
C. Immanuel Selvaraj
Centre for Plant Molecular Biology, Tamil Nadu Agricultural University, Coimbatore, India
P. Nagarajan
Centre for Plant Molecular Biology, Tamil Nadu Agricultural University, Coimbatore, India
K. Thiyagarajan
Centre for Plant Breeding and Genetics, Tamil Nadu Agricultural University, Coimbatore, India
M. Bharathi
Centre for Plant Protections Studies, Tamil Nadu Agricultural University, Coimbatore, India
R. Rabindran
Centre for Plant Protections Studies, Tamil Nadu Agricultural University, Coimbatore, India
International Journal of Plant Breeding and Genetics
Year: 2011 | Volume: 5 | Issue: 2 | Page No.: 111-129







ABSTRACT
Twenty six rice genotypes were selected based on the rice blast disease reaction and yield attributes. Blast disease screening was done for all the entries under natural and artificial conditions to know their disease reaction. Line x tester analysis was carried out from the 20 selected entries with 4 lines having higher to moderate yield and 16 testers which showed blast disease resistance. General combining ability of parents and specific combining ability and heterosis among 64 hybrids for yield and its components were analysed. Under natural conditions and artificial screening conditions, among the 26 genotypes screened, highly significant lower mean disease reaction score and mean Potential Disease Incidence [PDI%] was recorded by a West African land race, Moroberekan. Mean squares due to females were larger in magnitude than male parents for all the characters. The magnitude of SCA variance was higher than GCA variance for all the characters except grain yield where the GCA variance was higher. Among the lines, IR 50 was found to be a good general combiner for six traits. Among the testers, IR 64 recorded high per sec performance along with high gca effects for panicle length, filled grains per panicle, 1000 grain weight and grain yield per plant. Most of the hybrids recorded positive significant standard heterosis values for grain yield per plant. IR 50/IR 64 recorded highest heterosis values for relative heterosis and TN 1/IR 64 recorded high heterobeltiosis and standard heterosis over the standard check ASD 16. Among 26 selected F1s, Most of the combinations were resistant.
PDF Abstract XML References Citation
Received: October 18, 2010;
Accepted: December 18, 2010;
Published: March 25, 2011
How to cite this article
C. Immanuel Selvaraj, P. Nagarajan, K. Thiyagarajan, M. Bharathi and R. Rabindran, 2011. Studies on Heterosis and Combining Ability of Well Known Blast Resistant Rice Genotypes with High Yielding Varieties of Rice (Oryza sativa L.). International Journal of Plant Breeding and Genetics, 5: 111-129.
DOI: 10.3923/ijpbg.2011.111.129
URL: https://scialert.net/abstract/?doi=ijpbg.2011.111.129
DOI: 10.3923/ijpbg.2011.111.129
URL: https://scialert.net/abstract/?doi=ijpbg.2011.111.129
INTRODUCTION
Rice is the world’s most important cereal crop. The recorded rice consumption in 2005 in India was around 85 million tonnes. Ten years down the line, it is expected to soar up to 92 million tonnes demanding more production from the paddy fields which is most unlikely to happen given the scenario of dwindling paddy fields (Subbiah, 2006). In Tamil Nadu, during the year 2007-2008, rice is being grown in an area of 17.89 lakh hectares with an production of 50 Lakh tonnes (Statistical Hand Book, 2010).
The estimated doubling of the population by 2050 will require a similar increase in food production. This has to be achieved by the development of high yielding rice varieties with improved nutritional quality and tolerance to biotic and abiotic stresses. Asia’s Green Revolution achieved with increase in crop productivity that were sufficient to lower the proportion of population suffering from chronic hunger from 40 to 20% while the overall population growth is more than doubled. In addition, by increasing yields on land already in production, hundreds of millions of hectares of tropical forests and other natural environments were saved from conversion to agriculture (Toenniessen et al., 2003).
Unfortunately, these expectations are short lived because the large areas of high yielding but genetically identical cultivars proved to be susceptible to pest and diseases. Among the biotic stresses diseases continue to be the major threat for increased production. Hence, the most urgent need is to increase the yield of rice by managing the problems caused by biotic and abiotic stresses (Hittalmani et al., 1995).
