
M. A. Abdel-Moneam
Department of Agronomy, Faculty of Agriculture, Mansoura University, Egypt
International Journal of Plant Breeding and Genetics
Year: 2014 | Volume: 8 | Issue: 3 | Page No.: 153-163







ABSTRACT
Six genotypes of flax representing different agronomic characters were crossed in a half-diallel model in 2009/2010 winter season. The six parents and their 15 F1 crosses were evaluated in 2010/2011 and 2011/2012 seasons to study the combining ability and mode of gene action for yield and its components traits of flax. The results indicated clearly that mean squares of genotypes, general and specific combining ability were highly significant for all studied traits in both seasons and their combined data, except general combining ability for capsules No. plant-1 in both seasons and No. of basal branches plant-1 in the second season were insignificant. The GCA/SCA ratio was more than unity for days to the first flower, plant height and technical length, indicating that these traits are predominantly controlled by additive gene action. Whereas, GCA/SCA ratio was less than unity for No. of basal branches plant-1, No. of capsules plant-1, 1000-seed weight, seed yield plant-1 and straw yield plant-1, indicating that these traits are mainly controlled by non-additive gene action. The best general combiners were P1 (S2) for straw yield, plant height and technical length; P5 (Hermis) for seed and straw yields; P4 (Bleinka) for seed yield plant-1 and 1000-seed weight. Eight crosses and nine crosses showed significant or highly significant and positive SCA effects for seed yield plant-1 and straw yield plant-1, respectively, indicating that these crosses were the best combinations for increasing seed and straw yields.
PDF Abstract XML References Citation
Received: January 23, 2014;
Accepted: April 24, 2014;
Published: June 25, 2014
How to cite this article
M. A. Abdel-Moneam, 2014. Diallel Cross Analysis for Yield and its Related Traits in Some Genotypes
of Flax (Linum usitatissimum L.). International Journal of Plant Breeding and Genetics, 8: 153-163.
DOI: 10.3923/ijpbg.2014.153.163
URL: https://scialert.net/abstract/?doi=ijpbg.2014.153.163
DOI: 10.3923/ijpbg.2014.153.163
URL: https://scialert.net/abstract/?doi=ijpbg.2014.153.163
INTRODUCTION
Flax (Linum usitatissimum L.) is considered the most important past fiber crop in Egypt since several thousand years ago. Choice of parents for crossing is considered an important step in any plant breeding program aimed at improving yield and related attributes. Combining ability analysis is an important tool for the selection of desirable parents together with the information regarding to the nature and magnitude of gene effects controlling quantitative traits. It is important to accurately estimate the magnitude and relative proportion of the various components of genetic variance in order to understand the underlying type of gene action that controls the trait of interest, for example General Combining Ability (GCA) variance is a measure of additive effects of genes and of additive x additive epistatic interaction, while Specific Combining Ability (SCA) variance is measure of dominance and epistatic types of gene action. Many researchers studied the combining ability in flax, i.e., Patil et al. (1997), Foster et al. (1998), Abo El-Zahab and Abo-Kaied (2000), Abo-Kaied (2002) and El-Kady and Abo-Kaied (2010), they found that additive genetic variance had more important role in the inheritance of straw yield, plant height, technical stem length and 1000-seed weight. On the other hand, non-additive variance had an important role in the inheritance of No. of basal branches plant-1, seed yield plant-1 and capsules number plant-1 as reported by Shehata and Comstock (1971), Patil and Chopde (1981), Zahana (2006), Abo-Kaied et al. (2007) and El-Kady and Abo-Kaied (2009). While, Mishra et al. (2013) reported that GCA/SCA were less than 1 revealed the importance of non-additive gene action in the inheritance of all seed yield and its components traits.
Mohammadi et al. (2010) showed that additive gene actions had greater effects on inheritance of days to flowering, plant height, primary branches per plant and 1000-seed weight. However, number of capsules per plant was largely controlled by dominance genetic effects, whereas both additive and non-additive gene actions were important in genetic control of seed yield plant-1.
Therefore, the present investigation was designed to estimate the combining ability effects and the mode of gene action in the inheritance of some traits in six parental foreign flax genotypes and their 15 F1 crosses during two growing seasons.
