.8
DOI: 10.3923/ijpbg.2020.1.8
URL: https://scialert.net/abstract/?doi=ijpbg.2020.1.8
INTRODUCTION
Barley (Hordeum vulgare L.), a member of the grass family, is a major cereal grain and is ranked fourth among cereal crops in the world in terms of quantity produced (136 million t) and in area under cultivation (566,000Km²)1. Important uses include industrial processing as alcoholic and non-alcoholic beverages such as beer, wines, spirits, food for humans with 8 essential amino acids, carbohydrates and several minerals2 and medicinal uses as a component of various health foods to control urinary tract infections, remove toxic substances from kidney and reduce chances of type 2 diabetes among others3,4. Despite its role in the Kenyan economy, the annual barley yields remain very unpredictable and below5 3.0 t ha–1. Additionally, in the past 2 decades, the annual area under barley in Kenya has been on the decreasing trend (below 20,000 ha) since late 1990s and this has persisted to date6. As a result, deficits have been experienced in Kenya since most farmers hardly attain potential yield recorded at 5.5 t ha–1 under experimental conditions6.
Aluminium cation toxicity has been identified to be among the major abiotic factors responsible for low barley yields in Kenya and other parts of the world7-9. Specifically, in Kenya, approximately 7.5 million ha of agricultural land is acidic10,11 with pH below 6.0 and this problem is further magnified by inappropriate phosphate fertilizers preferred by most farmers in the region12. This implies that there is higher concentration of aluminium which affects growth and development of most crops including barley13. Several attempts have been made through breeding and releasing a number of barley varieties for commercial production in the past three decades. However, the time taken by the newly released varieties to reach commercialization stage is too long and does not synchronize well with the fast changing soil chemical status. This calls for not only a rapid screening method but also a reliable technique that shortens the period taken by breeders to release a new variety for commercial production.
Additionally, conventional, mutation and molecular breeding approaches have been used widely to produce barley genotypes with tolerance traits to aluminium toxicity. However, the problem still persist as the negative impact on yield soars both nationally and internationally14. Other than the effects on yields, exposure to aluminium toxicity interferes with normal physiological and biochemical functions in barley and exclusion or fixation mechanisms to enhance tolerance to aluminium toxicity9. Among the sensitive genotypes, aluminium cations causes root and shoot stunting, reduced nutrient uptake by the roots, increased cell wall and cell membrane rigidity and interference with the double helix structure of the deoxyribonucleic acid (DNA)9 among others. In the long run, the yields are significantly reduced.
Therefore as a step towards ensuring increased barley production under acidic soils in Kenya, there is need for a rapid screening technique that meets the breeders’ timeline. This aimed at identifying the commonly grown winter and spring adapted barley genotypes that can be utilized in a breeding program with the aim of conferring tolerance to aluminium toxicity under laboratory conditions using nutrient solution. This approach would shorten the time taken by breeders to produce new varieties with enhanced tolerance to aluminium toxicity and yields under acidic soil conditions.
MATERIALS AND METHODS
Study area: The study was conducted at the University of Eldoret, School of Agriculture and Biotechnology in the department of seed, crop and Horticultural Science, Seed Science Laboratory over a period of 6 months from January-June, 2018.
Germination of barley seed: The 16 winter and 16 spring adapted barley genotypes were assessed for their response to aluminium toxicity as described by Magnavaca with slight modification using 148 μM Al treatment from AlCl3.6H2O15,16. The seeds for each genotype were soaked in sterile distilled water for 5 min then surface sterilized with 1% sodium hypochlorite for 5 min to reduce the chances of contamination during germination. Sterilized seeds were rinsed 8 times using sterile distilled water to remove traces of chloride and pre-germinated on paper rolls that had been previously moistened with aerated distilled water then placed inside germination trays. The trays were then incubated in a growth chamber at 27°C under dark conditions for 3 days17.
