
Gehan G. Mostafa
Department of Horticulture (Ornamental Plants), Faculty of Agriculture, Beni Suef University, Egypt
Mona F. Abou Alhamd
Department of Botany, Faculty of Science, South Valley University, Qena, Egypt
International Journal of Plant Breeding and Genetics
Year: 2016 | Volume: 10 | Issue: 2 | Page No.: 110-115







ABSTRACT
Background: Chromosome doubling using colchicine has been used in plant breeding programs to produce the polyploidy plants that have superior agronomic traits over their diploid counter part. They have an increase in cell size, leading to larger reproductive and vegetative organs, increasing the production of important medicinal compounds and a higher tolerance to environmental stress. Methodology: Seeds of Celosia argentea were treated by 0, 0.01, 0.02, 0.05, 0.1 and 0.2% colchicine for 48 h to produce polyploidy plants. Stomata size and density, pollen grain size, number of chloroplast per guard cell and DNA content as well as morphological traits were used to detect the polyploidy plants. Results: Putative tetraploid plants were increased significantly in all studied traits either vegetative, flowering growth or phytochemical compound compared to diploid ones. Pollen viability and seed germination percentage were decreased in the tetraploid plants compared to the diploid ones. The DNA content in the tetraploid plant was two times more than the diploids. Other traits used to screen the tetraploid plants were found to be effective. Conclusion: Finally, colchicine is an effective method in the breeding programe of Celosia argentea.
PDF Abstract XML References Citation
Received: January 30, 2016;
Accepted: February 20, 2016;
Published: March 15, 2016
How to cite this article
Gehan G. Mostafa and Mona F. Abou Alhamd, 2016. Detection and Evaluation the Tetraploid Plants of Celosia argentea Induced by Colchicines. International Journal of Plant Breeding and Genetics, 10: 110-115.
DOI: 10.3923/ijpbg.2016.110.115
URL: https://scialert.net/abstract/?doi=ijpbg.2016.110.115
DOI: 10.3923/ijpbg.2016.110.115
URL: https://scialert.net/abstract/?doi=ijpbg.2016.110.115
INTRODUCTION
Celosia argentea is a cultivated annual plant belongs to the family Amaranthaceae. It is mainly used for flowerbeds in gardens. Celosia argentea is used as vegetable and medicinal plants. This plant is one of the main sources of pigments, proteins, calories, vitamins and minerals such as calcium, phosphorus and iron. It is also used for traiting dysenter, diarrhea, acute abdominal pain, inflamed stomach and skin eruption. It contains different compositions which occur as secondary metabolites such as alkaloid, glycosides, flavonoids, saponins, tannins and carbohydrate1-4.
Chromosome doubling using colchicine has long been used in plant breeding programs and progress in genetic study. Polyploidy plants may be found in agriculture and horticulture as they often-superior agronomic traits over their diploid counter part. For example, polyploids may have larger leaves and flowers, thicker stems and roots, darker green leaves and an increased width to length ratio of the leaves. They have an increase in cell size, leading to larger reproductive and vegetative organs, increasing the production of important medicinal compounds and a higher tolerance to environmental stress5-7.
Indirect methods have been developed for ploidy determination, cytological characteristics such as size of stomata cells, stomata density, pollen grain diameter and number of chloroplasts in guard cells can be used as indirect ways for determining polyploids. Total DNA content can also be a good indicator of ploidy level as it increases by chromosome doubling that happens in polyploidy. Therefore, optical density of extracted DNA, measured by spectrophotometry is quick and simple way to determine the ploidy change8.
This study aimed to produce tetraploid plants in Celosia argentea using colchicine treatments and evaluated their performance in vegetative and flowering growth and their content from phytochemical composition. Also, determine the ploidy induction using stomata size and density, pollen grain size, number of chloroplast per guard cell and DNA content.
MATERIALS AND METHODS
This study was carried out at the Nursery of Ornamental Plants, Faculty of Agriculture, South Valley University, Qena, Egypt.
Seeds of Celosia argentea var. spicata were treated by colchicine solutions at 0, 0.01, 0.02, 0.05, 0.1 and 0.2% for 48 h on 15 February, 2015. One hundred and twenty seeds were used for each treatment. The seeds were sown in three replications; each one contained four plastic pots. Each pot contained a mixture of clay, sand (1:1 v/v). After 2 months, the seedlings were transplanted into plastic pots containing the soil mixture of clay and sand (3:1 v/v).
All plants of the different treatments, which possess a superior growth were examined for tetraploid state.