Biotic and abiotic stresses cause severe yield losses which can be managed to certain extent by evolving genotypes employing resistance breeding strategies. To counter such yield losses is identification of resistant varieties available in nature and introgression of major resistance genes in high yielding varieties to increase productivity and crop diversification, while developing a more sustainable agriculture (Hittalmani et al., 2000). The other way is by elucidating the basis of plant resistance through a comprehensive analysis of the molecular events that occur during pathogen-host recognition and the subsequent defense responses (Er-ming et al., 2005). Among the available genetic resources to increase rice productivity, hybrid rice has faired well and secured a good track record in uplifting the curse of ‘yield barrier’. Rice hybrids have an yield advantage of about 15 to 20% or more over the best conventionally bred varieties (Virmani, 1996).
The rice blast disease caused by Magnaporthe grisea (Hebert) Barr. (Asexual form known as Pyricularia grisea (Cooke) Sacc.), is one of the most serious fungal diseases which are widespread threatening the world rice production. Genetic resistance to rice blast has been and continues to be extensively used by rice breeders and pathologists to combat this disease. Numerous races of the fungus are prevalent (Ou, 1985). Blast resistance genes, commonly called Pi genes, providing a broad spectrum of resistance against the most prevalent races can be extremely valuable in rice breeding efforts (Fjellstrom et al., 2003). In many cultivars, blast-resistance is quite short-lived in field conditions as the pathogen mutates very often favored by the environment to spread the disease. Hence, breeding for more durably resistant cultivars has become a priority in rice improvement programmes throughout the world.
Plant breeding strategies leading to selection of hybrids needs expected level of heterosis as well as the specific combining ability. In breeding high yielding varieties of crop plant for qualitative and quantitative traits, plant breeders often face with the problem of selecting parents and crosses. Combining ability analysis is one of the valuable tools available to ascertain the combining ability effects and helps in selecting the desirable parents and crosses for the exploitation of heterosis. Line x tester analysis provides information related to general combining ability (gca) and specific combining ability (sca) effects of parents and it is helpful in estimating various types of gene actions (Rashid et al., 2007). Ganapathy et al. (2007) assessed the combining ability four best cross combination by line x tester analysis in rice including Moroberekan/MDU 5 and Moroberekan/CO 47 as best crosses in terms of higher yield and drought resistant. Line x tester analysis was done similarly to identify the was done in rice (Devi et al., 2006; Venkatesan et al., 2008; Kumar et al., 2008).
The main objective of this breeding program is to select the high yielding hybrids with leaf blast resistance under field conditions. The choice of the parents for the breeding program was based on the earlier reports for the testers. The lines were selected based on three important factors. They are as follows: The extensive usage of these rice varieties for cooking purpose in Southern parts of India (TamilNadu, Andhra Pradesh, Karnataka) for its fine grain quality, (e.g., White ponni, BPT 5204 and IR 50), Grain yield per hectare, consistency of leaf blast disease reaction (TN 1).
The testers namely ARBN 97, ARBN 138, ARBN 139, ARBN 142, ARBN 144 and ARBN 153 harboring major blast resistance genes Pi-5(t), Pi-9(t), Pi-12(t), Pi-1(t) + Pi-2(t), Pi-1(t) + Pi-4(t) and Pi-5(t) respectively, were obtained from Central Rice Research Institute, CRRI, Cuttack in the year 2004 were used directly in the crossing program to study the yield and the reaction of high yielding combination for leaf blast disease reaction. The genotypes or the accessions CB98002, CB98004, CB98006 and CB98013 obtained from Paddy Breeding Station, Coimbatore, were selected based on the records of the earlier evaluation trials, with moderate to higher yield combined with moderate to resistant leaf blast disease reaction.
Inheritance of resistance in the varieties Te-tep and Tadukkan were found to be trigenic (Padmanabhan et al., 1973; Padmavathi et al., 2005). Te-tep was used as a resistance source in breeding program (Ahn and Ou, 1982; Dillon et al., 2006). Rice cultivar with durable resistance has been reported in many countries. Moroberekan (Japonica), in West Africa. (Nottegham, 1993; Wang et al., 1994; Chen et al., 1999; Jeon et al., 2003; Wu et al., 2004). IR 64 is one of the most cultivated varieties in Asia and is highly resistant to blast disease (Berruyer et al., 2003). IR 64 harboured more than four blast resistance genes. (Sallaud et al., 2003; Lawrence et al., 2000). ARBN 153 (Pai-Kan-Tao) is a Japonica type NIL consisted of major resistance gene for leaf blast as reported by Ahn et al. (1996), Chen et al. (1999), Inukai et al. (1994), Mackill and Bonman (1992). CO 39 was highly susceptible for blast disease (Chen et al., 1999; Padmavathi et al., 2005; Hittalmani et al., 1995; Jeon et al., 2003). IR 50 was used as a susceptible check (Calvero, 1992; Jyothi et al., 2001).
