
T. Sudhakar
Madhusmita Panigrahy
M. Lakshmanaik
A. Prasad Babu
C. Surendhar Reddy
K. Anuradha
B.P Mallikarjuna Swamy
N. Sarla
International Journal of Plant Breeding and Genetics
Year: 2012 | Volume: 6 | Issue: 2 | Page No.: 69-82







ABSTRACT
Introgression lines (IL) from crosses between elite cultivars and wild species are an important resource for mapping quantitative trait loci (QTLs) for yield. In this study, 25 KMR3-O. rufipogon ILs, from an advanced back cross population, were selected for high grain yield in field trials and characterized in detail for yield and related traits from seed to seed. The aim was to dissect the basis of high yield in the entire life cycle to identify common features in high yielding ILs. This study indicated that each IL has a different strategy or a combination of different yield traits to attain high yield. High yield in these ILs was associated with early flowering, high number of tillers, panicles, grains, seed weight per panicle and seed density. Seed germination rate, leaf SPAD chlorophyll meter value, plant height, panicle length and 100 seed weight did not have much impact on yield. Highest significant positive correlation was observed between number of tillers and number of productive tillers (0.98) as expected. A highest significant negative correlation was observed between seed density and seed compactness (-0.95). The new negative correlation between seed germination rate and leaf SPAD chlorophyll meter value in the top 10 ILs was supported by a non significant negative correlation (-0.17) in BC3F4 IL population and deserves further study. Restricting to a specific ideotype in selection may be counterproductive in rice breeding. These ILs are being used in rice improvement and to identify favorable genes from O. rufipogon for increasing yield.
PDF Abstract XML References Citation
Received: September 12, 2011;
Accepted: November 02, 2011;
Published: January 06, 2012
How to cite this article
T. Sudhakar, Madhusmita Panigrahy, M. Lakshmanaik, A. Prasad Babu, C. Surendhar Reddy, K. Anuradha, B.P Mallikarjuna Swamy and N. Sarla, 2012. Variation and Correlation of Phenotypic Traits Contributing to High Yield in KMR3-Oryza rufipogon Introgression Lines. International Journal of Plant Breeding and Genetics, 6: 69-82.
DOI: 10.3923/ijpbg.2012.69.82
URL: https://scialert.net/abstract/?doi=ijpbg.2012.69.82
DOI: 10.3923/ijpbg.2012.69.82
URL: https://scialert.net/abstract/?doi=ijpbg.2012.69.82
INTRODUCTION
Selection for yield and improving rice yield is most challenging in plant breeding as yield is the most dynamic trait and is influenced by various environmental and genetic factors (Sreedhar et al., 2011; Pimsaen et al., 2010; Christopher and Sangodoyin, 2011) and agronomic characters (Mobasser et al., 2007; Sadeghzadeh and Alizadeh, 2005; Asif et al., 2003). In addition, deteriorating environment brings down yield continuously. On the other hand, introgression lines from wild rice (Tian et al., 2006) and hybrids (Akinwale et al., 2011) offer tremendous potential for increasing yield and adaptability. Wild species of rice are potential reservoirs of useful QTLs / genes for enhancing yield (Tanksley and McCouch, 1997; Swamy and Sarla, 2008) and useful introgression lines have been identified (Sarla et al., 2009). It was found that during domestication process from wild to cultivated rice, 40% alleles were lost leading to lower genetic diversity of cultivated rice (Tian et al., 2006). Exploitation and utilization of the favorable alleles of wild rice that are lost or weakened in cultivated rice might help to overcome the current yield plateau and broaden the genetic base of popular varieties (Tanksley and McCouch, 1997; Gur and Zamir, 2004; Swamy and Sarla, 2008). It is considered easier to improve mega varieties with introgressions from other accessions rather than creating new cultivars whose adaptability is not known, since the mega varieties are well adapted, farmer accepted, popular with good yield and grain quality (Mackill et al., 2007; Prasad et al., 2001).