Morphological observations: After eight month from sowing, plant height, numbers of branches and leaves per plant, leaf area, stem diameter, length of the inflorescence in the main stem and number of inflorescences per plant were determined. Open pollinated seeds from diploid and confirmed tetraploid plants were collected and seed germination percentage was examined in petri dishes.
Phytochemical determination: Chlorophyll content (SPAD unit) was determined in the leaves with SPAD apparatus as described by Yadawa9. Anthocyanin determination was conducted in the inflorescences10.
Alkaloids were determined in the leaves11. Five grams of the powdered sample was extracted with 10 mL of petroleum ether. Then, petroleum ether were removed using aspirator. One gram of the extract was suspended in 10 mL of double distilled water and the pH was adjusted to 7.6. After shaking for 1 h the suspension was centrifuged. One milliliter of the supernatant was diluted to 50 mL with phosphate buffer. The absorbance was measured spectrophotometrically at 580 ηm.
Total phenols were determined with the folin-ciocalteu assay methods12,13. One milliliter of ethanolic extract was added to 9 mL of distilled deionized water. One milliliter of folin-ciocalteu reagent was added to the mixture and shaken. After 5 min, 10 mL of 7% Na2CO3 solution was added to the mixture. After incubation for 90 min at room temperature, the absorbance was determined at 750 ηm with a UV spectrophotometer. The data was expressed as milligrams of Gallic Acid Equivalents (GAE) per gram extract.
Polyploidy determination: Putative tetraploid plants were selected based on morphological traits from different treatments for further estimation to confirm the ploidy induction.
Size and density of stomata and chloroplast number measurements: Epidermal cells of two leaves from nine diploid plants and nine putative tetraploid plants were used. Epidermal cells of plant were obtained by applying thin layer of clear nail polish on the lower leaf surface and removing it after drying the polish (a baxial side). Imprints were placed on a microscope slide and observed through the light microscope at 1000x magnification equipped with digital camera. Number of stomata, number of chloroplast, length and width of stomata were measured using a line scale implanted in the micrometer ocular8.
Pollen fertility and diameter: Fertility of the pollen (PF) was determined using acetocarmine technique4. Poorly stained and shrunken pollens were considered as sterile.
![]()
Pollen diameter was also estimated in micrometer using a line scale implanted in the micrometer ocular.
DNA content determination: Genomic DNA was extracted from young leaves using extraction kit according to manufacture's instruction (Promega, USA). The DNA content was quantified by measuring absorbance at 260 ηm wave length using UV spectrophotometer and calculated14,15.
Statistical analysis: Data of the diploid and confirmed tetraploid plants were statistically analyzed using t-test after calculation the variance homogeneity16.
RESULTS AND DISCUSSION
Table 1 presents the mean values and variances of morphological traits for both diploid and tetraploid plants. The results of t-test indicated statistically highly significant differences between diploid and tetraploid plants for all studied traits.
Gigantism growth was found in polyploidy plants compared to diploids as shown in Fig. 1. Plant height was 45.6 and 98.0 cm for diploid and tetraploid plants, respectively as shown in Table 1. Numbers of branches were 9 and 91 for diploid and tetraploid plants, respectively. Large leaf number and area were found in tetraploid (227.5 and 132, respectively) compared to (44.0 and 63.6) for diploid plants. Also, stem diameter was increased in tetroploid plant (1.37 cm) compared to diploid (0.63 cm). Number of inflorescences and length of inflorescence were increased significantly in tetraploid 17.0 and 15.0, respectively compared to diploid (5.3 and 6.0).
Colchicine inhibits the formation of spindle fibers and temporarily arrest mitosis at the anaphase stage. At this point, the chromosomes are replicated but cell division do not occur resulting in polyploidy cells. Increasing ploidy often results in increased cell size that in turn results in thicker, broader leaves and larger flowers7.
Seeds of both diploid and putative tetraploid were collected and germinated on petri dishes. Seed germination percentage was significantly decreased from 69.53-16.35% when the ploidy level turned from diploid to tetraploid as shown in Table 1. This may be due to their thicker seed coat and seedling emergence strength17 or may be due to acute chromosomal damage, delay in the onset of mitosis, chromosomal aberrations induced enzyme activity such as catalase and lipase and hormonal activity resulted in reduced germination18.
Darker leaves and higher chlorophyll content in tetraploid were detected (41.6) compared to diploids (34.1). The dark green color is the result of bigger cells and more chlorophyll content19.
For medicinal plants, polyploids are usually more valuable because they exhibit increased biomass and content of effective compound19,20. Alkaloid content was 4.89-15.37 mg g–1 for diploid and tetraploid plants respectively as shown in Table 2. Phenols also was increased significantly in tetraploid plants (15.0 mg g–1) compared to 8.1 mg g–1 in diploid. Anthocyanin content as an antioxidant substance was also increased in the tetraploid plants (22.2 mg g–1) compared to diploids (10.6 mg g–1).