Experimental design for aluminium screening in the laboratory: The winter and spring barley genotypes were separately arranged in a completely randomized design consisting of 2 Al treatments (0 and 148 μM) and barley genotypes as experimental units replicated 5 times. The 8 L trays were used to hold nutrient solution under continuous aeration using a 50 L air compressor pump. The 3 day-old uniform-sized barley seedlings with no visible injury or root damage were placed inside plastic cups that are mounted on a perforated Styrofoam sheet, 35 seedlings/sheet (7 genotypes replicated 5 times). The transferred seedlings were then stabilized in a nutrient solution17 at a pH of 4.0 for 24 h. Thereafter, the seedlings were transferred to a freshly prepared nutrient solution containing aluminium as AlCl3 adjusted to a final concentration of 0 μM (control) and 148 μM aluminium (treatment). Each seedling was placed individually on cups (3 cm diameter) with small holes (0.5 cm) at the bottom of each cup to allow roots reach the solution inside a tray. The pH of the nutrient solution was maintained at 4.0 and monitored daily throughout the experiment.
After transfer into nutrient solution with and without aluminium, the seedlings were allowed to grow in a growth chamber for a photoperiod of 14 h of light and 10 h of darkness on daily basis. The photoperiod growth conditions were set at approximately 340 μmoles m–2 sec–1 of light intensity, temperature at 30°C and relative humidity at 70%. The dark growth conditions were set at approximately 22°C and 90% relative humidity inside the growth chamber.
Hematoxylin staining: Hematoxylin staining was used as a confirmatory test for tolerance to aluminium toxicity. All barley genotypes were subjected to hematoxylin root staining18 where 0.2% hematoxylin solution containing 0.02% potassium iodide was prepared in distilled water. The 5 day old seedlings grown under aluminium solution were gently shaken in 200 mL distilled water for 15 min on a mechanical shaker at 20 rpm to remove cations from the root surface. The water was then replaced by 200 mL of aqueous hematoxylin stain [0.2% hematoxylin and 0.02% potassium iodide, w/v] and left at the same low agitation for 20 min. The visual scores on degree of root staining was done on a 1-5 scale as follows: 1-Non-stained roots, classified as very tolerant, 2-Faintly stained roots, classified as tolerant, 3-Moderately stained roots, classified as moderately tolerant, 4-Well stained roots, classified as sensitive, 5-Deeply stained roots, classified as very sensitive genotypes.
Data collection on response to aluminium toxicity: The aluminium tolerant and sensitive genotypes were differentiated based on net root length (NRL), relative net root growth (RNRG), degree of hematoxylin staining and percent response17. The root length measurements were performed using a calibrated ruler. The initial root length (IRL) was measured at the time of seedling transfer to the nutrient solution. Five days after transfer into aluminium treatments with 0 and 148 μM, the final root length (FRL) was measured. The NRL was obtained from the difference between FRL and IRL (NRL = FRL-IRL). The relative net root growth (RNRG) was obtained by dividing the corresponding NRL in 148 μM aluminium treatment set by NRL in 0 μM aluminium treatment (RNRG = NRL (Al)/NRL (Control). The greater the RNRG, the more resistant a genotype is to aluminium toxicity. The percent response to aluminium toxicity was also calculated using the formula:
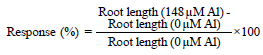
Statistical analysis: Data on NRL, RNRG, response (%), hematoxylin staining intensity were subjected to analysis of variance (ANOVA) on Genstat statistical software release 14.1 VSN International Ltd at 5% level of significance. The staining intensity data on a 1-5 scale was first transformed (Log10+1) before performing an ANOVA to ensure the data set assumes the normal distribution thus obeying the central limit theorem (CLT). Mean differences among the barley genotypes belonging to winter and spring growth adaptations was separated using Duncan multiple range.
RESULTS
Winter and spring adapted genotypes showed mixed reactions to aluminium cation toxicity. In particular, both winter and spring barley differed significantly in terms of net root length (NRL), response (%), relative net root growth (RNRG) and degree of hematoxylin staining (p<0.05). Mixed reaction to aluminium toxicity was evident across a number of genotypes irrespective of whether they are winter or spring adapted.
Among the spring adapted barley, with reference to NRL, Sabini, Nguzo and HKBL 1674-4 genotypes were the most tolerant to Al toxicity and scored 7.0, 5.6 and 5.3 cm, respectively. In contrast to tolerance, HKBL 1861-1, Ngao and HKBL 1805-3 were the most sensitive to Al toxicity at 1.8, 2.7 and 2.8 cm in the same order. The relative net root growth (RNRG) ratio ranked genotype HKBL 1719-4 to be the most tolerant to Al toxicity with a score of 1.0, indicating that the root length under 148 μM and 0 μM did not differ in length. In addition, majority of the spring barley were tolerant to Al and scored above 0.7 in terms of RNRG but HKBL 1861-1 consistently remained the most sensitive to Al toxicity just as observed earlier in terms of NRL (Table 1).