In the light of the above facts and considering the potentials of resistance breeding, the present study was undertaken with the following objectives: 1) selection of highly reputed blast resistant genotypes with resistance genes and blast susceptible varieties having moderate to higher yield and to screen the rice genotypes for leaf blast disease reaction at two environments, 2) effecting hybridization among the chosen blast resistant and susceptible genotypes, study their heterosis pattern, combining ability, screening the F1 hybrids under artificial condition for rice blast resistance.
MATERIALS AND METHODS
Twenty six rice genotypes representing different geographical origin were obtained from Paddy Breeding Station, Coimbatore and Central Rice Research Institute (CRRI), Cuttack in the year 2004. (ARBN, Asian Rice Biotechnological Network) lines were introgressed with leaf blast disease resistance genes (Table 1).
Field screening for leaf blast disease reaction: All the rice genotypes were screened at Hybrid Rice Evaluation Centre, Gudalur, Tamilnadu, India (hot spot for leaf blast), where disease occurrence is throughout the year and maximum during winter season. Each entry was sown in a single row and replicated thrice with every adjacent row planted with Bharti, (a highly susceptible local cultivar for leaf blast). The entire nursery was surrounded on all sides by two rows of Bharti, as a spreader source for the pathogen. The observation of disease reaction was recorded, when the susceptible check was severely infected by leaf blast.
| Table 1: | Details of rice genotypes |
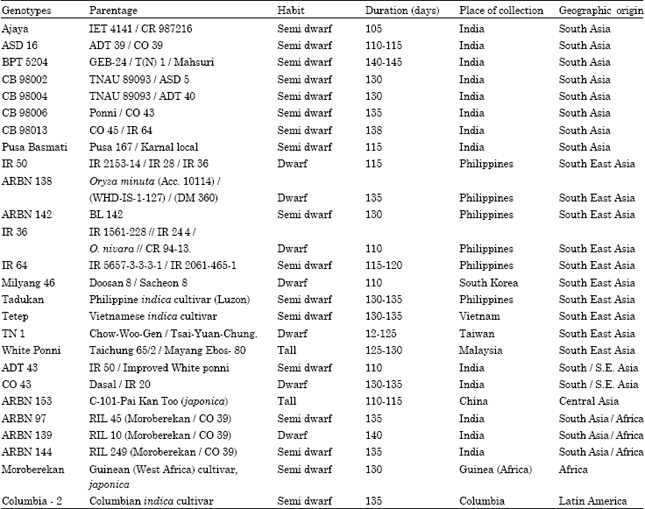 | |
Individual plant in each entry was scored based on the leaf blast severity following Standard Evaluation System (SES, IRRI, 2002) on a 0-9 scale as detailed at 35th day after sowing, when the susceptible check (Bharti) was fully infected. The Potential Disease Incidence (PDI%) was worked out using the formula given by McKinney (1923):
Artificial screening for leaf blast disease reaction: Artificial screening for rice blast disease was done in the specially constructed screen house with good irrigation facilities fitted with mist blowers, which can spray water in a fine mist inside the chamber. Subsequently, the seedlings were misted 4-5 times at intervals. The screen house was maintained at 32-37°C (day temperature) and 94 to 96% Relative Humidity (RH) for the potential disease occurrence. The rate of sporulation increases with increase in relative humidity provided with lower night temperature with minimum of 25°C. Inoculations with M. grisea Hebert (Barr) were performed 3 weeks after sowing either by spraying with conidial suspensions. For the spray method, 30 mL of a 50,000 conidia suspension with 0.5% gelatin were sprayed on each tray. The observation on the disease incidence was recorded, when the susceptible check was severely infected by blast. Observations were recorded from 20 plants in each entry following Standard Evaluation System (SES, IRRI, 2002) on 0-9 scale at 25th day after sowing. The resistant check used was IR 64. Observations were recorded in plants, when they were at third leaf stage. The Grade and criterion based on standard evaluation system is as follows, score 0-No lesions observed; score 1- Small brown specks of pin point size or larger brown specks without sporulating centre; score 3- Small roundish to slightly elongated necrotic grey sporulating spots about 1-2 millimeters in diameter with a distinct brown margin; score 5- Narrow or slight elliptical lesions, 1-2 mm in breadth, more than 3 mm long with brown margin; score 7-Broad spindle shaped lesion with yellow, brown or purple margin; score 9-Rapidly coalescing small, whitish, greyish or bluish lesions without distinct margins.
