
Valiollah Rameeh
Agriculture and Natural Resources Research Center of Mazandran, Sari, Iran
International Journal of Plant Breeding and Genetics
Year: 2011 | Volume: 5 | Issue: 4 | Page No.: 349-357







ABSTRACT
Linextester analysis of two testers of spring type and six lines of winter type rapeseed varieties were used to estimate combining ability and heterosis of phonological traits, plant height and grain yield. Significant mean squares of parents vs. crosses revealed significant heterosis for all the traits but linextester mean square which indicating the non additive genetic effects was only significant for plant height and grain yield. High narrow sense heritability estimates for phonological traits indicating the importance of additive genetic effects for these traits. Significant Specific Combining Ability (SCA) effects of plant height and grain yield were observed for only a few crosses. Most of the crosses had significant over winter and spring parents heterosis for all the traits, therefore heterosis of crosses is more reliable than SCA effects in winterxspring crosses of rapeseed. All of the crosses had significant negative over winter parent heterosis effects for days to flowering, days to end of flowering and plant. Significant positive over spring parent heterosis effect was displayed for days to maturity and plant height in most of the crosses, indicating, in compare to their spring parents most of F1 genotypes were tall and late maturity. Significant positive over winter and spring parents heterosis effects was observed for grain yield in some of crosses which had high performances of these traits.
PDF Abstract XML References Citation
Received: February 28, 2011;
Accepted: May 16, 2011;
Published: July 09, 2011
How to cite this article
Valiollah Rameeh, 2011. Heterosis and Combining Ability Assessment for Phenological Traits, Plant Height and Grain Yield in WinterxSpring Combinations of Rapeseed Varieties. International Journal of Plant Breeding and Genetics, 5: 349-357.
DOI: 10.3923/ijpbg.2011.349.357
URL: https://scialert.net/abstract/?doi=ijpbg.2011.349.357
DOI: 10.3923/ijpbg.2011.349.357
URL: https://scialert.net/abstract/?doi=ijpbg.2011.349.357
INTRODUCTION
The oilseed Brassicas are the world’s third most important source of oils and their production has witnessed a steady upward movement by the aim of conventional and modern plant breeding approaches (Sabaghnia et al., 2010). In rapeseed breeding for hybrid and open pollinated varieties, general and specific combining ability effects (GCA and SCA) are important indicators of the potential of inbred lines in hybrid combinations. The relative importance of additive (GCA) and non-additive (SCA) gene actions within a breeding population is important to determine which breeding procedure will efficiently improve the performance of the characters of interest (Singh and Chaudhury, 1977). Exploitation of genetic variability in any crop species is considered to be critical for making further genetic improvement in grain yield as well as other economically important traits (Mahmood et al., 2003; Inamullah et al., 2006; Rehman et al., 2009). The limited geographic range of Brassica napus combined with intensive quality breeding has led to narrow genetic basis in this species (Seyis et al., 2005). Rapeseed cultivars are classified as winter or spring types according to their vernalization requirement in order to induce flowering. Due to restricted genetic basis of spring type of rapeseed varieties, winter type of rapeseed varieties can be merit candidate for increasing yield potential and genetic variation of spring type in winter x spring types or semi-winter type combinations (Qian et al., 2007). Butruille et al. (1999) investigated the possibility to increase yield of spring material by using winter germplasm. In other reports only spring material with spring material (Brandle and McVetty, 1990; Engquist and Becker, 1991; Rameeh, 2010) or winter material with winter material was crossed (Amiri-Oghana et al., 2009). The linextester analysis is one of the efficient methods of evaluating large number of inbreds as well as providing information on the relative importance of GCA and SCA effects for interpreting the genetic basis of important plant traits (Singh and Chaudhury, 1977). By using this method, phonological traits, grain yield and other yield associated traits were studied for combining ability, heterosis heritability (Shen et al., 2002; Wang et al., 2007). In earlier studies on spring cultivars of oilseed rape (Huang et al., 2010; Rameeh, 2011) were noted the important role of GCA and SCA effects for days to flowering but due to high heritability estimates for this trait in these reports, the prime importance of GCA effects were emphasized. Likewise, studies with winter cultivars of this species (Amiri-Oghana et al., 2009; Sabaghnia et al., 2010; Rameeh, 2011) showed both additive and dominance gene effects to have a significant role in the inheritance of flowering time. Significant negative GCA and SCA effects were reported for days to flowering and plant height. Significant GCA and SCA effects were reported for grain yield in Ethiopian mustard (Teklewold and Becker, 2005) and repeseed (Rameeh, 2010).