Figure 2 shows that stomata size and density. Length and width of the stomata in putative morphologically tetraploid plants were measured as shown in Table 3. The obtained results showed that, stomata size was highly significant increased compared to diploids.
| Table 1: | Comparison between diploid and tetraploid plants of Celosia argentea concerning to plant height, number of branches and leaves, leaf area, stem diameter, length of inflorescence, number of inflorescences per plant and seed germination percentage |
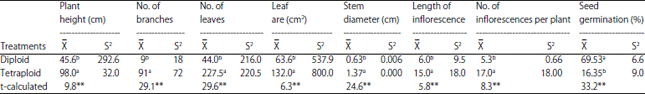 | |
| **Highly significant at p = 0.01, : Mean values and S2: Variance | |
 | |
| Fig. 1(a-b): | Photograph showing, (a) Tetraploid plant of Celosia argentea produced by colchicine treatments compared to (b) Diploid plants |
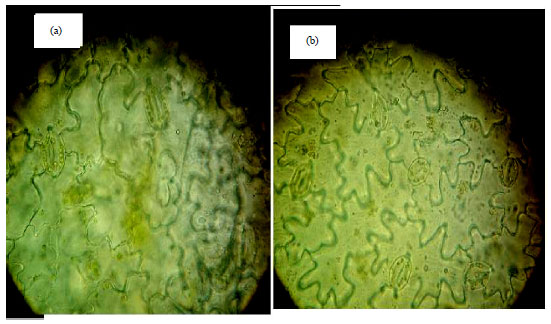 | |
| Fig. 2(a-b): | Photograph showing stomata size and density in the (a) Tetraploid plant of Celosia argentea produced by colchicine treatments compared to (b) Control (diploid) plants |
| Table 2: | Comparison between diploid and tetraploid plants of Celosia argentea concerning to chlorophyll content (SPAD unit), anthocyanin content, alkaloid and phenols content |
 | |
| NS: Not significant at p = 0.01, **Significant at p = 0.01, : Mean values and S2: Variance | |
Mean values were 100.0 and 147.5 μm for diploid and tetraploid respectively with respect to stomata length and 55 and 96.6 μm for stomata width.
| Table 3: | Comparison between diploid and tetraploid plants of Celosia argentea concerning to number of stomata/microscopic field, number of chloroplast per guard cell, length and width of stomata (μm), pollen viability percentage, pollen diameter (μm) and total DNA content (μg mL–1) |
 | |
| **Highly significant at p = 0.01, : Mean values and S2: Variance | |
Stomata size and number can be an indicator of polyploidy levels and it has been used in different plant types for determining ploidy levels7,8. The counted stomata number in the microscope field was 24 for diploids and 13.3 in the polyploidy plants as shown in Table 3.
Larger stomata cells and decrease in stomata number were found in tetraploids in many studies research19. They stated that, stomata size and counting were identified as the most effective criteria of polyploidy, due to easier and economical application. When polyploidy level increased, plant cells became larger, while the number of stomata per area decreased.
Stomata measurement was used with morphological changes as pre selection method after colchicine treatment21. Number of chloroplast per guard cell was highly significantly greater in the tetraploid plant (12.8) than diploid plants (7.6). No significant decrease were found in the tetraploid plants with respect to pollen viability (88%) compared to diploid plant (94.3%) as shown in Table 3. The chromosomal aberrations, some genetic and physiological changes might have caused pollen sterility18. Pollen diameter in the putative tetraploid was significantly increased (286 μm) compared to diploid ones (131.66 μm). Greater number of chloroplasts per guard cell and pollen diameter were found in the polyploidy plant rather than diploid in the other studies7,22.
Total DNA content was increased significantly in the tetraploid plants (30 μg mL–1) compared to (14 μg mL–1) in the diploid ones as shown in Table 3. Many colchicine studies proved by treatment that the tetraploid cells contained DNA almost two times more than diploid ones15. Total DNA content of plants increases as a result of chromosome doubling that happens in polyploidization. This can be used as an indicator for ploidy determination8.
Colchicine has been widely used for chromosome doubling in a variety of plant species leading to the production of novel germplasm that can be used as bridging materials or as direct commercial cultivars with improving characteristics15.
CONCLUSION
Tetraploid Celosia argentea plants with bigger vegetative and flowering growth and with highly phytochemical compound content were obtained using colchicine treatments. Putative polyploidy was detected through examination of stomata size, stomata number, pollen grain size, number of chloroplast per guard cells and confirmed with DNA content.