MATERIALS AND METHODS
The genetic materials used in this investigation as parents included six foreign genotypes of flax. The parent's names and their origin are presented in Table 1.
In 2009/2010 winter season, the six parental genotypes were sown at various dates in order to overcome the differences in flowering time. All possible cross combinations excluding reciprocals were made among the six genotypes, giving 15 F1 crosses. The parents and their F1 crosses were evaluated in the Farm of Agron, Department of Agronomy, Faculty of Agriculture, Mansoura University, Egypt during two growing seasons, 2010/2011 and 2011/2012. The experiments were laid out in a randomized complete block design with three replications. Each plot consisted of one row for parents and its F1 crosses. Rows were 3 m long and spaced 20 cm apart. Single seeds were hand drilled at 5 cm spacing within row. All other cultural practices were applied as recommended for flax cultivation. At harvest, individual guarded plants were taken at random from each row; 10 plants from each parent and F1 per replication for recording the studied traits.
Studied traits: Days to first flower (day), plant height (cm), technical length (cm), No. of basal branches plant-1, No. of capsules plant-1, 1000-seed weight (g), seed yield plant-1 (g) and straw yield plant-1 (g).
Statistical analysis: The data obtained for each trait was analyzed on plot mean basis in both parents and F1 generation. All obtained data was subjected to the statistical analysis of the randomized complete block design to test the differences among various genotypes under each season according to Snedecor and Cochran (1980). Combined analysis was performed between the two seasons to indicate the seasonal year effects.
| Table 1: | Flax genotypes names and their origin |
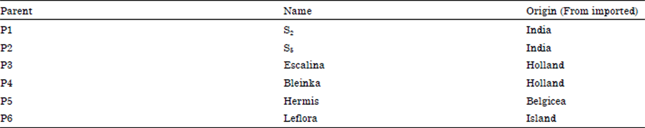 | |
| Table 2: | Analysis of variance from method 2 model 1 and expected mean squares for combining ability analysis |
 | |
| Me: Error mean squares of the main randomized complete block design divided by number of replications (Me: Me/r), P: Number of parents | |
Treatments were compared using the Least Significant Differences values (LSD) at 5% probability according to Gomez and Gomez (1984). An ordinary analysis using Griffing (1956) method 2 model 1 (fixed effects) was applied to estimate General Combining Ability (GCA) and Specific Cambining Ability (SCA) effects, as shown in Table 2.
RESULTS AND DISCUSSION
Analysis of variance for the studied traits under both seasons and their combined data are presented in Table 3. The results indicated clearly that mean squares of genotypes were highly significant for all studied traits in both seasons and their combined data. These results reflected an indication to the presence of real differences between the studied genotypes for these traits. General (GCA) and specific (SCA) combining ability mean squares presented in Table 3 were highly significant for all studied traits in both seasons and their combined data, except general combining ability for No. of capsules plant-1 in both seasons and No. of basal branches plant-1 in the second season were insignificant.
The GCA/SCA ratio was more than unity for each of days to first flower, plant height and technical length in both seasons and their combined data (Table 3), indicating that these traits are predominantly controlled by additive gene action. Therefore, it could be concluded that selection procedures based on the accumulation of additive gene effect would be more effective in the early segregating generations for improving these traits. These results are in general agreement with those previously reported by Foster et al. (1998), Abo El-Zahab and Abo-Kaied (2000), Abo-Kaied (2002), El-Kady and Abo-Kaied (2010) and Mohammadi et al. (2010). On the other hand, GCA/SCA ratio was less than unity for each of No. of basal branches plant-1, No. of capsules plant-1, 1000-seed weight, seed yield plant-1 and straw yield plant-1 traits in both seasons and their combined data (Table 3), indicating that these traits are mainly controlled by non-additive gene action. Therefore, it could be concluded that hybridization procedures based on dominance gene effect would be more effective for improving these traits. Similar results in general reported previously by Shehata and Comstock (1971), Patil and Chopde (1981), Zahana (2006), Abo-Kaied et al. (2007), El-Kady and Abo-Kaied (2009) and Mishra et al. (2013).