Introgression lines are homozygous lines with small chromosomal segments from the donor into the recurring parent by consecutive backcrossing and selfing (Tian et al., 2006). Two yield QTLs, yld 1.1 and yld 2.1 which helped increase yield by 18%, were identified in a Malaysian accession of O. rufipogon (Xiao et al., 1996, 1998). This accession was used as a donor in many studies and using different recipients resulting in identification of many QTLs for yield and its related traits (Xiong et al., 1999; Moncada et al., 2001; Septiningsih et al., 2003; Thomson et al., 2003; McCouch et al., 2007). Using introgression lines several authors have demonstrated that ILs are useful genetic material for improving various agronomic traits in popular varieties, identification of new genes and for map-based cloning of QTLs (Xie et al., 2008; Rahman et al., 2007; Yoon et al., 2006). Ideotype breeding has been advocated specially to obtain high yield from indica x japonica crosses (Peng et al., 2008). Tremendous variability exists in the wild species or arises de novo when crossing elite cultivars with the wild accessions. It is important to know which yield related trait or yield component can help to enhance yield and up to what extent but this has not been analyzed in introgression lines. It would be useful if guidelines for selecting key traits of importance can be developed for backcross derivatives of elite x wild crosses. This paper reports characterization and analysis of yield and related traits from seed to seed in a set of 25 high yielding KMR3-O. rufipogon ILs (Marri et al., 2005; Babu et al., 2009). The objective of the study was to verify if there is a typical high yielding ideotype or there are common yield enhancing traits across introgression lines. It emerges that different introgression lines increase yield by different phenotypic strategies thus bringing to question the ideotype concept when considering introgressions from wild species.
MATERIALS AND METHODS
Selection of plant material: The 25 introgression lines used in this study originated from the mapping population IR58025A/O. rufipogon//IR58025B///IR58025B////KMR3 test cross progeny in which a major effect yield-enhancing QTL yld 2.1 was identified (Marri et al., 2005). In order to start a marker assisted backcross program to transfer this QTL to the recurrent parent KMR3, two plants, namely #26 and #105 were identified as having the O. rufipogon loci for the yld2.1 QTL flanking markers RM262 and RM263. In BC1 (among 215 plants) and BC2 (among 406 plants) about 30% of plants had both the flanking markers RM262 and RM263 from O. rufipogon. Plants #26 and #105 were crossed with KMR3 to give 215 BC1F1 plants. In this population, genotypic selection was done using flanking markers for yld 2.1. Selected BC1F1 plants were backcrossed with KMR3 to give 406 BC2F1 plants. Selected BC2F1 plants with flanking markers for yld 2.1 were backcrossed again with KMR3 to obtain BC3F1 population of 504 plants which were selfed to obtain BC3F2 population (Babu et al., 2009). From this population 60 lines were selected for various yield related traits such as tiller number, number of productive tillers, panicle length, spikelets per panicle. The 60 families were field evaluated in RBD design for phenotype with KMR3 as check during 2009.
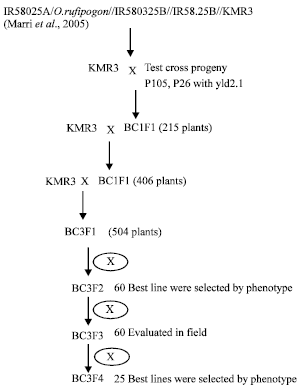 | |
| Fig. 1: | Flow diagram of the breeding program for development of KMR3-O. rufipogon introgression lines |
The highest yielding 25 lines were selected for yield related traits from these 60 lines and field evaluated in next season also. Flow diagram explaining the method for development of the ILs is described in Fig. 1.
Phenotypic evaluation of mapping population: BC3F3 plants derived from the mapping population were planted with a plant to plant distance of 15 and 20 cm between rows in an experimental plot at DRR (Directorate of Rice Research) Hyderabad, India during Kharif (wet) season in 2008. The 25 BC3F4 families were selected from 60 BC3F3 families grown under irrigated conditions and analyzed for yield-related traits during Rabi (dry) 2009 (5 rows each) and Kharif (2009) (20 rows each) in DRR field. A randomized block design was followed in two replications with KMR3 as check variety. Phenotypic observations were recorded for yield and related traits from five middle plants of each of the families in both seasons. In addition plot yield was also taken for each family.
Analysis of yield-related traits: Seed weight (sdwt)- average weight of 100 matured and uniformly dried seeds (g), shoot length (shl)-length of shoot from bottom to tip of the longest leaf (cm), root length (rl)-length of the root from base to root tip (cm), germination percent (ge)-percentage of seed germinated noted every 24 h after 72 h and upto 7 days, leaf length (ll)-length from the bottom to the tip of the flag leaf (cm), leaf width (lw)-width of the middle portion of the flag leaf (cm), plant height of 1 month old plants (ph)-length of the tallest tiller from soil surface to the tip of the panicle (cm), tiller number per plant (nt)-total number of tillers per plant, plant height of mature plants (pha)-length of the tallest tiller from soil surface the tip of the panicle (cm), number of panicles (np)-total number of panicles with seed set exceeding 15%, panicle length (pal)-length (cm) from neck to last spikelet of main panicle, 50% flowering (df)-number of days taken for 50% flowering in that family, seed weight per panicle (gr wt/pa)-filled seed weight (g) averaged over five randomly chosen panicles, yield per plant (yi/p) - mean of the weight of filled grains (g) from 5 plants, plot yield-average weight of filled grains from the plot of 102 m. The data for the above parameters was obtained by taking mean of two experimental observations in two seasons in the field. Following grain analysis traits were measured twice using Bio-vis image analysis software and the mean of 20 seeds was taken; seed area (ga), seed length (gl), seed weight (gw), seed perimeter (gp), seed density (gd), seed roundness (grd), seed compactness (gc), seed elongation (gel), seed roughness (grg), seed perimeter/length (gp/l) and seed perimeter/weight (gp/w). The 25 BC3F4 families were grown in petridish in the incubator, provided uniform conditions and germination percentage was recorded at every 24 h interval.