Selection, hybridization and biometric observations of F1s and parents: Twenty genotypes with 16 rice blast resistant lines viz., (ARBN 97, ARBN 138, ARBN 139, ARBN 142, ARBN 144, ARBN 153, IR 64, CB 98002, CB 98004, CB98013, CB 98006, Columbia 2, Milyang-46, Moroberekkan, Tadukkan, Te-tep) and four high yielding blast susceptible testers (IR 50, White ponni, TN 1, BPT5204) were selected based on the blast disease reaction after the screening procedures. The genotypes were raised in nursery beds with three staggered sowings at 10-15 days interval to ensure synchronized flowering to enable hybridization. Hybridization was carried out by wet cloth method or blowing method (Chaisang et al., 1967) and clipping method (Jennings et al., 1979). Crosses were effected between four female lines and sixteen male parents in a Line x Tester mating design and a total of 64 cross combinations were obtained. The 64 hybrids and 20% were raised along with the standard check ASD-16 in a Randomized Block Design which was replicated five times by adopting a spacing of 20x20 cm between rows and plants in a single row of each 1.5 m length consisting of 10 plants per row Single seedling per hill was planted. The recommended packages of practices were followed.
The following biometrical observations were recorded for the randomly selected hybrids and parents. Days to 50% flowering (DFL), Plant Height (PH), number of tillers per plant (NOTP), number of productive tillers per plant (NOPTP), Panicle Length (PL), number of filled grains per panicle (NOFLP), Days to Maturity (DM), Test Weight (TW) and Grain Yield per Plant (GYP).
Natural screening for rice blast disease was done separately by raising selected F1 hybrids at hot spot location of Tamilnadu (Gudalur, Nilgris District) to check the disease reaction of hybrids. The mean values recorded for nine characters in the parents and F1 generation was subjected to statistical analysis. The data was subjected to combining ability analysis following the method given by Kempthrone (1957). The ratio of GCA/SCA was done for each trait to determine the predominance of additive or non-additive gene action, assuming simple additive dominance model. Standard heterosis was worked out as percent mean deviation of the mean F1 performance over the mean performance of the standard variety (ASD 16). The statistical analysis was done using the INDOSTAT statistical package (Indostat services, Hyderabad, India).
RESULTS AND DISCUSSION
The 26 rice genotypes selected based on earlier reports were subjected to natural screening at Hybrid Rice Evaluation Centre (HREC), Gudulur (hot spot for leaf blast).
| Table 2: | Rice blast disease reaction at HRE, Gudalur (Field screening) |
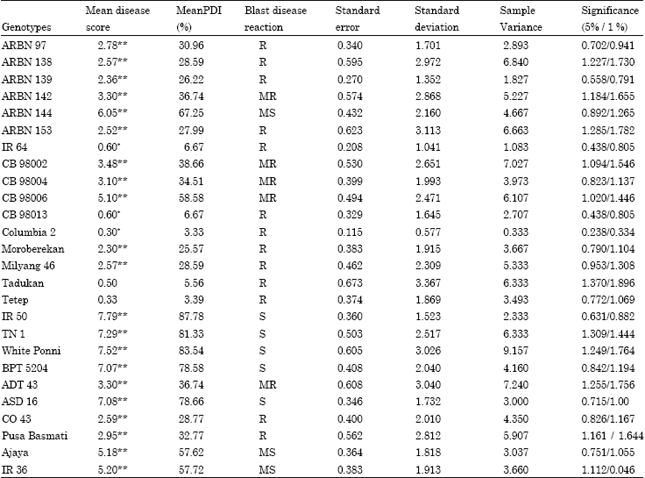 | |
| *-Significant at 5 % level; **-Significant at 1 % level, (SES, 2002). Blast disease score: 1-3.0 = R,(Resistant). 3.1-5.0 = MR(Moderately Resistant). 5.1-7.0 = MS(Moderately Susceptible). 7.1-9.0 = S (Susceptible) | |
Among the genotypes screened, highly significant lower mean disease reaction score (2.30) and mean PDI per cent (25.25) was recorded by Moroberekan, followed by ARBN 139, ARBN 153, ARBN 138 and Milyang-46 with the mean disease reaction scores and mean PDI per cent of (2.36 and 26.22%), (2.52 and 27.99%), (2.57 and 28.59%) and (2.57 and 28.59%), respectively. Significant lowest mean leaf blast disease reaction score (0.30) and mean Potential Disease Incidence percentage (3.33) was recorded by Columbia-2, followed by IR 64 and CB98013 (0.60 and 6.67%). Highly significant, higher mean disease reaction scores and mean PDI per cent was recorded by IR 50 (7.79 and 87.78%) followed by White Ponni (7.52 and 83.54%), TN 1 (7.29 and 81.33%), ASD 16 (7.08 and 78.66%) and BPT 5204 (7.07 and 78.58) (Table 2).