Mean performance of genotypes: Data in Table 4 indicated that the P4 (Bleinka) and crosses P2xP6, P3xP5 and P4xP5 were the earliest in flowering in both seasons and their combined data. P1 (S2) and P3 (Escalina) and crosses P1x3, P1x4 and P2x3 gave the highest values of plant height in both seasons and their combined data. The parent P1 (S2) and crosses P1x3 and P1x5 gave the highest values of technical length in both seasons and their combined data. Parents P4 (Bleinka) and P5 (Hermis) and crosses P2x4, P2x3, P1x3 and P5x6 gave the highest values for No. of basal branches plant-1 in both seasons and their combined data.
| Table 3: | Mean squares from analysis of variance, for general and specific combining abilities for all studied traits in the 1st and 2nd seasons and their combination |
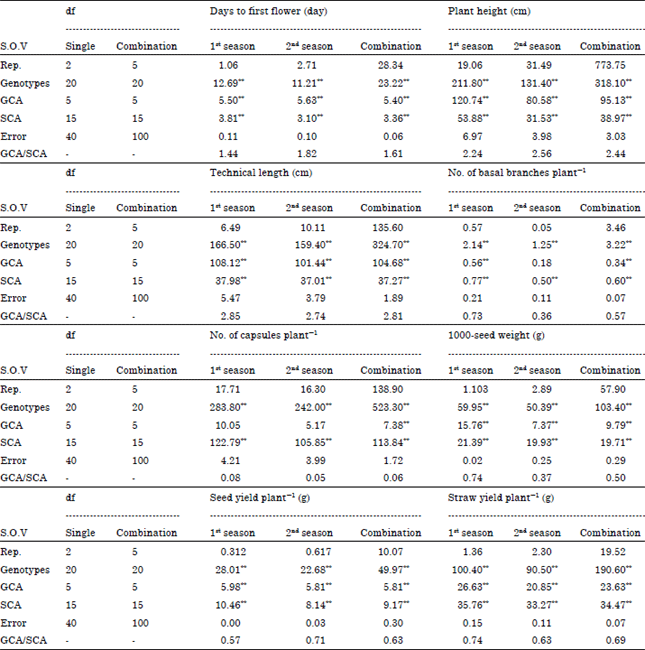 | |
| *,**Significant at 0.05 and 0.01 levels of probability, respectively | |
Parents P1 (S2) and P6 (Leflora) and crosses P2x4, P3x4, P1x5, P4x5 and P5x6 gave the highest values for No. of capsules plant-1 in both seasons and their combined data. The parent P6 (Leflora) and cross P3xP6 gave the highest values of 1000-seed weight in both seasons and their combined data. The parent P5 (Hermis) and cross P4x5 gave the highest values for seed yield plant-1 in both seasons and their combined data. The parents P1 (S2) and P2 (S5) and the crosses P1x2 and P4x5 gave the highest values of straw yield plant-1 in both seasons and their combined data, as shown in Table 4.
| Table 4: | Mean performance of flax genotypes (parents and their 15 F1 crosses) for all studied traits in the 1st and 2nd seasons and their combination |
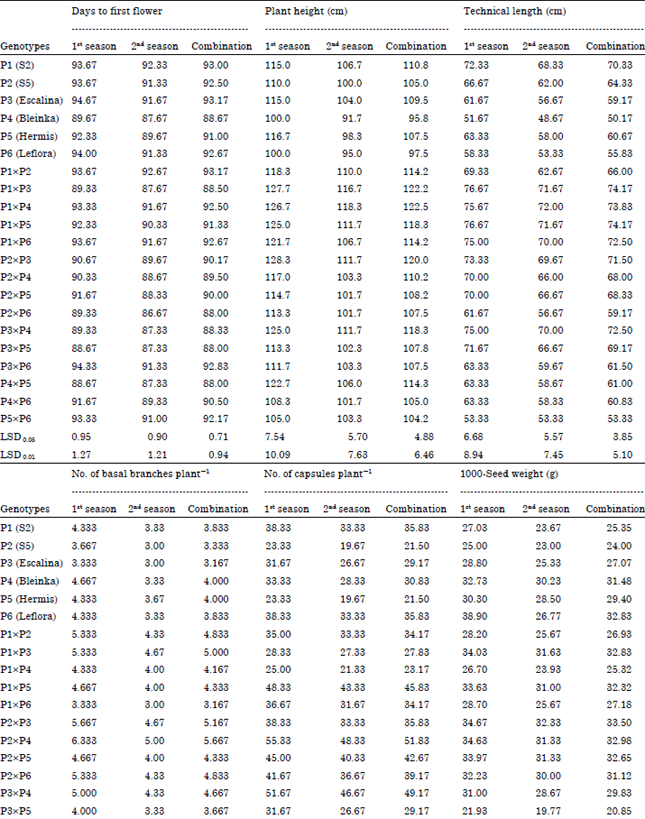 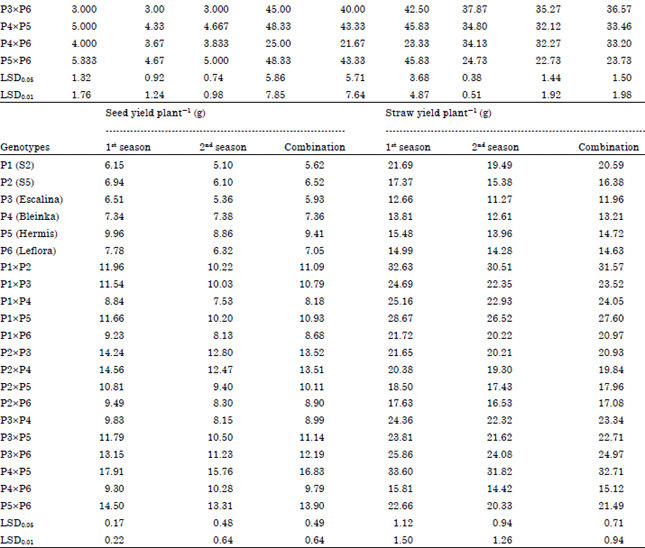 | |