Trait correlation and statistical analysis: Correlation between 27 vegetative, mature plant morphological traits and seed traits pairs in BC3F4 population was calculated at two probability levels (p<0.05 and p<0.01) in Microsoft Excel using trait averages. The significant correlation was indicated at p = 0.05 and p = 0.01, respectively. Coefficient of variation of 7 mature plant morphological traits was computed by using formula:
RESULTS
Trait segregation and field performance: The values of seven yield related traits in BC3F4 and KMR3 showed large variation (Table 1). Phenotypic transgressive variation was observed for all the traits. The coefficient of variation for yi/p was highest (i.e., 25.63), while that for df was the lowest (i.e., 3.2). Yield per plant and plot yield for each IL was recorded in the field (Fig. 2). Yi/p was highest in IL50 (i.e., 89.5 g) where as lowest yi/p was recorded in IL491 (i.e., 36.9 g). Plot yield did not consistently reflect the yi/p. Plot yield was highest in IL50 (i.e., 1101 g). However, IL410 had lowest (i.e., 718 g) plot yield but it was among the top 5 yielding ILs according to yi/p.
Trait analysis and field performance: The frequency distribution of phenotypic characteristics for each trait in the best 25 ILs is shown in Fig. 3. Most of the traits showed transgressive segregation. However, some traits such as leaf length in seedling stage were positively skewed towards less length whereas, percentage germination and seed length/width ratio were negatively skewed towards higher ratio.
| Table 1: | Statistics of yield-related traits of KMR3 and the 25 ILs |
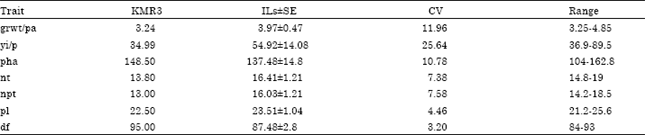 | |
| Seed weight per panicle (gr wt/pa)-filled grain weight (g) averaged over five randomly chosen panicles, Yield per plant (yi/p)-mean of the weight of filled grains in g from 5 plants, Plant height at maturity (pha)-length of the tallest tiller from soil surface to the tip of the panicle in cm, Tiller number per plant (nt)-total number of tillers per plant, Number of productive tillers (npt)-total number of panicles with seed set exceeding 15%, Panicle length (pl)-length in cm from neck to last spikelet of main panicle, 50% flowering (df)-number of days taken for 50% flowering in that family, SE standard error | |
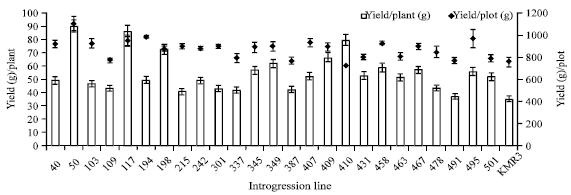 | |
| Fig. 2: | Yield of 25 introgression lines in the field study compared to KMR3. Yield per plant-average weight of filled grains (g) from 5 plants and plot yield-average weight of filled grains from 66 plants (33 plants in 2 rows). The data represented is the mean of two experimental observations in two seasons in the field. Yield/plant: hollow Square and yield/plot: filled square |
Nearly 80% of the ILs were shorter (plant height of 1 month old plants) than KMR3. Panicle length was more than KMR3 in 82% of the ILs. However, the highest yielding line IL 50 showed only 2.2% increase in panicle length (Fig. 5). Germination percent at 4 days was higher than KMR3 in 40% ILs and less in 30% of ILs. Twenty-four ILs outperformed KMR3 for seed weight per panicle. Fifteen lines showed at least 20% increase in seed weight per panicle with a maximum increase of 50% in IL 50. All the ILs outperformed KMR3 for yield per plant with a range of increase from 5 to 155%. The line IL 50 showed the highest increase in yield (Fig. 5). All the ILs outperformed KMR3 for number of productive tillers. The highest increase was in IL 117(i.e., 42.3%; Fig. 5) which was also the second highest yielding line. Days to 50% flowering was less in all the ILs compared to KMR3. These results indicate that for increasing yield in case of introgressions from O. rufipogon, following traits need improvement. Early flowering is a positive trait. Obviously, number of tillers, number of panicles and seed weight per panicle need to be higher than the restorer line KMR3. Panicle length or 100 seed weight need not be much higher than that of the restorer. Vegetative traits like leaf length and weight only indicate higher initial vigor of the plant but not necessarily higher yield. Long shoot and root in seedling stage is a positive trait and may confer vigor in the adult plant. These results confirm that genes introgressed from O. rufipogon into an elite background can improve key agronomic traits, even though O. rufipogon is phenotypically inferior to the cultivated variety.