REFERENCES
- Akinyemi, S.O.S. and H. Tijani-Eniola, 2000. Effects of cassava density on productivity of plantain and cassava intercropping system. Fruits, 55: 17-23.
Direct Link - Patel, B.A., P.U. Patel and R.K. Patel, 2010. Physicochemical and phytochemical investigations of seeds of Celosia argentea Linn. Int. J. Pharmaceut. Applied Sci., 1: 124-126.
Direct Link - Abubakar, A., A.O. Falusi, O.A.Y. Daudu, A.O. Oluwajobi, M.C. Dangana and D.R. Abejide, 2015. Mutagenic effects of sodium azide and fast neutron irradiation on the cytological parameters of M2 Lagos spinach (Celosia argentea var cristata L.). World J. Agric. Res., 3: 107-112.
CrossRefDirect Link - Kermani, M.J., V. Sarasan, A.V. Roberts, K. Yokoya, J. Wentworth and V.K. Sieber, 2003. Oryzalin-induced chromosome doubling in Rosa and its effect on plant morphology and pollen viability. Theoret. Applied Genet., 107: 1195-1200.
CrossRefDirect Link - Shao, J., C. Chen and X. Deng, 2003. In vitro induction of tetraploid in pomegranate (Punica granatum). Plant Cell Tissue Org. Cult., 75: 241-246.
CrossRefDirect Link - Grouh, M.S.H., H. Meftahizade, N. Lotfi, V. Rahimi and B. Baniasadi, 2011. Doubling the chromosome number of Salvia hains using colchicine: Evaluation of morphological traits of recovered plants. J. Med. Plants Res., 5: 4892-4898.
Direct Link - Moghbel, N., M.K. Borujeni and F. Bernard, 2015. Colchicine effect on the DNA content and stomata size of Glycyrrhiza glabra var. glandulifera and Carthamus tinctorius L. cultured in vitro. J. Genet. Eng. Biotechnol., 13: 1-6.
CrossRefDirect Link - Yadawa, U.L., 1986. A rapid and nondestructive method to determine chlorophyll in intact leaves. HortScience, 21: 1449-1450.
Direct Link - Fuleki, T. and F.J. Francis, 1968. Quantitative methods for anthocyanins. 1. Extraction and determination of total anthocyanin in cranberries. J. Food Sci., 33: 72-77.
CrossRefDirect Link - Singleton, V.L., R. Orthofer and R.M. Lamuela-Raventos, 1999. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In: Methods in Enzymology, Burslem, G.L. (Ed.), Academic Press, Cambridge, Massachusetts, ISBN: 9780121822002, pp: 152-178.
CrossRefDirect Link - Salem, M.Z.M., Y.M. Gohar, L.M. Camacho, N.A. El-Shanhorey and A.Z.M. Salem, 2013. Antioxidant and antibacterial activities of leaves and branches extracts of Tecoma stans (L.) Juss. ex Kunth against nine species of pathogenic bacteria. Afr. J. Microbiol. Res., 7: 418-426.
Direct Link - Abou Elyazid, D.M. and A.R. El-Shereif, 2014. In vitro induction of polyploidy in Citrus reticulata Blanco. Am. J. Plant Sci., 5: 1679-1685.
CrossRefDirect Link - Sung, J.M. and K.Y. Chiu, 1995. Hydration effect on seedling emergence strength of watermelon seeds differing in ploidy. Plant Sci., 110: 21-26.
CrossRefDirect Link - Roychowdhury, R. and J. Tah, 2011. Chemical mutagenic action on seed germination and related agro-metrical traits in M1 Dianthus generation. Curr. Bot., 2: 19-23.
Direct Link - Tulay, E. and M. Unal, 2010. Production of colchicine induced tetraploids in Vicia villosa roth. Caryologia, 63: 292-303.
CrossRefDirect Link - Gao, S.L., D.N. Zhu, Z.H. Cai and D.R. Xu, 1996. Autotetraploid plants from colchicine-treated bud culture of Salvia miltiorrhiza Bge. Plant Cell Tissue Organ Cult., 47: 73-77.
CrossRefDirect Link - Jadrna, P., O. Plavcova and F. Kobza, 2010. Morphological changes in colchicine-treated Pelargonium × hortorum L.H. Bailey greenhouse plants. Horticult. Sci., 37: 27-33.
Direct Link - Gu, X.F., A.F. Yang, H. Meng and J.R. Zhang, 2005. In vitro induction of tetraploid plants from diploid Zizyphus jujuba Mill. cv. Zhanhua. Plant Cell Rep., 24: 671-676.
CrossRefDirect Link