Heterosis and heterobeltiosis have extensively been explored and utilized for boosting various quantity and quality traits in different crops (Khan et al., 2004; Parvez Sofi et al., 2007; Bagheri et al., 2008; Chapi et al., 2008; Alghamdi, 2009; Mahmoud and Ahmed, 2010; Immanuel Selvaraj et al., 2011). With sufficient level of heterosis, commercial production of hybrid varieties would be justified (Nassimi et al., 2006; Radoev et al., 2008) The availability of genetically distant plant material is of great importance for successful hybrid breeding programs because breeders want to exploit the expected heterosis effects. This effect is higher in cases where the parents are relatively different on genetic base. The extent of heterosis in rapeseed has been analyzed in a number of studies with widely varying results, depending on the materials used. For grain yield in spring rapeseed hybrids, an average high parent heterosis of 30% with a range of 20-50% was observed, while for winter rapeseed hybrids an average high parent heterosis of 50% was reported, ranging from 20 to 80% as reviewed by McVetty (1995). Flowering is the most critical stage influencing the yield of oilseed rape (Faraji et al., 2008). The unset of flower initiation can have strong influence on flower, pod and seed number (Diepenbrock, 2000; Downey and Rimmer, 1993; Yasari and Patwardhan, 2006). Habekotte (1997) used a sensitivity analysis within a crop growth model to study options for increasing grain yield in winter oilseed rape. The most promising crop type for high grain yield combined late maturity with early flowering (Downey and Rimmer, 1993). Heterosis is commercially exploited in rapeseed (Brassica napus L.) and its potential use has been demonstrated in turnip rape (B. rapa L.) and Indian mustard (B. juncea L.) for most agronomic traits (Teklewold and Becker, 2005; Zhang and Zhu, 2006). Early flowering in brassica can provide adequate time for grain formation process and can certainly cause early maturity and higher yields; therefore negative heterosis is desirable for flowering. Significant negative mid-parent and better-parent heterosis were reported for days to 50% flowering and physiological maturity (Nassimi et al., 2006). In this study mid parent heterosis for days to 50% flowering and physiological maturity ranged from -0.04 to -2.78 and -0.01 to -3.06, respectively and also its high parent heterosis for these traits ranged from -0.92 to -2.78 and -0.01 to -4.08, respectively. Significant correlation was reported between post-anthesis duration with some of important agronomic traits including number of pods per plant, 1000 seed weight and oil yield (Jeromela et al., 2007).
Although linextester analysis are frequently used in oil seed rape breeding to assess genetic parameters for yield associated traits but in a few studies were focused on heterosis over winter or spring parents. The objectives of this study were therefore (i) to identify general and specific combining abilities and narrow-sense heritability for phonological traits, plant height and grain yield in adapted rapeseed cultivars and (ii) comparison of heterosis over winter and spring parents for studied traits.