The same 26 genotypes were subjected to the artificial screening at Paddy Breeding Station (PBS), Coimbatore. Highly significant, lower mean disease reaction score (0.84) and mean PDI per cent (9.33) was recorded by Moroberekan, followed by Columbia 2 (0.88 and 9.77%) and ARBN 142 (1.0688 and 11.25%). Significantly lowest mean disease reaction scores and PDI% was recorded by IR 64 (0.61 and 6.81%) followed by Tadukan (0.81 and 9.03%). Highly significant, higher mean disease reaction scores was recorded by TN 1 (8.60 and 95.55%) followed by White Ponni (8.50 and 94.50), BPT 5204 (8.25 and 91.70%), ARBN 153 (7.56 and 83.99%) and ASD 16 (7.21 and 80.14%) (Table 3).
| Table 3: | Rice blast disease reaction at PBS, Coimbatore (Artificial screening) |
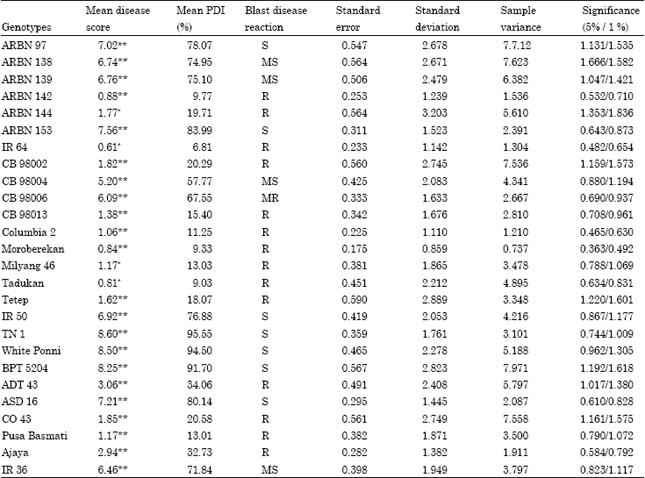 | |
| *-Significant at 5 % level; **-Significant at 1 % level (SES, 2002). Blast disease score: 1-3.0 = R (Resistant). 3.1-5.0 = MR (Moderately Resistant). 5.1-7.0 = MS(Moderately Susceptible). 7.1-9.0 = S (Susceptible) | |
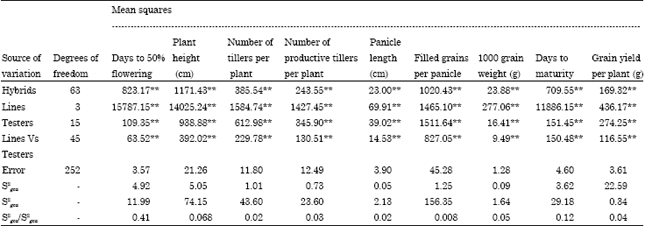
The analysis of variance has shown (Table 4) significant divergences among the parents for all the characters which can generate potential and promising hybrids. The hybrids also showed significant differences for all the characters. The contribution of lines x testers’ interaction was higher for the characters viz., number of tillers per plant, productive tillers per plant, number of filled grains per panicle and grain yield per plant. The contribution of females was higher for plant height, days to 50% flowering, days to maturity and test weight (Table 5). Analysis of variance showed highly significant differences among genotypes for all the characters. The variance due to parents, hybrids and parent - hybrid interactions were highly significant for all the nine characters. Mean squares due to females were larger in magnitude than male parents for all the characters. The magnitude of SCA variance was higher than GCA variance for all the characters except grain yield where the GCA variance was higher (Table 6).