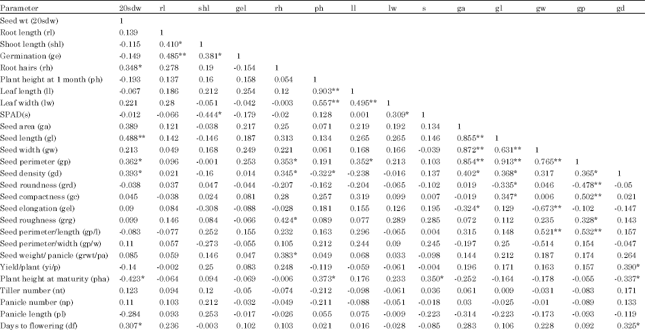
Phenotypic trait correlations: The correlation between traits was estimated by regressing phenotypic values of one trait on those of another trait (Table 2). Among all the traits, highest significant positive correlation was observed between nt and npt (0.98). Highest significant negative correlation was observed between grd and gc (0.95). Grd showed significant negative correlation with maximum number of other traits (gc; grg; gel; gp/l; pha; pl; gl and gp), where as gc showed maximum number of significant positive correlations (gel; grg; gp/l; gp/w; pha; pl; gl and gp). Most of the adult phenotypic traits showed minimum correlations (only 1 or 2) with any other mature plant trait. Yi/p showed significant positive correlation only with 2 parameters gr wt/pa and gd. Df showed significant positive correlation with sdwt and gd. Pha showed maximum correlations among all the traits.
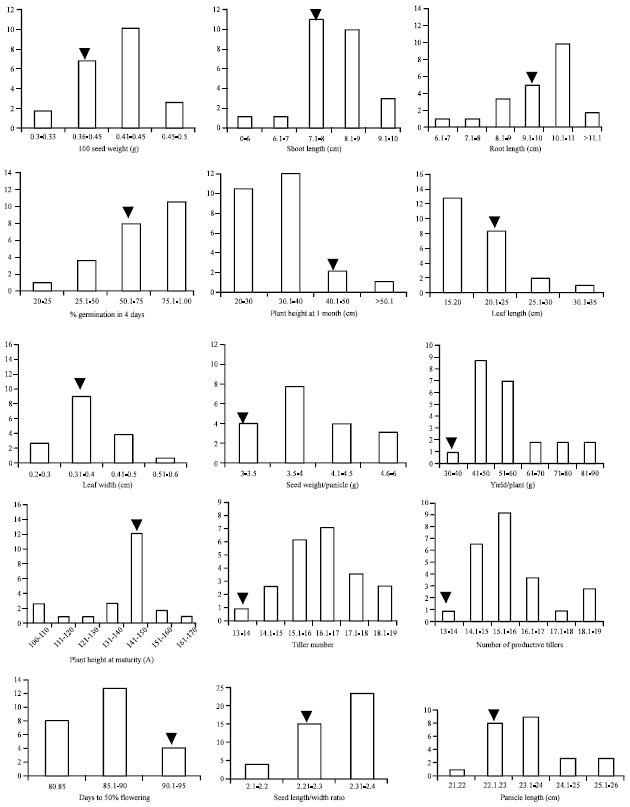 | |
| Fig. 3: | Frequency distribution of traits in the 25 high yielding introgression lines of KMR3. Arrowheads indicate KMR3. Y-axis: number of individuals |
Seed germination-extent and rate: Rate of germination was analyzed in 25 high yielding ILs till 7 days after sowing (Fig. 4). The control line KMR3 showed a sigmoid pattern of germination and only 90% seeds germinated even after 7 days. The lower yielding ILs such as 491 and 215 showed earlier germination than KMR3 (94 and 100% germination, respectively after 4 days).
