
K. D. Adjata
Universite de Lome, Ecole Superieure d Agronomie, Lab. De Virologie et de Biotechnologies Vegetales (LVBV), B.P.1515 Lome, Togo
E. Muller
CIRAD-UMR BGPI (Biologie et Genetique des Interactions Plantes-Parasites) TA 41 / K, Campus International de Baillarguet, 34398 Montpellier cedex 5, France
M. Peterschmitt
CIRAD-UMR BGPI (Biologie et Genetique des Interactions Plantes-Parasites) TA 41 / K, Campus International de Baillarguet, 34398 Montpellier cedex 5, France
M. Aziadekey
Universite de Lome, Ecole Superieure d Agronomie, Lab. De Virologie et de Biotechnologies Vegetales (LVBV), B.P.1515 Lome, Togo
O. Traore
Institut de l Environnement et de Recherche Agricole (INERA), 01 B.P. 476 Ouagadougou 01, Burkina Faso
M. Abala
Universite de Lome, Ecole Superieure dAgronomie, Lab. De Virologie et de Biotechnologies Vegetales (LVBV), B.P.1515 Lome, Togo
N. Tchala
Universite de Lome, Ecole Superieure dAgronomie, Lab. De Virologie et de Biotechnologies Vegetales (LVBV), B.P.1515 Lome, Togo
K. Alomassou
Universite de Lome, Ecole Superieure dAgronomie, Lab. De Virologie et de Biotechnologies Vegetales (LVBV), B.P.1515 Lome, Togo
Y. M.D. Gumedzoe
Universite de Lome, Ecole Superieure dAgronomie, Lab. De Virologie et de Biotechnologies Vegetales (LVBV), B.P.1515 Lome, Togo
International Journal of Plant Breeding and Genetics
Year: 2011 | Volume: 5 | Issue: 2 | Page No.: 150-158







ABSTRACT
Four new clones (KH01, KH02, KH03 and KH04) obtained from genetic crossing between M. glaziovii (a wild variety) and Kanigbeli (a local cassava cultivar cultivated in Togo were planted in the field and were characterized morphologically on the basis of their behaviour vis-à-vis of Begomovirus infections and their agronomic performances. The results from the behavioural evaluation of the four selected clones (KH01, KH02, KH03 and KH04) vis-à-vis of Begomovirus infections and their agronomic performances, after three generations of genetic crossing, reveal that the selected clones present the best viral tolerance (mark 2). And concerning their agronomic performances, the selected clones have a good harvest index and a good effectiveness of production.
PDF Abstract XML References Citation
Received: September 03, 2010;
Accepted: December 08, 2010;
Published: March 25, 2011
How to cite this article
K. D. Adjata, E. Muller, M. Peterschmitt, M. Aziadekey, O. Traore, M. Abala, N. Tchala, K. Alomassou and Y. M.D. Gumedzoe, 2011. Strategies of Effective Control of Cassava Mosaic Disease by Genetic Selection in Togo. International Journal of Plant Breeding and Genetics, 5: 150-158.