MATERIALS AND METHODS
Six winter rapeseed (B. napus L.) cultivars including Zarfam, Licord, Talayeh, Okapi, Modena and Opera as Lines were crossed with two spring testers including H1 and H2 based on line x tester crossing scheme during 2006-07. Twelve F1s along with their parents were grown in a randomized complete block design with three replications at Biekol Agriculture Research Station, located in Neka, Iran (13°, 53' E longitude and 43° 36' N latitude, 15 m above sea level) during winter 2007-08. Each plot was consisted of four rows 5 m long and 40 cm apart. The distance between plants on each row was 5 cm resulting in approximately 300 plants per plot which were sufficient for F1 genetic analysis in each experiment. The soil was classified as a deep loam soil (Typic Xerofluents, USDA classification) contained an average of 280 g clay kg-1, 560 g silt kg-1, 160 g sand kg-1 22.4 g organic matter kg-1 with a pH of 7.3. Soil samples were found to have 45 kg ha-1 (mineral N in the upper 30-cm profile. Fertilizers were applied at the rates of 100: 50: 90 kg ha-1 of N: P:K, respectively. All the plant protection measures were adopted to make the crop free from insects. Grain yield (adjusted to kg ha-1) was recorded based on three middle rows of each plot. The data were recorded on ten randomly selected plants of each entry of each replication for days to flowering, days to end of flowering, duration of flowering and days to maturity and plant height. Grain yield (adjusted to kg ha-1) was recorded based on two middle rows of each plot.
Data for the genotypes were subjected to line x tester analysis (Singh and Chaudhury, 1977) to estimate General Combining Ability (GCA) and Specific Combining Ability(SCA). A t-test was used to test whether the GCA and SCA effects were different from 0. For each hybrid and each studied trait, the difference between hybrid and the mean of each spring and winter type parent was computed separately. A Least Significant Difference (LSD) was used to test whether these differences were different from 0.
RESULTS
Linextester analysis of variance: There were significant differences among crosses for all the traits except duration of flowering which implies the existence of genetic variability among crosses (Table 1). Variances among spring genotypes, used as testers, were greater than winter types genotypes, indicating some degrees of paternal effects on the studied traits. Variance of lines x tester was significant for plant height and grain yield, indicating the importance of non additive genetic effects for these traits. High narrow-sense heritability estimates for days to flowering, days to end of flowering, duration of flowering and days to maturity indicating the prime importance of additive genetic effects for these traits.
General combining ability effects: Significant negative GCA effect of days to flowering was detected for Zarfam and also it was negative for H1 and Okapi. Significant negative GCA effect of days to end of flowering was estimated for H1 and Zarfam and also it was negative for Opera and Licord (Table 2). Non of lines and testers had significant GCA effects for duration of flowering but out of 8 parents (lines and testers), 4 parents including H2 as tester and also licord, Modena and Opera as lines had non significant negative GCA effects for this trait.
| Table 1: | Analysis of variance for phenological traits, plant height and grain yield of rapeseed (Brassica napus L.) based on line x tester method |
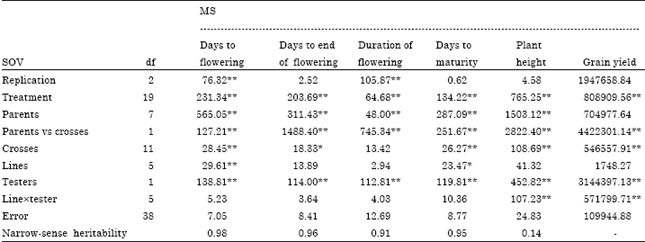 | |
| *, ** Significant at p<0.05 and 0.01, respectively | |
| Table 2: | Estimates of GCA effects for phenological traits, plant height and seed yield of rapeseed (B. napus L.) |
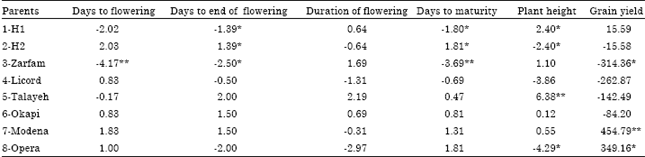 | |
| *, ** Significant at p<0.05 and 0.01, respectively | |
Significant negative GCA effects were displayed for H1 and Zarfam and these genotypes had also significant negative GCA effects for days to end of flowering. Significant negative GCA effect of plant height was detected for H2 and Opera and also H1 and Talayeh had significant positive GCA effects for this trait. The lines including Modena and Opera had significant positive GCA effects for grain yield.