Estimates of General Combining Ability effects (GCA): Data in Table 5 indicated that the parents P4 (Bleinka) and P5 (Hermis) showed high significant and negative GCA effects for days to the first flower in both seasons and their combined, indicating that these parents could be considered as good combiners for earliness. For plant height and technical length, P1 (S2) and P3 (Escalina) showed high significant and positive GCA effects in both seasons and their combined, indicating that these parents could be considered as good combiners for tallness of plant height and technical length. For No. of basal branches plant-1, P2 (S5) gave significant and highly significant positive GCA effects in the first season and combined, respectively, indicating that this parent could be considered as good combiner for this trait. For No. of capsules plant-1, P4 (Bleinka) and P6 (Leflora) showed significant positive GCA effects in combined data, indicating that these parents could be considered as good combiners for this trait. For 1000-seed weight, P4 (Bleinka) gave highly significant positive GCA effects in the two seasons and combined, indicating that this parent could be considered as good general combiner for increasing the weight of seeds. For seed yield plant-1, P4 (Bleinka) and P5 (Hermis) showed high significant and positive GCA effects in both seasons and their combined, indicating that these parents could be considered as good combiners for increasing seed yield.
| Table 5: | Estimates of GCA effects of six parents flax genotypes for studied the traits in the 1st and 2nd seasons and their combination |
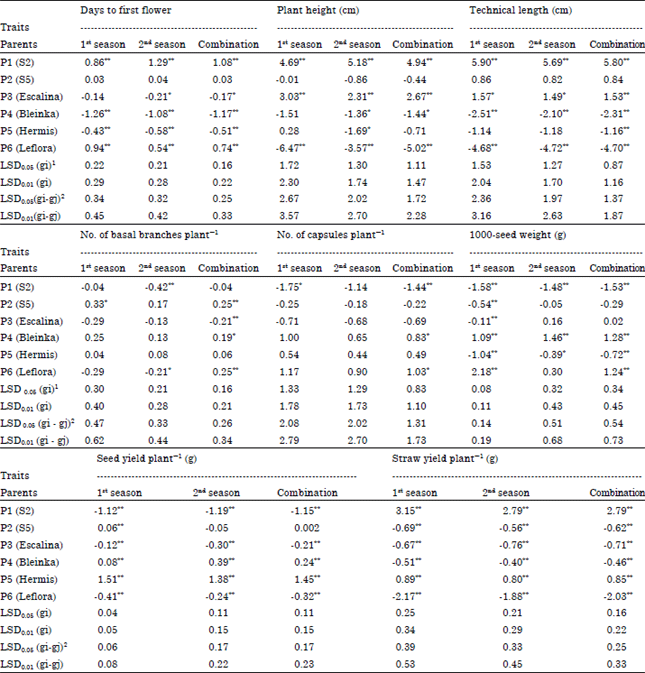 | |
| *,**Significant at 0.05 and 0.01 level of probability, respectively, 1Least significant difference for a GCA effect, 2Least significant difference for the difference between estimates of GCA effects | |
While for straw yield plant-1, P1 (S2) and P5 (Hermis) showed high significant and positive GCA effects in both seasons and their combined, indicating that these parents could be considered as good combiners for increasing straw yield. So, these parents could be used as parents for the development of desirable hybrids.