DOI: 10.3923/ijpbg.2011.150.158
URL: https://scialert.net/abstract/?doi=ijpbg.2011.150.158
DOI: 10.3923/ijpbg.2011.150.158
URL: https://scialert.net/abstract/?doi=ijpbg.2011.150.158
INTRODUCTION
Cassava (Manihot esculenta Crantz) is a major factor in food security across Africa. However, the crop is susceptible to losses due to biotic stresses, in particular to viruses of the genus Begomovirus (family Geminiviridae) that cause Cassava Mosaic disease (CMD) which is the major problem of cassava production in most countries in sub-Saharan regions (FAO, 2003). This Cassava Mosaic Disease (CMD) can result in massive economic impact and can destabilized food security in Africa, as evidenced during the severe CMD in 1990s (Legg and Thresh, 2003), with losses of up to 20 to 95% in the case of the virus of Cassava Mosaic Disease (CMD) (Fargette et al., 1988). CMD is caused by either ACMV alone or by virulent recombinants of different cassava begomoviruses and synergism among these begomoviruses (Legg and Fauquet, 2004; Hong et al., 1993; Saunders et al., 2002). The important problem among the viral agents of the disease (Begomovirus) is their permanent recombination (Padidam et al., 1999) which allow them to overcome the resistance of new created clones. Another important thing about these recombinant begomoviruses is their virulence which has an important impact on the severity of CMD and the fall of productivity. So, there is a need of creating variability among the existing cultivated cassava population in Togo through genetic improving. Hahn et al. (1980) produced a hybrid between cassava and M. glaziovii proved to be much more productive than the existing clones used in Nigeria and in other countries in the sub- region. But, the problem encountered with these new cultivars, is that once introduced, in any case in Togo, they often lose their resistance. Nassar (1992) manipulated the meiotic restitution occurring in interspecific hybrids of M. pseudoglaziovii with cassava to produce triploid types that showed very good productivity under semi-arid conditions. If genes which control productivity in cassava are polygenes with additive model action, as it is the case for many crops, certain trissomics of this crop may be more productive than their diploid ancestors. In general, the production of polyploidy via sexual means is advantageous both from genetic and evolution viewpoints, as it offers a vigorous heterotic effect and releases useful genetic variability adaptations. In the area of resistance to plant viruses, the research team has been trying its efforts to build a fundamental knowledge base to respond to real agronomic problems (Lecoq et al., 2004). Since it was noticed that, all the improved CMD-resistant cultivars which have been developed at the IITA and introduced in Togo, have never lasted in the conditions of the country, an action towards the selection of new cultivars in the environmental conditions of Togo is important.
The aim of this research was, according to the new development of CMD in Togo, to create variability among the population of cassava varieties cultivated in Togo through the selection of new clones resistant/tolerant to begomoviruses by using genetic crossing between M. glaziovii (a wild variety) and Kanigbeli, a local cassava cultivar cultivated in Togo and focusing on their agronomic performances.
MATERIALS AND METHODS
Origin of planting material: The genotypes were selected based on Plant Vigor (PV), Flowering Ability (FA), Sprouting Ability (SA) and Resistance/Tolerance (R/T) to CMD. All possible crosses involving 3 parents with various degrees of resistance to CMD (resistant, moderately resistant and susceptible varieties) were made by using the complete mating scheme at the High School of Agriculture’ research station in Lome (Togo) during 2003/2004 and 2004/2005 growing seasons and the seeds were made available by LVBV (Laboratory of Plant Virology and Biotechnology) for the study.
Planting materials: Seeds from F3 generation, obtained by crossing a local cultivar Kanigbeli with Manihot glaziovii (KH), were planted in pots in nursery prior to transplanting in the field, and watered twice a day over three weeks and then once a day until the seedlings were established. Established seedlings were transplanted in the field to produce woody cuttings for the study. Mature stakes (25 cm long) of the parents were planted at the beginning of the rainy season (June 2006) in order to have enough planting material. Observations of the clones were based essentially on Plant Vigor (PV), Flowering Ability (FA), Sprouting Ability (SA) and Resistance/Tolerance (R/T) to CMD.
Field experiments: Mature stems of six clones including four ((KH01, KH02, KH03 and KH04) clones bred against CMD at SEA of ESA (UL) and two cultivars (Gbazekoute (GBAZE), an improved cultivar) and Fetonegbodji (FETO, a local susceptible cultivar)) used as references, were planted in randomized complete blocks design with two replicates. Each plot consisted of a minimum of 20 plants planted according to the cultural scheme 1x1 m, giving a plant population of 10,000 plants per hectare. The blocks are 1.5 m from each other and in each block, the plots are 1.5 m from each other. No fertilizer or herbicide was applied during the course of the experiment and hand weeding was done when necessary.
Parameter evaluation: The following parameters have been evaluated during the experimentation:
| • | Identification of infected plants at the sprouting |
| • | Evaluation of disease progression within the plots twice a month |
| • | Evaluation of disease symptom severity on each plant twice a month |
Study of clone behaviour vis-à-vis of CMD: To study the behaviour of each clone to the CMD, all the plants of each clone were first marked and then inspected twice a month in order to study the visual progression and the pressure of the disease within the plots. This observation was done during the whole vegetative phase. The study of the pressure of the disease on the clones consists in giving marks ranging from 1 to 5 (mark 1 = no visible symptom; 2 = 25% of the leaves of the plant are infected; 3 = 50% of the leaves of the plant are infected; 4 = 75% of the leaves of the plant are infected and 5 = 75 to 100% of the leaves of the plant are infected). The study of disease progression within the plots consists in marking twice a month, new infected plants.