The response (%) measurements further confirmed Sabini to be most tolerant genotype to Al toxicity by recording the lowest percentage. In this respect, other genotypes including HKBL 1719-4, HKBL 1774-3, Nguzo and those with percent response below 20% were tolerant to Al cation toxicity.
| Table 1: | Response of spring barley to aluminium toxicity in nutrient solution containing 148 μm Al3+ under laboratory conditions |
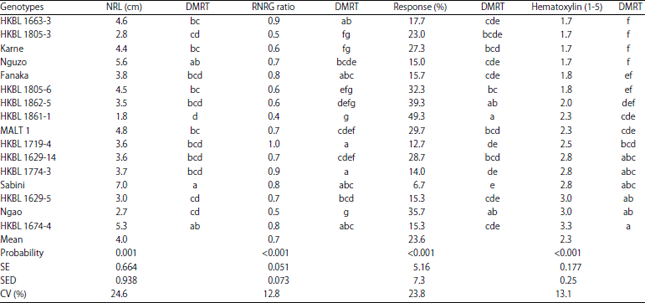 | |
NRL: Net root length, RNRG: Relative net root growth, DMRT: Duncan multiple range test | |
Similarly, HKBL 1861-1, Ngao and HKBL 1805-6 expressed higher sensitivity to Al toxicity and most of the observations in terms of percent response correspond to NRL and RNRG scores. In terms of the degree of hematoxylin staining, it is evident that different spring adapted barley genotypes utilized different mechanisms of responses to Al toxicity. For instance, HKBL 1663-3, HKBL 1805-3, Karne and Nguzo exhibited the highest tolerance to Al cation toxicity by recording the least degree of staining (Fig. 1a,b).
Group ‘a’ and ‘c’ were considered tolerant to aluminium toxicity. However, in group ‘a’, much aluminium cations were absorbed hence much staining but in group ‘c’, the aluminium was excluded from the roots thus low degree of staining. Group ‘b’ absorbed more aluminium cations which significantly affected root growth and development thus sensitive. The same scenario was observed in Sabini and HKBL 1719-4 genotypes which despite absorbing much Al cations into their root systems, they still expressed tolerance in terms of NRL, RNRG and percent response. The roots of such genotypes are less affected when exposed to higher concentration of Al cation, an indication of a totally different tolerance mechanism compared to those that exclude Al cations from their roots (Table 1 and Fig. 1A).
The winter adapted barley expressed similar patterns of response to Al cation toxicity in terms of NRL, RNRG, percent response and degree of hematoxylin staining. However, majority of the genotypes were more tolerant to Al toxicity compared to the spring adapted barley. For example, Quench, Titouan and SY Batyk were the most tolerant to Al toxicity not only in reference to NRL but also in terms of RNRG, percent response and hematoxylin staining except that SY Batyk used a different mechanism of tolerance compared to Quench and Titouan. Grace was more tolerant to cation toxicity than Aliciana but the two genotypes were the best in terms of excluding aluminium from their roots (Table 2).
Other than tolerance, NFC Tipple, Beatrix and Scrabble genotypes were more sensitive to Al toxicity with reference to their performance in terms of NRL, RNRG and percentage response. However, despite their nature of being sensitive to mineral toxicity, Scrabble and Beatrix absorbed much Al cations into their roots as indicated by the degree of hematoxylin staining. On the other hand, NFC Tipple excluded much cation from the root system but still remained very sensitive to low doses of Al cation especially in terms of percent response which corresponds to RNRG and NRL scores (Table 2).
The degree of hematoxylin staining and root morphologies formed three major groups among the winter adapted barley genotypes screened against Al cation toxicity. These include groups ‘d’, ‘e’ and ‘f’ just like the spring barley genotypes. Group ‘d’ were highly sensitive to aluminium toxicity while groups ‘e’ and ‘f’ were tolerant but with different response mechanisms of tolerance.