Specific combining ability effects: The result of SCA effects of crosses for studied traits is presented in Table 3. Non of crosses had significant SCA effect of phonological traits. Out of 12 crosses, 6 crosses had non significant negative SCA effects of days to flowering. For days to end of flowering the crosses TalayehxH1 and LicordxH2 had high negative SCA effects of days to end of flowering. Non of combinations had significant positive SCA effect for duration of flowering and LicordxH1 and OperaxH1 had high positive SCA effect for this trait. The crosses including TalayehxH1 and TalayehxH2 had significant negative and positive SCA effects of plant height, respectively. The combinations ZarfamxH1 and LicordxH2 had significant positive SCA effects of grain yield.
Heterosis: Significant negative over winter parent heterosis of days to flowering and days to end of flowering were observed for all the combinations (Table 4). The crosses including OperaxH1, LicordxH1, TalayehxH1 and ModenaxH1 had high significant negative over winter parent heterosis of days to flowering and they were considered as good combinations for reducing this trait.
| Table 3: | Estimates of SCA effects for phenological traits, plant height and grain yield in crosses of two testers and six lines of rapeseed |
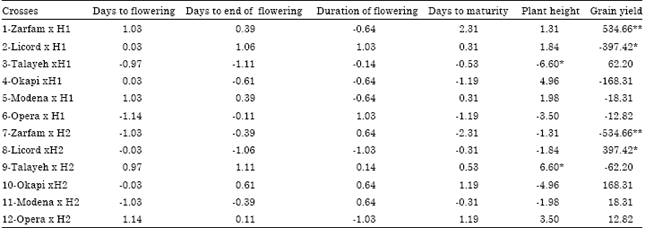 | |
| *, ** Significant at p<0.05 and 0.01, respectively | |
| Table 4: | Heterosis over winter parent for phenological traits, plant height and grain yield in crosses of two testers and six lines of rapeseed |
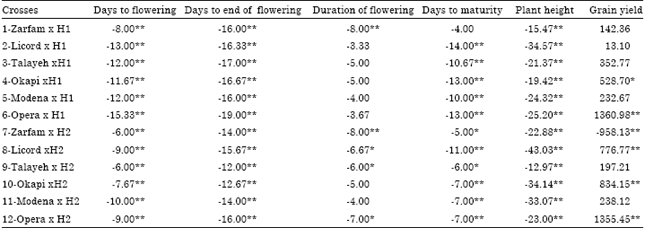 | |
| *, ** Significant at p<0.05 and 0.01, respectively | |
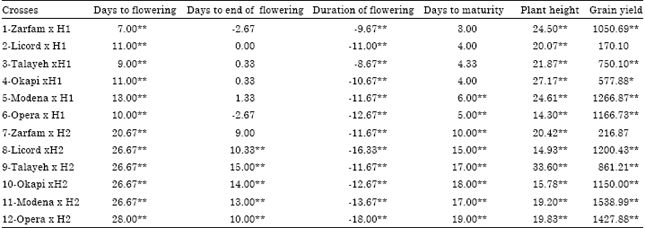
| Table 5: | Heterosis over spring parent for phenological traits, plant height and grain yield in crosses of two testers and six lines of rapeseed |
 | |
| *, ** Significant at p<0.05 and 0.01, respectively | |
The crosses including OperaxH1 and TalayehxH1 had high significant negative over winter parent heterosis of days to end of flowering (Table 4). Out of 12 crosses, 5 crosses had significant negative over winter parent heterosis for duration of flowering. All of the crosses had significant negative heterosis of plant height over winter parents. All of combinations had significant positive over spring parents heterosis for days to flowering and also out of 12 crosses, 5 crosses had significant positive over spring parents heterosis for days to end of flowering (Table 5). All of the combinations had significant negative and positive over spring parents heterosis for duration of flowering and plant height, respectively. Eight crosses had significant positive over spring parents heterosis for days to maturity. The crosses including OkapixH1, OperaxH1, LicordxH2, OkapixH2 and OperaxH2 had significant positive over winter and spring parents heterosis for grain yield and most of these crosses had high grain yield performance.