| Table 2: | Significant correlation coefficients (r) among traits in KMR3 derived BC3F4 population. r = 0.32 at p = 0.05 (*); r = 0.45 at p = 0.01 (**) |
 | |
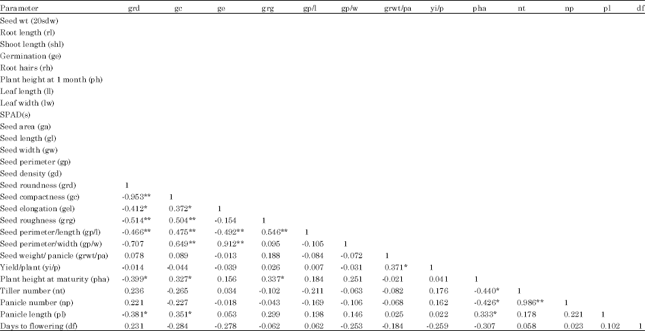 | |
| Plant phenotypic traits: Seed weight (sdwt)- average weight of 100 matured and uniformly dried seeds, Root length (rl)-length of the root from base to root tip in cm, Shoot length (shl)-length of shoot from bottom to tip of the longest leaf in cm, Germination percent (ge)-percent seed germinated noted every 24 h after 72 h and upto 7 days, Root hairs (rh) – No of root hairs at 7 days of germination, Plant height in 1 month old plants (ph) - length of the tallest tiller from soil surface the tip of the panicle in cm, Tiller number per plant (nt)-total number of tillers per plant, Leaf length (ll)-length from the bottom to the tip of the flag leaf in cm, Leaf width (lw) -width of the middle portion of the flag leaf in cm, SPAD(s)- SPAD chlorophyll meter value in 1 month old plants, Seed analysis traits: Seed area (ga), Seed length (gl), Seed width (gw), Seed perimeter (gp), Seed density (gd), Seed roundness (grd), Seed compactness (gc), Seed elongation (gel), Seed roughness (grg), Seed perimeter/l | |
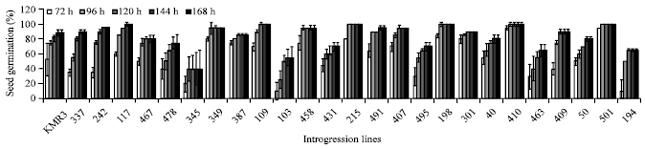 | |
| Fig. 4: | Germination time course of 25 high yielding introgression lines of KMR3. Seed germination from a total of 20 seeds was counted after 3 days and recorded after every 24 h till 7 days. Data represented here is mean of 3 individual experiments (n = 3) |
On the other hand IL 337 which was among the lower yielding lines of the 25 lines, showed sigmoid pattern of germination but with a steep slope (late germinating i.e., 90% germination after 6 days). It was significant that the highest yielding line IL 50 showed a straight-line slope for rate of germination and reached only 80% after 7 days. The high yielding line IL 117 showed similar pattern of germination like KMR3 although with higher rate. Another high yielding IL410 showed quick germination with 95% germination after 3 days and reaching to 100% after 4 days. The IL 345 having lowest rate of germination (i.e., 40% after 7 days) could also show considerable increase in yield (i.e., 61.7%) over KMR3. It appears that late and steady germination as in IL50 may be an adaptation to ensure highest survival and growth in unpredictable environmental conditions.
Analysis of mean percentage phenotypic variation of top ten high-yielding ILs: The top ten ILs having higher grain yield per plant were selected for the analysis of mean percentage phenotypic variation of some seedling and phenotypic traits (Fig. 5). When compared with KMR3, these high yielding ILs showed an apparent increase in yi/p by 5% in IL491 to 155% in IL50. However, 9 of the 10 ILs showed increase in plot yield compared to KMR3 which ranged from 0.5% in IL387 to 45.5% in IL50. IL410 showed 5% decrease in plot yield compared to KMR3, which had 125% increase in terms of yi/p. Sdwt of four ILs showed a decrease (i.e., IL50, IL410, IL198 and IL409) but the others had higher values than KMR3. Ge was 20% less than KMR3 in the highest yielding IL (IL50). Ph and pha, ll, SPAD value and df were less compared with KMR3 in almost all the ILs indicating that these traits are negative indicators of higher yield. All the ILs outperformed the recurrent parent KMR3 by 8-37% and 15-42% for nt and npt, respectively and 22% for grwt/pa with an average of 1-49%. Percentage increase in gd was higher in 5 of the top 10 Ils (i.e., IL50, IL117, IL410, IL198 and IL409), whereas it was negative or nearly zero in 5 of the 10 bottom ILs (i.e., IL301, IL387, IL337, IL215 and IL491). It is concluded that the number of tillers, number of productive tillers, seed weight per panicle and seed density are the major positive traits influencing the yield of the ILs positively.