Study of the performance of each clone vis-à-vis of CMD: To study the performance of each clone vis-à-vis of CMD, the plants were harvested according to their level of the severity of symptom score. So, for each level of symptom severity considered, 5 plants were harvested. All these considerations should allow bringing out the performance of each clone. For this last aspect, the harvest index was the yield characteristic considered. In fact, the harvest index (H.I.) which is calculated by dividing the weight of tuberous roots by the weight of the whole plant, gives an idea on the percentage on the useful part of the plant harvested.
Data analysis: The harvest index was calculated according to Hunt (1982). Data collected were subjected to Analysis of Variance (ANOVA) using generalized Linear Model (GLM) procedures (SAS Institute Inc., 2000) and mean differences determined by LSD at 5% level of significance. The progression of CMD within the plots was analysed by using the mathematical model of disease progress of Berger (1977) and of Van der Plank (1963).
RESULTS
Disease evolution within the plots: Results obtained from the disease progression study showed that, throughout the trial, the susceptible local reference FETO was completely infected 100% (Table 1).
The study of CMD symptom score on the clones: From the analysis of the visual measurement of the progression of the disease within the plots, the level of symptom severity of CMD on each cultivar was established. The level of symptom severity of the disease on the clones depends on the susceptibility of each clone (Table 2). This level of symptom severity is respectively 2.5-3.5; 1-1.2; 1-1.2; 1-1.3; 1.2-1 and 1.4-1.1 for Feto, Gbaze, KH01, KH02, KH03 and KH04 (Table 2).
| Table 1: | Time evolution of the incidence (%) of CMD within various clones |
 | |
| LSD (5%) = 46.82 | |
| Table 2: | Level of CMD symptom score on the clones in the time |
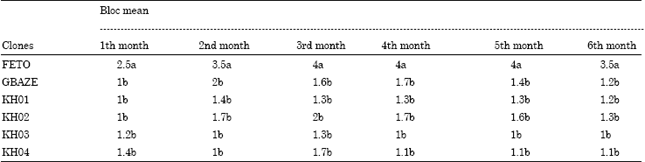 | |
| LSD (p<0.05) = 1.839. Values with different letters are not significantly different | |
Evaluation of agronomic aptitudes of the clones: Development and growth of the various selected clones. The size of the various clones gradually and regularly increased, as the Table 3 indicates below. A statistical analysis of the variance at 5% indicates that the variation observed in the height of the clones could be an effect due to the nature of each clone. However, the separation of the averages does not show a difference statistically significant (p< 0.05) only from the fourth month of vegetation.
The mean yield of tuberous roots of each clone in contrast with the yield of reference cultivars: Results obtained from the production of each clone according to the yield characteristics among which, the harvest index which is calculated by dividing the weight of tuberous roots by the weight of the whole plant (Table 4), reveal that the impact of CMD on the mean yield of infected plants depended on the level of symptom severity, on the cultivars and on the susceptibility to CMD. It is remarkable to notice that apart from the susceptible clone (FETO), which recorded the highest disease score, all the clones overcome the disease.
Concerning data obtained for the average length of the stalk (Table 4), all the clones are ranged between 3 and 4 cm. This character is very important for breeders. By order of magnitude, this character is as follows: GBAZE>KH02>KH04>KH01>KH03>FETO (Table 5). There is no significant difference between the intra- and inter-clones. Results from the analysis of the number of root tuber by clone also revealed that there is no significant difference between the clones. Nevertheless, the highest average obtained was 11 tubers.