| Table 6: | Estimates of SCA effects of 15 F1 single crosses flax genotypes for the studied traits in 1st and 2nd seasons and their combination |
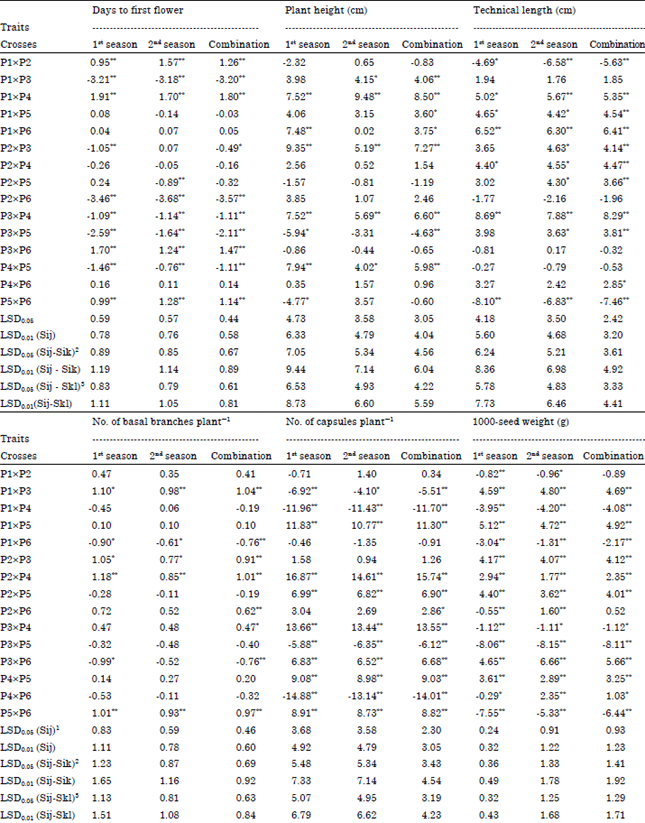 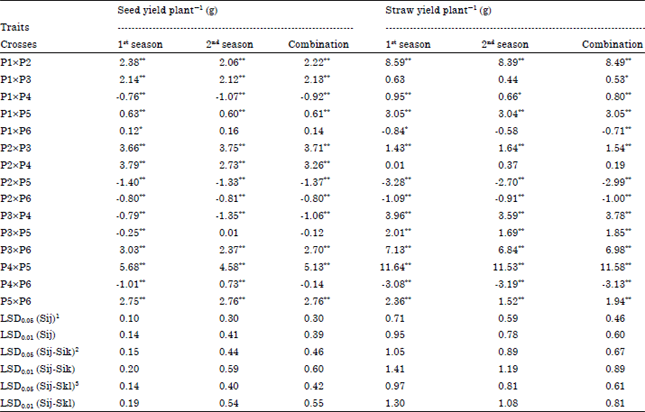 | |
| *,**Significant at 0.05 and 0.01 level of probability, respectively, 1Least significant difference for a SCA effect, 2Least significant difference for the difference between two SCA effects for a common parent, 3Least significant difference for the difference between two SCA effects for a non-common parent | |
These results are in agreement with those obtained by Abo El-Zahab and Abo-Kaied (2000), Abo-Kaied (2002), Zahana (2006), Abo-Kaied et al. (2007), El-Kady and Abo-Kaied (2009), El-Kady and Abo-Kaied (2010) and Mishra et al. (2013), where they observed significant and positive and negative GCA effects for these traits in their respective studies.