DISCUSSION
Significant differences was observed among the crosses for all the traits except duration of flowering. Genetic variability among crosses was important for the studied traits except duration of flowering. The testers were more varied than the lines for all the traits. Due to adaptability of winter type parents of rapeseed to cold regions and the place of the experiment under study was classified as moderate region, the variations of winter parents for grain yield were low. Although significant parent vs crosses mean squares for all the traits indicating heterosis and the importance of non additive genetic effects for the traits but high narrow sense heritability estimates for phenological traits indicating the prime importance role of additive genetic effects for these traits. Similarly, various studies on spring (Huang et al., 2010; Rameeh, 2011) and winter (Amiri-Oghana et al., 2009; Sabaghnia et al., 2010) cultivars of oilseed rape have shown the important role of additive genetic effects for phenological traits.
Significant negative GCA effect of days to flowering for the line in including Zarfam and its negative effects for tester such as H1, inicating these line and tester have additive genetic effects for decreasing this trait. In earlier studies (Huang et al., 2010; Sabaghnia et al., 2010; Rameeh, 2011) were reported significant negative GCA effects for days to flowering. All genotypes with significant negative GCA effect for end of flowering had significant negative or negative GCA effect for days to flowering. The variation of GCA effects for duration of flowering was less than to other phenological traits and non of lines and testers had significant GCA effects for this trait. Due to early maturity is the most important breeding subject in north province in Iran to have enough opportunity for cultivation of second crop, so the genotypes including H1 and Zarfam with significant negative GCA effects for days to end flowering are more desirable. Decreasing of plant height is one the most ideo- type characteristics in rapeseed breeding (Downey and Rimmer, 1993), therefore the genotypes including H2, Licord and Opera with significant negative GCA effect for plant height were considered as good combiners for this trait. In previous study (Teklewold and Becker, 2005) was reported significant negative GCA effect for plant height in Brassica juncea. The lines including Modena and Opera with significant positive GCA effects for grain yield were good combiners for this trait. This finding is in agreement with Rameeh (2010) and Amiri-Oghana et al. (2009) reports for significant positive GCA effects of parents for grain yield in spring and winter rapeseed, respectively.
Due to high narrow sense heritability estimates and prime importance of additive genetic effects of phenological traits, non significant SCA effects of crosses were detected for these traits. This finding is in contrast to earlier studies on spring cultivars of oilseed rape have shown the important role of SCA effects for phenological traits (Nassimi et al., 2006) and also winter cultivars of this species (Sabaghnia et al., 2010). The crosses including TalayehxH1, OperaxH1 ZarfamxH2 and Modena x H2 with significant negative SCA effects for days to flowering and days to maturity are favored. TalayehxH1 with significant negative SCA effect for plant height is suitable crosses for decreasing this trait. The crosses including ZarfamxH1 and LicordxH2 with significant positive SCA effects for grain yield were detected as good combinations for increasing this trait. Similarly, in earlier studies were reported significant SCA effect of crosses for grain yield in on various studies on spring (Huang et al., 2010; Rameeh, 2011) and winter (Amiri-Oghana et al., 2009; Sabaghnia et al., 2010) cultivars of oilseed rape have shown the important role of additive genetic effects for phenological traits.