| Table 3: | Evolution of the average height (mean ± SD) of the various clones |
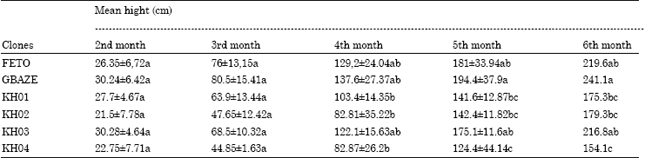 | |
| LSD (p< 0.05) = 44,69. Values with different letters are not significantly different | |
| Table 4: | Some characteristics of output in fresh tubers (mean±SD) |
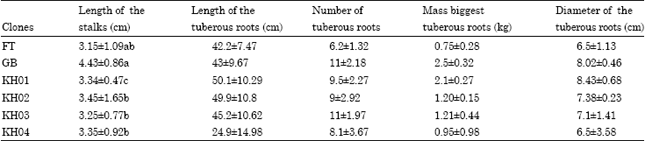 | |
| LSD (5%) = 1.920.Values with different letters are not significantly different | |
| Table 5: | Output in fresh root tubers of the clones, 9 months after plantation of the cuttings (mean±SD) |
 | |
| Table 6: | Relation between dry matter (DM) tuberous root (X) and that of the whole plant (Y) and the harvest effectiveness of the various clones |
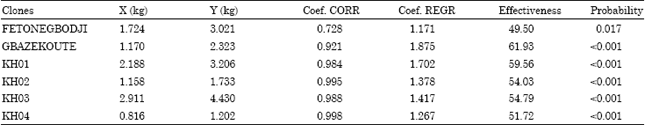 | |
| X: Mean productivity of dry matter (DM), Y: Quantity of DM by clone, Coef, CORR: Coefficient of correlation; Coef, REGR: Coefficient of regression | |
Harvest effectiveness determination: The output in root tubers is a very significant parameter of comparison of cassava clones. Unfortunately, this character (quantitative character) is very influenced by the environment. The harvest effectiveness appears to be a good criterion of selection, because little influenced by environmental conditions (Table 6).
Dry matter rate determination: The variance analysis of the dry matter rate was not significant (p<0.05), as well for the block as for the type of clone. That means that all the studied clones would present almost identical outputs in various processed products, under the same condition.
DISCUSSION
Incidence and propagation of the CMD within the selected clones: The visual observations made during this trial, revealed that the effect of CMD on the different clones relied on the initial condition of the planting material used. All the planting material used except the planting material from the local reference, are tolerant to CMD. Indeed, during the vegetation cycle, clones KH01, KH02, KH03 and KH04 presented a progressive and regular tendency to the evolution of CMD with a reduction of the number of infected individuals in time. This is a proof that these clones have high levels of tolerance to the CMD and would confirm the observations of Akano et al. (1997) who showed that improved clones having accumulated genes of resistance to the virus diseases acquire levels of resistance, proportional to the number of accumulated genes. In this study, symptoms totally disappear from the fourth month after plantation. Mean yield analysis in regard with the level of symptom severity on the infected plants reveal that the level of symptom could have an important impact on the yield of the clones. Thus, the local reference which was severely attacked, demonstrated the least productivity (3.13±0.71). The analysis of the losses according to the level of symptom severity revealed that the losses rely on the clone’s susceptibility (Table 4). This observation reported already in 1951 by Cours supposes that at the level of the considered clones, the first cause of the appearance of the symptoms (activity of the viruses in the plant) would be destroyed. It is not a question here of the disappearance of the viruses but of their activity like several authors showed it such as (Cours, 1951; Njock et al., 1996). Moreover, for the considered clones, the observation showed that the leaves presenting the symptoms were for the majority, located at the base of the plant. And thus after total fall of these leaves and this plant become without apparent symptoms but are not healthy. It was the particular case of the selected clones (KH01, KH02, KH03 and KH04) whose incidence rate passes from 15% in the first month to 0% in the second month after the installation.