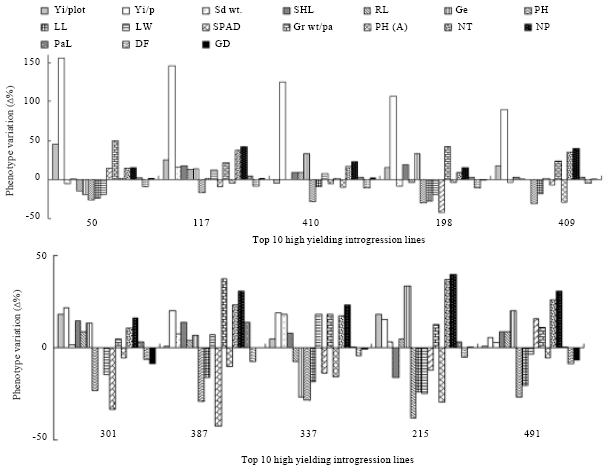 | |
| Fig. 5: | Analysis of top 10 high-yielding ILs. Mean percentage phenotypic variation (Δ%) over the recurrent parent KMR3 for Yield (g)/plot (Yi/plot); grain yield/plant (Yi/pl); 100 seed weight (100 Sd wt); shoot length in seedling stage (SHL); root length in seedling stage (RL); percentage of germination after 96 hours (Ge); plant height in seedling stage (PH); leaf length in seedling stage (LL); leaf width in seedling stage (LW); SPAD chlorophyll meter value (S); seed weight per panicle (Gr wt/pa); plant height in adult stage (PHA); number of tillers (NT); number of productive tillers (NP); panicle length (PaL); days of 50% flowering (DF); seed density (GD). The Δ% was calculated as 100(IL-mean of KMR3)/Mean of KMR3. Horizontal line at ordinate zero represents KMR3 value |
DISCUSSION
Several investigations have been made earlier to define a robust method of selection for yield which can withstand dynamic and unfavorable environment. To this end, ideotype approach (Peng et al., 2008), pedigree selection (Ali, 2011) and many other (Sofi et al., 2006) have been shown to be beneficial in various studies. Various selection strategies and genes have been shown to increase productivity by delaying heading date, increasing plant height and panicle size (Ali, 2011; Xue et al., 2008). However, high yield in the present KMR3 ILs was associated with dwarfness, low 100 seed weight and early flowering. The strategies of the KMR3 ILs to increase yield do not strictly support the earlier reports. In the present study, it is shown that each IL follows different paths to increasing yield. Recently, it was also shown that yield based selection is as effective as phenotypic trait based selection in the late generations of breeding (Yuan et al., 2011) supporting the results of this study from seed to seed.
The contributing factors for high yield are different in different ILs and were considered as follows. The highest yield of IL50 was associated with its slow but steady rate of germination, lower plant height, higher SPAD value, early flowering, higher seed weight per panicle and high seed density despite not much increase in number of panicles and panicle length compared to KMR3. The slow rate of germination indicated a communicative behavior of the sibling roots and could be an advantageous feature for seedling establishment in case of an unpredictable or unfavorable environmental delay in germination as reported in Arabidopsis (Biedrzycki et al., 2010). The link between germination rate and SPAD chlorophyll value in ILs is supported by the differential effect of the embryonic axis exertion on developing photosynthetic system and chlorophyll formation (Moore and Lovell, 1970). In another similar instance, impaired growth during germination due to a mutation in Starch Branching Enzyme (SBE) was observed in maize inbred lines (Xia et al., 2011).
The next highest yielding IL 117 had higher rate of seed germination, longer root and shoot length in seedling stage, highest increase in number of panicles and seed density, increase in panicle length, moderate increase in seed weight per panicle despite having less SPAD value and less plant height compared to KMR3. The ILs 410 and 409 which are among the top five yielders show nearly similar pattern of increase and decrease in traits with IL 337, one of the bottom 5 high yielders. However, IL 410 showed 125% yield increase and associated traits were high seed density, faster rate of germination, decrease in SPAD value, number of panicles and seed weight per panicle. The line IL 337 was among the bottom 5 lines probably due to the moderate rate of seed germination, least SPAD value and less seed density despite having more number of panicles and higher seed weight per panicle. An interesting and new observation is that higher seed germination rate was associated with low SPAD value and vice versa only in all these top 10 high yielding ILs. The above observation was supported by a negative correlation (i.e., -0.17) between germination % (ge) and SPAD meter value (s), however, this correlation was not significant when estimated by regressing phenotypic values of the BC3F4 IL population (Table 2).