Most of winterxspring combinations had significant of heterosis effects for all the traits in compare to their SCA effects. Significant negative over winter parent heterosis of days to flowering and days to maturity were observed for all the combinations indicating early flowering and early maturity were controlled by dominant genes, therefore in crosses of winterxspring types of rapeseed varieties it is possible to improve early maturity spring lines. This findings is in agreement with Nassimi et al. (2006) who reported significant negative mid-parent and better-parent heterosis for days to 50% flowering and physiological maturity and also in their study mid parent heterosis for days to 50% flowering and physiological maturity ranged from -0.04 to -2.78 and -0.01 to -3.06, respectively. Spring type of rapeseed genotypes are more early maturity than winter type rapeseed varieties, so significant positive over spring parents heterosis were observed for days to flowering and days to maturity. Significant negative heterosis effect of duration of flowering was observed only for spring parents, indicating the moderate or warm regions like the place of this study spring type of rapeseed had more duration of flowering than winter type. The plant height of F1 progenies of winter x spring type varieties were more taller and shorter than spring type and winter type parents, respectively, so significant negative heterosis of plant height was observed only for spring type parent. Out of 12 crosses, 5 and 10 crosses had significant positive heterosis effects of grain yield over than spring and winter types parents, respectively. The crosses including OkapixH1, OperaxH1, LicordxH2, Okapi xH2 and OperaxH2 with significant positive over winter and spring parents heterosis for grain yield were considered as good combinations and most of these crosses had high grain yield performance. Similarly, for grain yield in spring rapeseed hybrids, an average high parent heterosis of 30% with a range of 20-50% was observed, while for winter rapeseed hybrids an average high parent heterosis of 50% was reported, ranging from 20 to 80% as reviewed by McVetty (1995).
In conclusion all of phenological traits were more heritable than the other studied traits, so the efficiency of selection for these traits will be high. Due to more variation of heterosis than SCA effects of crosses for all the traits, therefore selection of merit combinations based on heterosis is more easier than SCA effect. All of the crosses had significant negative heterosis effects for days to flowering, days to maturity and plant height over than spring type parents, so F1 progenies of winterxspring types rapeseed varieties were early flowering, early maturity and shorter than spring type parent.
ACKNOWLEDGMENTS
The author wish to thanks from Agricultural and Natural Resources Research Center of Mazandaran and Seed and Plant Institute Improvement (SPII) for providing genetic materials and facility for conducting the experiment.
REFERENCES
- Alghamdi, S.S., 2009. Heterosis and combining ability in a diallel cross of eight faba bean (Vicia faba L.) genotypes. Asian J. Crop Sci., 1: 66-76.
CrossRefDirect Link - Amiri-Oghana, H., M.H. Fotokianb, F. Javidfar and B. Alizadeh, 2009. Genetic analysis of grain yield, days to flowering and maturity in oilseed rape (Brassica napus L.) using diallel crosses. Int. J. Plant Prod., 2: 19-26.
Direct Link - Bagheri, N., N.B. Jelodar and A. Ghanbari, 2008. Diallel analysis study of yield and yield-related traits in rice genotypes. Int. J. Agric. Res., 3: 386-396.
CrossRefDirect Link - Brandle, J.E. and P.B.E. McVetty, 1990. Geographical diversity parental selection and heterosis in oilseed rape. Can. J. Plant Sci., 70: 935-940.
CrossRef - Chapi, O.G., A.S. Hashemi, E. Yasari and G.A. Nematzadeh, 2008. Diallel analysis of seedling traits in canola. Int. J. Plant Breed. Genet., 2: 28-34.
CrossRefDirect Link - Diepenbrock, W., 2000. Yield analysis of winter oilseed rape (Brassica napus L.): A review. Field Crops Res., 67: 35-49.
CrossRefDirect Link - Downey, R.K. and S.R. Rimmer, 1993. Agronomic improvement in oilseed brassicas. Adv. Agric., 50: 1-66.
CrossRef - Faraji, A., N. Latifi, A. Soltani and A.H.S. Rad, 2008. Effect of high temperature and supplemental irrigation in flower and pod formation in two canola (Brassica napus L.) cultivars at mediterranean climate. Asian J. Plant Sci., 7: 343-351.
CrossRefDirect Link - Habekotte, B., 1997. Evaluation of seed yield determining factors of winter oilseed rape (Brassica napus L.) by means of crop growth modeling. Field Crops Res., 54: 137-151.
CrossRef - Huang, Z., P. Laosuwan, T. Machikowa and Z. Chen, 2010. Combining ability for seed yield and other characters in rapeseed. Suranaree J. Sci. Technol., 17: 39-47.
Direct Link - Selvaraj, C.I., P. Nagarajan, K. Thiyagarajan, M. Bharathi and R. Rabindran, 2011. Studies on heterosis and combining ability of well known blast resistant rice genotypes with high yielding varieties of rice (Oryza sativa L.). Int. J. Plant Breed. Genet., 5: 111-129.