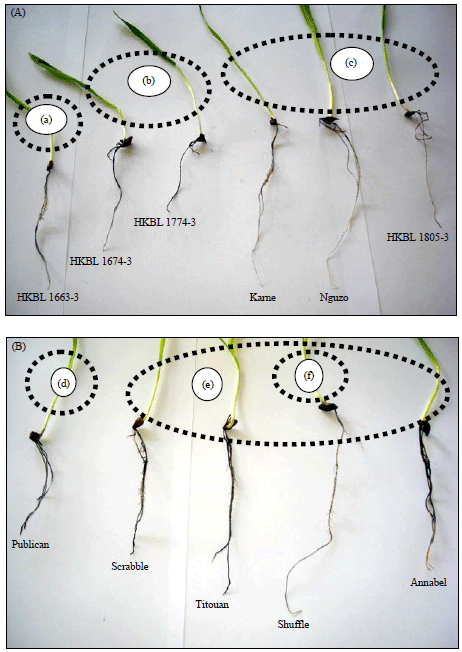 | |
| Fig. 1(A-B): | Grouping of (A) Spring barley the ‘a’, ‘b’ and ‘c’ groupings represent the major categories in terms of response to aluminium toxicity and (B) Winter barley based on degree of hematoxylin staining, the ‘d’, ‘e’ and ‘f’ represents major groups formed by genotypes under aluminium toxicity |
(a) Tolerant by fixation, (b) Sensitive, (c) Tolerant by exclusion, (d) Sensitive, (e) Tolerant by fixation, (f) Tolerant by exclusion | |
However, when the spring and winter genotypes are compared with reference to effects of Al cations on roots and the degree of staining, it is evident that the winter genotypes are more sensitive than spring genotypes (Fig. 1B).
DISCUSSION
Varying responses to aluminium toxicity by the winter and spring adapted barley indicates that the screened barley were diverse in their genetic composition hence influencing the response mechanisms to cation toxicity. In terms of net root length, relative net root growth, percent response and degree of hematoxylin staining, the observed tolerance and sensitive responses could be due varying genetic make of each genotype19. In addition, the stunted roots under the influence of aluminium toxicity which resulted into low net root length and relative net root growth could be due to interference with cell division at the root tips.
| Table 2: | Response of winter barley to aluminium toxicity in nutrient solution containing 148 μM Al3+ under laboratory conditions |
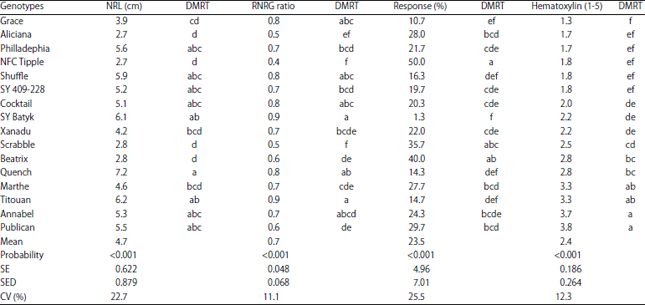 | |
NRL: Net root length, RNRG: Relative net root growth, DMRT: Duncan multiple range test | |
As a result, there was increased cell wall rigidity among the sensitive genotypes possibly due to cross-linking pectins and reduced DNA replication which increased rigidity of the double helix structure20. Varying degree of staining intensity indicates that irrespective of the winter or spring adaptation traits, the genotypes expressed different mechanisms to counter the effect of aluminium cations in the roots systems. For example, high staining intensity coupled with stunted root system in sensitive genotypes could indicate that such roots absorbed much aluminium into the root system and after absorption, the cations remained in its active and toxic state thus affecting the root development. Moreover, it is possible that some winter and spring genotypes utilized fixation and exclusion mechanisms to reduce the effects of cations on root growth and development. For instance, genotypes with low hematoxylin staining intensity could have utilized exclusion mechanism21 hence did not absorb aluminium from the nutrient solution. This also means that the aluminium concentration in the roots of varieties that used exclusion method was low.
Other than the exclusion from the root cells and organelles, some genotypes absorbed much aluminium into their root and shoot tissues and fixed hence the effect on growth and development was minimal. This means that such genotypes used fixation mechanism of aluminium tolerance to reduce the effects of aluminium toxicity. It could also mean that such genotypes had high internal tolerance to aluminium cation and therefore, despite the accumulation of much cations in their tissues, the uptake and transport of essential nutrients such as Ca, Mg, K and P as well as the activity of many enzymes and metabolic pathway for repair mechanism were less affected22. Specifically, group ‘a’ consisted of the barley genotypes that utilized fixation mechanism to immobilize the Al cation within the root tips thus no interference with root growth and development despite absorption of substantial aluminium cation. The second group ‘b’ was very sensitive to Al cation which interfered with cell division and normal physiological functions23 as evidenced by root stunting. Among the spring adapted barley such as Karne, Nguzo and HKBL 1805-3 genotypes in group ‘c’, exclusion mechanism was evidently expressed and the roots were less stained. This may imply that such genotypes have genetically triggered cell functions whose role is to exclude Al cation from being absorbed into the xylem and phloem tissues at the root tip.