Jennings (1964) proposed that selection for the stay-green trait combined with large, erect flag leaves is a valuable breeding objective for increased yield potential. Donald (1968) proposed the ideotype approach for plant breeding. He defined crop ideotype as an idealized plant type with a specific combination of characteristics favorable for photosynthesis, growth and grain production. Later Peng et al. (1994) proposed New Plant Type (NPT) and then a revised ideotype was proposed later (Peng et al., 2008) to define a plant type that was theoretically efficient. However, the results obtained in this study of the ILs do not support their ideotype concept. Yuan et al. (2011) suggested that yield-based selection was as effective in increasing rice grain yield as trait-based selection in the late generations of the breeding cycle supporting our results in the present study. As a follow up from the current work, two of the ILs were nominated to multi-location trials of the All India Coordinated Rice Improvement Programme (AICRIP) during Kharif, 2009. The results show that these lines were in the top 10 lines at 12 locations among 65 entries tested at 35 locations (DRR, 2010). These results from KMR3-O. rufipogon introgression lines are also supported by observations on Swarna-O. nivara introgression lines.
CONCLUSION
The current study indicated that while introgressing from O. rufipogon the following traits need to be improved for increasing yield: (1) tiller number and number of panicles per plant should be higher; (2) early flowering (3) steady rate of germination (4) high seed weight per panicle and (5) high seed density. Seedling phenotypes did not show strong correlation with increasing yield in this study. Yield/plant does not always reflect the plot yield. The novel association of seed germination rate with SPAD value in most of these high yielding ILs needs further study to dissect its link with high yield. Late and steady germination may be an adaptation to ensure highest survival and growth in unpredictable environmental condition as shown by highest yielding IL in this study. Thus it is clear that it is possible to have diametrically opposite strategies in each yield component to increase yield and restricting to a specific ideotype in selection approaches may be counterproductive in rice breeding.
ACKNOWLEDGMENT
This study was supported by the Department of Biotechnology, Govt of India (BT/AB/03/FG/DRR-II/01/2004-05). We thank G Ashok Reddy for technical assistance in field work and Dr BC Viraktamath, Project Director for encouragement and support.
REFERENCES
- Akinwale, M.G., G. Gregorio, F. Nwilene, B.O. Akinyele, S.A. Ogunbayo, A.C. Odiyi and A. Shittu, 2011. Comparative performance of lowland hybrids and inbred rice varieties in Nigeria. Int. J. Plant Breed. Genet., 5: 224-234.
CrossRefDirect Link - Ali, M.A., 2011. Pedigree selection for grain yield in spring wheat (Triticum aestivum L.) under drought stress conditions. Asian J. Crop Sci., 3: 158-168.
CrossRefDirect Link - Asif, M., M.Y. Mujahid, I. Ahmad, N.S. Kisana, M. Asim and S.Z. Mustafa, 2003. Determining the direct selection criteria for identification of high yielding lines in bread wheat (Triticum aestivum L.) bread wheat (Triticum aestivum L.). Pak. J. Biol. Sci., 6: 48-50.
CrossRefDirect Link - Biedrzycki, M.L., T.A. Jilany, S.A. Dudley and H.P. Bais, 2010. Root exudates mediate kin recognition in plants. Communicative Integrative Biol., 346: 28-35.
PubMed - Akinbile, C.O. and A.Y. Sangodoyin, 2011. Evapotranspiration, soil and water quality implications on upland rice production. Asian J. Crop Sci., 3: 169-178.
CrossRefDirect Link - Marri, P.R., N. Sarla, L.V. Reddyand and E.A. Siddiq, 2005. Identification and mapping of yield and yield related QTLs from an Indian accession of Oryza rufipogon. BMC Genet., 6: 33-33.
CrossRefDirect Link - McCouch, S.R., M. Sweeney, J. Li, H. Jiang and M. Thomson et al., 2007. Through the genetic bottleneck: O. rufipogon as a source of trait-enhancing alleles for O. sativa L. Euphytica, 154: 317-339.
CrossRef - Mobasser, H.R., D.B. Tari, M. Vojdani, R.S. Abadi and A. Eftekhari, 2007. Effect of seedling age and planting space on yield and yield components of rice (Neda Variety). Asian J. Plant Sci., 6: 438-440.
CrossRefDirect Link - Moncada, P., C.P. Martinez, J. Borrero, M. Chatel Jr. and H. Gauch et al., 2001. Quantitative trait loci for yield and yield components in an Oryza sativa × Oryza rufipogon BC2F2 population evaluated in an upland environment. Theor. Applied Genet., 102: 41-52.
CrossRef - Moore, K. and P. Lovell, 1970. Differential effects of the embryonic axis on chlorophyll production and photosynthesis of mustard cotyledons. Planta, 93: 289-294.