Once the best combiners are identified, they can be crossed together to obtain the promising hybrid combinations. In recurrent selection schemes, parents possessing high combining ability can be intercrossed to accumulate desirable alleles within base populations (Christie and Shattuck, 1992).
Estimates of Specific Combining Ability effects (SCA): Data in Table 6 indicated that, five crosses i.e., P1xP3, P2xP6, P3xP4, P3xP5 and P4xP5 showed high significant and negative SCA effects (desirable direction) for days to first flower trait in both seasons and their combined, indicating that these crosses were the best combinations for earliness. For plant height, four crosses viz., P1xP4, P2xP3, P3xP4 and P4xP5 showed significant or highly significant and positive SCA effects (desirable direction) in both seasons and their combined indicating that these crosses were the best combinations for tallness. For technical length, crosses P1xP4, P1xP5, P1xP6, P2x P4 and P3xP4 gave significant or highly significant and positive SCA effects in both seasons and their combined, indicating that these crosses were the best combinations for this trait. For No. of basal branches plant-1, four crosses i.e., P1xP3, P2xP3, P2xP4 and P5xP6 showed significant or highly significant and positive SCA effects in both seasons and their combined indicating that these crosses were the best combinations for this trait. For No. of capsules plant-1, seven crosses i.e., P1xP5, P2xP4, P2xP5, P3xP4, P3xP6, P4xP5 and P5xP6 gave highly significant and positive SCA effects in both seasons and their combined indicating that these crosses were the best combinations for this trait. For 1000-seed weight, seven crosses viz., P1xP3, P1xP5, P2xP3, P2xP4, P2xP5, P3xP6 and P4xP5 gave highly significant and positive SCA effects in both seasons and their combined indicating that these crosses were the best combinations for increasing the weight of seeds. For seed yield plant-1, eight crosses i.e., P1xP2, P1xP3, P1xP5, P2xP3, P2xP4, P3xP6, P4xP5 and P5xP6 showed highly significant and positive SCA effects in both seasons and their combined indicating that these crosses were the best combinations for increasing seed yield. For straw yield plant-1, nine crosses viz., P1xP2, P1xP4, P1xP5, P2xP3, P3xP4, P3xP5, P3xP6, P4xP5 and P5xP6 showed significant or highly significant and positive SCA effects in both seasons and their combined, indicating that these crosses were the best combinations for increasing straw yield. It is worthy to note that most of superior F1 crosses in their SCA effects for seed and straw yields and most of yield components traits include at least one of their parents of high GCA effects for the same traits. These results are in general agreement with those previously reported by Abo El-Zahab and Abo-Kaied (2000), Abo-Kaied (2002), Zahana (2006), Abo-Kaied et al. (2007), El-Kady and Abo-Kaied (2009), El-Kady and Abo-Kaied (2010) and Mishra et al. (2013), where they observed significant positive and negative SCA effects for these traits in their respective studies.
REFERENCES
- Christie, B.R. and V.I. Shattuck, 1992. The diallel cross: Design, analysis and use for plant breeders. Plant Breed. Rev., 9: 9-36.
CrossRef - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wily and Sons, Hoboken, New Jersey, ISBN: 9780471870920, Pages: 704.
Direct Link - Griffing, B., 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci., 9: 463-493.
Direct Link - Foster, R., H.S. Pooni and I.J. Mackay, 1998. Quantitative analysis of Linum usitatissimum crosses for dual-purpose traits. J. Agric. Sci., 131: 285-292.
Direct Link - Mishra, R., S. Marker, V. Bhatnagar and D. Mahto, 2013. Combining ability and heterosis for seed yield and its component in linseed (Linum usitatissimum L.). Adv. Life Sci., 2: 44-47.
Direct Link - Mohammadi, A.A., G. Saeidi and A. Arzani, 2010. Genetic analysis of some agronomic traits in flax (Linum usitatissimum L.). Aust. J. Crop Sci., 4: 343-352.
Direct Link - Patil, V.D. and P.R. Chopde, 1981. Combining ability analysis over environments in diallel crosses of linseed (Linum usitatissimum L.). Theor. Applied Genet., 60: 339-343.
CrossRef - Shehata, A.H. and V.E. Comstock, 1971. Heterosis and combining ability estimates in F2 flax populations as influenced by plant density. Crop Sci., 11: 534-536.
CrossRef