CrossRefDirect Link - Inamullah, H.A., M. Fida, G.H. Siraj-ud-din, and G. Rahmani, 2006. Evaluation of the heterotic and heterobeltiotic potential of wheat genotypes for improved yield. Pak. J. Bot., 38: 1159-1167.
Direct Link - Jeromela, M.A., R. Marinkovic, A. Mijic, M. Jankulovska and Z. Zdunic, 2007. Interrelationship between oil yield and other quantitative traits in rapeseed (Brassica napus L.). J. Cent. Eur. Agric., 8: 165-170.
Direct Link - Khan, M.S., I.H. Khalil and M.S. Swati, 2004. Heterosis for yield components in sunflower (Helianthus annuus L.). Asian J. Plant Sci., 3: 207-210.
CrossRefDirect Link - Mahmood, T., M. Ali, S. Iqbal and M. Anwar, 2003. Genetic variability and heritability estimates in summer mustard (Brassica juncea). Asian J. Plant Sci., 2: 77-79.
CrossRefDirect Link - Mahmoud, A.M. and T.A. Ahmed, 2010. Magnitude of combining ability and heterosis as influenced by type of soil in grain sorghum (Sorghum bicolor L. Moench). Asian J. Crop Sci., 2: 1-11.
CrossRefDirect Link - Nassimi, A.W., Raziuddin, S. Ali and N. Ali, 2006. Study on heterosis in agronomic characters of rapeseed (Brassica napus L.) using diallel. J. Agron., 5: 505-508.
CrossRefDirect Link - Sofi, P.A., A.G. Rather and Z. Dar, 2007. Association of heterotic expression for grain yield and its component traits in maize (Zea mays L.). Int. J. Agric. Res., 2: 500-503.
CrossRefDirect Link - Qian, W., O. Sass, J. Meng, M. Li, M. Frauen and C. Jung, 2007. Heterotic patterns in rapeseed (Brassica napus L.): I. Crosses between spring and Chinese semi-winter lines. Theoret. Appl. Genet., 115: 27-34.
CrossRef - Radoev, M., H.C. Becker and W. Ecke, 2008. Genetic analysis of heterosis for yield and yield components in rapeseed (Brassica napus L.) by quantitative trait locus mapping. GenetIcs, 179: 1547-1558.
Direct Link - Rameeh, V., 2010. Combining ability and factor analysis in F2 diallel crosses of rapeseed varieties. Plant Breed. Seed Sci., 62: 73-83.
CrossRef - Rameeh, V., 2011. Heritability and other genetic parameters assessment for flowering associated stress indices in oil seed rape varieties. Int. J. Plant Breed. Genet., 5: 268-276.
CrossRefDirect Link - Rehman, A.U., M.A. Ali, B.M. Atta, M. Saleem, A. Abbas and A.R. Mallahi, 2009. Genetic studies of yield related traits in mungbean (Vigna radiata L. Wilczek). Aust. J. Crop Sci., 3: 352-360.
Direct Link - Sabaghnia, N., H. Dehghani, B. Alizadeh and M. Mohghaddam, 2010. Diallel analysis of oil content and some agronomic traits in rapeseed (Brassica napus L.) based on the additive-dominance genetic model. Aust. J. Crop Sci., 4: 609-616.
Direct Link - Seyis, F., W. Friedt and W. Luhs, 2005. Development of resynthesized rapeseed (Brassica napus L.) forms with low erucic acid content through in ovulum culture. Asian J. Plant Sci., 4: 6-10.
CrossRefDirect Link - Teklewold, A. and H.C. Becker, 2005. Heterosis and combining ability in a diallel cross of Ethiopian mustard inbred lines. Crop Sci., 45: 2629-2635.
Direct Link - Yasari, E. and A.M. Patwardhan, 2006. Physiological analysis of the growth and development of canola (Brassica napus L.) under different chemical fertilizers application. Asian J. Plant Sci., 5: 745-752.
CrossRefDirect Link