Symptoms severity on the attacked clones: Apart from the improved clone GBAZE and the selected clones (KH01, KH02, KH03 and KH04), it has been observed that the local reference FETO (susceptible clone), expressed an increase in the symptoms severity between the third and fifth month. An analysis of the variance (5%) showed that there was significant difference between this clone and the others. Results on the incidence of the disease within the clones show that during the first three months the clone FETO has presented more infected plants than all the other clones. Observations carried out between the first and the second month revealed that there may be a time necessary for the immune system of the plant to stop the progression of the virus. Thus, the inhibition phenomenon of virus progression related to the abscission of the leaves carrying the symptoms, would explain the regression of the symptoms severity after three months of vegetations. Because almost observations showed that clones with a high level of resistance: KH01, KH02, KH03 and KH04 respectively, leaves carrying symptoms are located at the base of the plant. Resistance to CMD, acquired from Manihot glaziovii is a polygenic resistance and implies recessive genes for additive purposes (Mba and Dixon, 1997; Mahungu et al., 1994; Jennings, 1994). On this basis, the selected clones KH01, KH02, KH03 and KH04 having presented the same behavior of resistance could be classified among the resistant clones. The pressure or the severity of symptom score was also important on the plots with the susceptible cultivar (Fetonegbodji); this level of symptom severity is respectively 2-4, 1-1.2, 1-1.1, 1-1.5, 1.4-1 and 1.4-1 for FETO, GBAZE, KH01, KH02, KH03 and KH04. If the level of symptom severity is also remarkable on the susceptible cultivar (Fetonegbodji), it could be said that other biotic factors for example whiteflies (Bemisia tabaci) would play an important role; they are not the transmitters of viruses to the plants, but by their feeding on the plants could also play a secondary influence on the reaction of these plants facing the CMD.
Agronomic aptitudes: growth in height of the various clones: The homogeneity of growth observed in the first months could be explained by the fact that at these moments, the growth of each clone relies mainly on the reserves contained in the cuttings. The difference comes only in time according to the quantity of the accumulated dry matter. These observations support those of Indira et al. (2000) and Singh and Chaudhary (2006) which showed that the size of cassava clones (improved as local) does not present a significant difference during the first months in growth. The highly significant difference observed between the clones, as from the fourth month, for the height indicates a variability of the importance of the vegetative parts of the plant and consequently of the photosynthetic effectiveness. In conclusion, the character height of the clones is largely influenced by the conditions of the environment and the type of stem of the clone which is a character of genetic determinism.
Height analysis reveal that clone FETO (local reference), with an average height of 219.6 cm, after six months, is significantly different from clones KH01, KH02 and KH04 (p<0.05). On the other hand there is no significant difference between FETO and clone KH03. Clones KH01, KH02, KH03 and KH04 are significantly different from GBAZE (improved reference).
Output in fresh tuber: The characteristics observed of the output explicitly the length of the tubers, their size (as indicate above) specify that the cycle of production is not yet in its term. The outputs obtained are, respectively 31.3, 83.5, 73.7, 79.5, 82.6 and 74.7 t ha-1 for clones FETO, GBAZE, KH01, KH02, KH03 and KH04 (Table 5). The selected clones are significantly different from the local reference. The outputs obtained indicate that the level of tolerance to Cassava Mosaic Disease (CMD) affects the output in tubers of the studied clones. These hybrids having presented very interesting outputs under the conditions which prevailed during our experimentation, notwithstanding the infection with the virus diseases, have a very high level of resistance to the CMD.
The hybrid clones KH are more performing than the local reference FETO (susceptible cultivar) for output. This typical example illustrates the observations of Thottappilly et al. (2003). The behavior of the hybrid clones KH as well illustrates the situation of tolerance of a given cultivar vis-à-vis of the infection of a pathogenic agent, in our case, the CMD. These results reveal that a use of the selected hybrid clones would ensure better outputs than those known today with the use of the local reference FETO and the improved reference GBAZE. This observation has also been done by Panahyan-e-Kivi et al. (2009).
Harvest index and effectiveness: The values of the harvest indices of all the clones ranged between 0.5 and 0.68. These values indicate that all the clones mobilize a significant share (more than half) of their dry matter synthesized in the development of tuberous roots. That confirms their great agronomic aptitudes for the production. This accumulation of the dry matter in the roots is more significant for the clone KH01 (harvest index = 0.68). It is followed by clones KH03 (0.66), KH04 (0.64) and KH02 (0.58). The clone FETO has accumulated more the dry matter in its vegetative part than in the roots. Indeed, its harvest index, equal to 0.5, means that it has accumulated in the roots only half of the dry matter mobilized for its vegetative growth (Islam et al., 2002). The increase in the harvest indices was accompanied for the clone KH01 by a weak development of the vegetative. Indeed these authors found that harvest indices of more than 0.60 were accompanied, for certain varieties, of a weak development of the vegetative axes. That would compromise the reproduction of new farming cycles, criterion of great importance for the farmers.