With reference to RNRG and percentage response, some genotypes were less sensitive to variation in Al cation in nutrient solution. In particular, HKBL 1719-4 genotype recorded the highest RNRG of 1.0, implying that the roots treated with 148 μM and that under 0 μM Al did not differ significantly despite absorbing much Al cations into the root systems. Similarly, the same genotype had the least percentage response, a further indication that the effects of Al cation on roots were insignificant. This could mean that in such genotypes, Al cation does not affect the physiological functions such as the uptake and transport of essential nutrients, plasma membrane, membrane transport proteins and activities of enzymes among others24. However, for the winter adapted barley, the RNRG and percentage response indicate that these genotypes are more sensitive to Al cations compared to the spring adapted barley. This finding explains why most of the winter adapted barley does not perform in a wider range of agro-ecological zones in Kenya and this corresponds to the findings by other researchers. Irrespective of genetic make-up and adaptability, 3 distinct response groups namely sensitive, tolerant by fixation and tolerant by exclusion mechanisms were formed. This clearly corresponds to the previous findings which documented that tolerance to aluminium toxicity in barley monogenic and is controlled by a single dominant gene located25 in chromosome 4. For instance, when barley roots are exposed to Al cation, this gene provide a signal transduction pathway such as mitogen-activated protein kinase (MAPK) which transmits signal for stresses including Al cation toxicity26. However, it is also possible that Al sensitive genotypes such as Publican and HKBL 1674-4 do not contain such genes hence no signals are induced even if the roots are exposed to aluminium cation toxicity.
Contrary to the sensitive or tolerant responses, some barley genotypes expressed mixed responses to aluminium toxicity in terms of percentage response, RNRG and hematoxylin staining. It is possible that such genotypes express a number of genes that functions differently in terms of growth stimulation and tolerance mechanisms when exposed to aluminium toxicity27. This could also be due to the varied location of the Al tolerance gene as documented in wheat and rye where in wheat, major genes influencing Al tolerance are located on the short arm of chromosome 5A and long arms of chromosomes 2D and 4D. Therefore, being that wheat and barley are closely related, it is possible that when subjected to Al stress, the response mechanisms and variations observed were influenced by the genes expressed and the chromosomes in which the Al tolerance gene is located for each barley genotype. Based on these observations and the key findings, the study recommends the need to establish the actual signal transduction mechanisms, the genetic loci and the biochemical changes in barley under the influence of aluminium toxicity.
CONCLUSION
Spring adapted barley expressed more tolerance to aluminium toxicity compared to the winter adapted genotypes. Therefore, studies aiming at screening for tolerance to aluminium toxicity should exploit the genetic potentials of Fanaka, MALT 1, Aliciana and Grace genotypes which shows tolerance to aluminium cation toxicity.
SIGNIFICANCE STATEMENT
This study has discovered that a number of commonly grown winter and spring adapted barley genotypes in Kenya can be a good source of germplasm especially with tolerance to aluminium toxicity as the main trait of importance. The study also provides barley breeders with the fast and reliable technique that can be beneficial to screen thousands of genotypes within a short period of time. This will ensure breeders synchronize well with the fast changing soil biochemistry and chemical status and produce commercial varieties that are truly adapted to the prevailing acidic soils.
ACKNOWLEDGMENT
The authors acknowledge Regional Universities Forum for Capacity Building in Agriculture (RUFORUM) and University of Eldoret-East African Breweries Limited Collaborative Barley Research on crop improvement program for availing the research funds which made these findings possible.
REFERENCES
- Wang, J.P., H. Raman, G.P. Zhang, N.J. Mendham and M.X. Zhou, 2006. Aluminium tolerance in barley (Hordeum vulgare L.): Physiological mechanisms, genetics and screening methods. J. Zhejing Univ. Sci. B, 7: 769-787.