CrossRefDirect Link - Pimsaen, W., S. Jogloy, B. Suriharn, T. Kesmala, V. Pensuk and A. Patanothai, 2010. Genotype by environment (G×E) interactions for yield components of jerusalem artichoke (Helianthus tuberosus L.). Asian J. Plant Sci., 9: 11-19.
CrossRefDirect Link - Babu, A.P., M.S. Ramesha, T. Sudhakar, C.S. Reddy, B.P.M. Swamy, B.C. Viraktamath and N. Sarla, 2009. Marker assisted introgression of QTL yld2.1 or sub QTL regions from O. rufipogon increases yield of KMR3 and derived hybrids. Proceedings of the 7th International Symposium on Rice Functional Genomics, November 16-19, 2009, International Rice Research Institute, pp: 44.
- Sadeghzadeh, D. and K. Alizadeh, 2005. Relationship between grain yield and some agronomic characters in durum wheat under cold dryland conditions of Iran. Pak. J. Biol. Sci., 8: 959-962.
CrossRefDirect Link - Septiningsih, E.M., J. Prasetiyono, E. Lubis, T.H. Tai, T. Tjubaryat, S. Moeljopawiro and S.R. McCouch, 2003. Identification of quantitative trait loci for yield and yield components in an advanced backcross population derived from the Oryza sativa variety IR64 and the wild relative O. rufipogon. Theor. Applied Genet., 107: 1419-1432.
Direct Link - Sofi, P., A.G. Rather and M.Y. Zargar, 2006. Breaking yield barriers in rice: Remodeling photosynthesis. Am. J. Plant Physiol., 1: 113-126.
CrossRefDirect Link - Tanksley, S.D. and S.R. McCouch, 1997. Seed banks and molecular maps: Unlocking genetic potential from the wild. Science, 277: 1063-1066.
CrossRefPubMedDirect Link - Thomson, M.J., T.H. Tai, A.M. McClung, X.H. Lai and M.E. Hinga et al., 2003. Mapping quantitative trait loci for yield, yield components and morphological traits in an advanced backcross population between Oryza rufipogon and the Oryza sativa cultivar Jefferson. Theor. Applied Genet., 107: 479-493.
Direct Link - Tian, F., D.J. Li, Q. Fu, Z.F. Zhu, Y.C. Fu, X.K. Wang and C.Q. Sun, 2006. Construction of introgression lines carrying wild rice (Oryza rufipogon Griff.) segments in cultivated rice (O. sativa L.) background and characterization of introgressed segments associated with yield-related traits. Theor. Applied Genet., 112: 570-580.
Direct Link - Xia, H., M. Yandeau-Nelson, D.B. Thompson and M.J. Guiltinan, 2011. Deficiency of Maize Starch-Branching Enzyme I results in altered starch fine structure, decreased digestibility and reduced coleoptile growth during germination. BMC Plant Biology, 11: 95-95.
PubMed - Xie, X., F. Jin, M.H. Song, J.P. Suh and H.G. Hwang et al., 2008. Fine mapping of a yield-enhancing QTL cluster associated with transgressive variation in an Oryza sativa × O. rufipogon cross. Theor. Applied Genet., 116: 613-622.
PubMed - Xiong, L.Z., K.D. Liu, X.K. Dai, C.G. Xu and Q. Zhang, 1999. Identification of genetic factors controlling domestication‐related traits of rice using as F2 population of a cross between Oryza sativa L. and O. rufipogon. Theor. Applied Genet., 98: 243-251.
Direct Link - Xiao, J., J. Li, S. Grandillo, S.N. Ahn, L. Yuan, S.D. Tanksley and S.R. McCouch, 1998. Identification of trait-improving quantitative trait loci alleles from a wild rice relative, Oryza rufipogon. Genetics, 150: 899-909.
Direct Link - Xiao, J., S. Grandillo, S.N. Ahn, S.R. Mccouch, S.D. Tanksley, J. Li and L. Yuan, 1996. Genes from wild rice improve yield. Nature, 384: 223-224.
CrossRefDirect Link - Xue, W.Y., Y. Xing, X. Weng, Y. Zhao and W. Tang et al., 2008. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet., 40: 761-767.
CrossRefDirect Link - Yoon, D.B., K.H. Kang, H.J. Kim, H.G. Ju and S.J. Kwon et al., 2006. Mapping quantitative trait loci for yield components and morphological traits in an advanced backcross population between Oryza grandiglumis and the O. sativa japonica cultivar Hwaseongbyeo. Theor. Applied Genet., 112: 1052-1062.
PubMedDirect Link - Yuan, W., S. Peng, C. Cao, P. Virk and D. Xing et al., 2011. Agronomic performance of rice breeding lines selected based on plant traits or grain yield. Field Crops Res., 121: 168-174.
CrossRefDirect Link