The harvest effectiveness specifies more the distribution of the dry matter in the plant. Thus more the effectiveness is large, less the clone is early and thus less it can be adapted to the environmental conditions. Among the hybrids, the clone KH04 would be the earliest followed by clone KH02 and then clone KH03 and finally the clone KH01.
CONCLUSION
One of the objectives of this study is to create variability among cassava population in Togo by genetic improving. Results obtained from cassava clones after three generations of genetic crossing revealed that not only these receive the expected genes, meaning that they received the polygenic resistance, but also the productivity performance. At this stage of our study we can say that it is possible to develop a stable CMD resistance in cassava in Togo.
ACKNOWLEDGMENT
We are grateful to the Direction de la Recherche of the University of Lome for sponsoring this project. This work is published with the permission of Pr. Y.M.D. GUMEDZOE, Director of the Direction de la Recherche of the University of Lome, Togo.
REFERENCES
- Berger, R.D., 1977. Application of epidemiological principles to achieve plant disease control. Ann. Rev. Phytopathol., 15: 165-181.
CrossRefDirect Link - Cours, G., 1951. Le manioc a Madagascar. Memoires de l'Institut Scientifique de Madagascar. Serie B Biologie Vegetale, 2: 203-400.
Direct Link - Fargette, D., C. Fauquet and J.C. Thouvenel, 1988. Yield losses induced by African cassava mosaic virus in relation to the mode and the date of infection. Trop. Pest Manage., 34: 89-91.
CrossRefDirect Link - Hahn, S.K, E.R. Terry and K. Leuschner, 1980. Breeding cassava for resistance to cassava mosaic disease. Euphytica, 29: 673-683.
CrossRefDirect Link - Hong, Y.G., D.J. Robinson and B.D. Harrison, 1993. Nucleotide sequence evidence for the ccurrence of three distinct whitefly transmitted geminiviruses in cassava. J. Gen. Virol., 74: 2437-2443.
PubMed - Legg, J.P. and C.M. Fauquet, 2004. Cassava mosaic geminiviruses in Africa. Plant Mol. Biol., 56: 585-599.
PubMed - Mahungu, N.M., A.G.O. Dixon and Kumbira, 1994. Breeding cassava for multiple pest resistance in Africa. Afr. Crop Sci. J., 2: 539-552.
Direct Link - Padidam, M., S. Sawyer and C.M. Fauquet, 1999. Possible emergence of new geminiviruses by frequent recombination. Virology, 265: 218-225.
CrossRefPubMedDirect Link - Panahyan-e-Kivi, M., A. Ebadi, A. Tobeh and Sh. Jamaati-e-Somarin, 2009. Evaluation of yield and yield components of lentil genotypes under drought stress. Res. J. Environ. Sci., 3: 456-460.
CrossRefDirect Link - Islam, M.R., M. Mahbubul Islam, M. Nuruzzaman, S. Akhtar and A.F. Mollah et al., 2002. Study of harvest index and genetic variability in white jute (Corchorus capsularis) germplasm. J. Biological Sci., 2: 358-360.
CrossRefDirect Link - Saunders, K., N. Salim, V.R. Mali, V.G. Malathi, R. Briddon, P.G. Markham and J. Stanley, 2002. Characterisation of Sri Lankan cassava mosaic virus and Indian cassava mosaic virus: Evidence for acquisition of a DNA B component by a monopartite Begomovirus. Virology, 293: 63-74.
PubMed - Singh, G.P. and H.B. Chaudhary, 2006. Selection parameters and yield enhancement of wheat (Triticum aestivum L.) under different moisture stress conditions. Asian J. Plant Sci., 5: 894-898.
CrossRefDirect Link - Van der Plank, J.E., 1963. Plant Disease: Epidemics and Control. Academic Press, Cambridge, Massachusetts, ISBN: 978-0-12-711450-7, Pages: 349.
CrossRefDirect Link - Lecoq, H., B. Moury, C.J.M. Desbiez, A. Palloix and M. Pitrat, 2004. Durable virus resistance in plants through conventional approaches: A challenge. Virus Res., 100: 31-39.
CrossRef