CrossRefDirect Link - Demirevska, K., L. Simova-Stoilova, V. Vassileva, I. Vaseva, B. Grigorova and U. Feller, 2008. Drought-induced leaf protein alterations in sensitive and tolerant wheat varieties. Gen. Applied Plant Physiol., 34: 79-102.
Direct Link - Abate, E., S. Hussien, M. Laing and F. Mengistu, 2013. Aluminium toxicity tolerance in cereals: Mechanisms, genetic control and breeding methods. Afr. J. Agric. Res., 8: 711-722.
Direct Link - Muindi, E.M., E. Semu, J.P. Mrema, P.W. Mtakwa and C.K. Gachene 2016. Soil Acidity Management by Farmers in the Kenya Highlands. J. Agric. Ecol. Res. Int., 5: 1-11.
CrossRefDirect Link - Were, J.O. and J.O. Ochuodho, 2014. Sensitivity of barley (Hordeum vulgare) to phosphate fertilizer formulations under acidic soils in Kenya. Int. J. Res. Stud. Biosci., 2: 50-59.
Direct Link - Kisinyo, P.O., C.O. Othieno, S.O. Gudu, J.R. Okalebo and P.A. Opala et al., 2013. Phosphorus sorption and lime requirements of maize growing acid soils of Kenya. Sustainable Agric. Res., 2: 116-123.
CrossRefDirect Link - Ma, J.F., S.J. Zheng, X.F. Li, K. Takeda and H. Matsumoto, 1997. A rapid hydroponic screening for aluminium tolerance in barley. Plant Soil, 191: 133-137.
CrossRefDirect Link - Maxim, P. and Z. Duþã, 1996. Aluminium tolerance of barley: I. Efficiency of in vivo procedures in estimation of genotypic differences. Roman. Agric. Res., 5: 21-35.
Direct Link - Bal, E.B.B. and A. Alkus, 2011. Determination of in vitro responses of Turkish barley cultivars to presence of aluminium. J. Anim. Plant Sci., 21: 503-507.
Direct Link - Evans, O., L. Dickson, M. Thomas, W. Beatrice and A. Joyce et al., 2012. Development of maize single cross hybrids for tolerance to low phosphorus. Afr. J. Plant Sci., 6: 394-402.
Direct Link - Cançado, G.M.A., L.L. Loguercio, P.R. Martins, S.N. Parentoni, E. Paiva, A. Borém and M.A. Lopes, 1999. Hematoxylin staining as a phenotypic index for aluminum tolerance selection in tropical maize (Zea mays L.). Theor. Applied Genet., 99: 747-754.
CrossRefDirect Link - Jaiswal, S.K., J. Naamala and F.D. Dakora, 2018. Nature and mechanisms of aluminium toxicity, tolerance and amelioration in symbiotic legumes and rhizobia. Biol. Fertil. Soils, 54: 309-318.
CrossRefDirect Link - Eekhout, T., P. Larsen and L. de Veylder, 2017. Modification of DNA checkpoints to confer aluminum tolerance. Trends Plant Sci., 22: 102-105.
CrossRefDirect Link - Gupta, B. and B. Huang, 2014. Mechanism of salinity tolerance in plants: Physiological, biochemical and molecular characterization. Int. J. Genom., Vol. 2014.
CrossRefDirect Link - Parmar, P., B. Dave, A. Sudhir, K. Panchal and R.B. Subramanian, 2013. Physiological, biochemical and molecular response of plants against heavy metals stress. Int. J. Curr. Res., 5: 80-89.
Direct Link - Bojórquez-Quintal, E., C. Escalante-Magaña, I. Echevarría-Machado and M. Martínez-Estévez, 2017. Aluminum, a friend or foe of higher plants in acid soils. Front. Plant Sci., Vol. 8.
CrossRefDirect Link - Ma, Y., C. Li, P.R. Ryan, S. Shabala and J. You et al., 2016. A new allele for aluminium tolerance gene in barley (Hordeum vulgare L.). BMC Genomics, Vol. 17, No. 1.
CrossRefDirect Link - Osawa, H. and H. Matsumoto, 2001. Possible involvement of protein phosphorylation in aluminum-responsive malate efflux from wheat root apex. Plant Physiol., 126: 411-420.
CrossRefDirect Link - Gupta, N., S.S. Gaurav and A. Kumar, 2013. Molecular basis of aluminium toxicity in plants: A review. Am. J. Plant Sci., 4: 21-37.
Direct Link