The relative estimates of variances due to specific combining ability (S2sca) effects were predominant for all the characters studied indicating the predominance of non-additive component, except for grain yield per plant for which the variance due to general combining ability (S2gca) effects were more suggesting the preponderance of additive component of genetic variation, indicating both additive and non additive gene variations are important.
| Table 4: | Analysis of variance for Line x Tester |
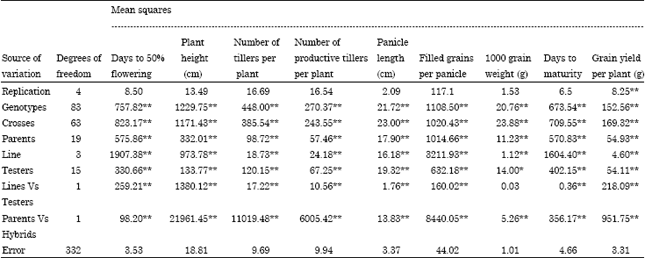 | |
| *-significant at 5% level, **-significant at 1% level | |
| Table 5: | Analysis of variance for combining ability |
 | |
| *-significant at 5% level, **-significant at 1% level | |
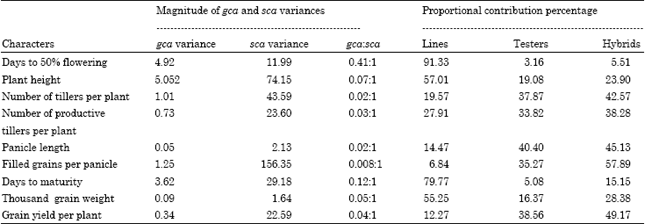
| Table 6: | Magnitude of gca and sca variances and proportional contribution to total variances due to lines, testers and hybrids for various characters |
 | |
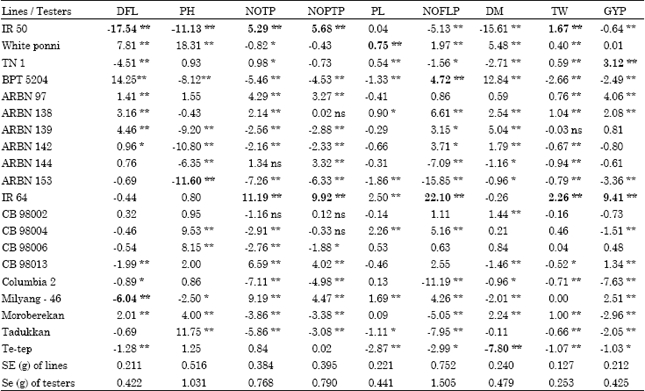
| Table 7: | General combining ability (gca) effects of lines and testers |
 | |
| *,**Means significant at 5% and 1% probability level, respectively. Bold representation indicates top 2 rankings | |
The ratio of variances (S2gca/S2sca) due to general and specific combining ability effects ranged from 0.008 to 0.41. Similar results indicating the predominance of non-additive gene action for the above mentioned traits were reported earlier by (Radhidevi et al., 2002; Annadurai and Nadarajan, 2001; Sarker et al., 2002; Ganesh et al., 2004). The presence of greater non-additive genetic variance for all the characters offers the scope for exploitation of hybrid vigour through heterosis breeding.
These observations suggest that a breeding method that can incorporate both additive and non additive genetic components would be a useful strategy. Recurrent selection method, which provides better opportunity for selection, recombination and accumulation of desired genes, should help to increase fixable as well as non-fixable types of gene effects.
The selection of parents based on per se performance may not always result in producing superior segregants. Dhillon (1975) pointed out that combining ability of parents gives useful information on the choice of parents in terms of expected performance of their progenies. In the present investigation, among the lines, IR 50 was found to be a good general combiner for six traits viz., days to 50% flowering (negative), plant height (negative) and days to maturity (negative direction). For the other traits like number of tillers per plant, number of productive tillers per plant and 1000 grain weight the same line IR 50 was found to be the good general combiner. The varieties White ponni, BPT 5204 and TN 1 recorded higher gca values for panicle length, filled grains per panicle and grain yield per plant respectively.
Among the testers, IR 64 was the good general combiner for six traits viz., number of tillers per plant, number of productive tillers per plant, panicle length, filled grains per panicle, 1000 grain weight and grain yield per plant. The testers Milyang-46, ARBN 153 and Te-tep recorded higher gca values (negative direction) for days to 50% flowering, plant height and days to maturity.
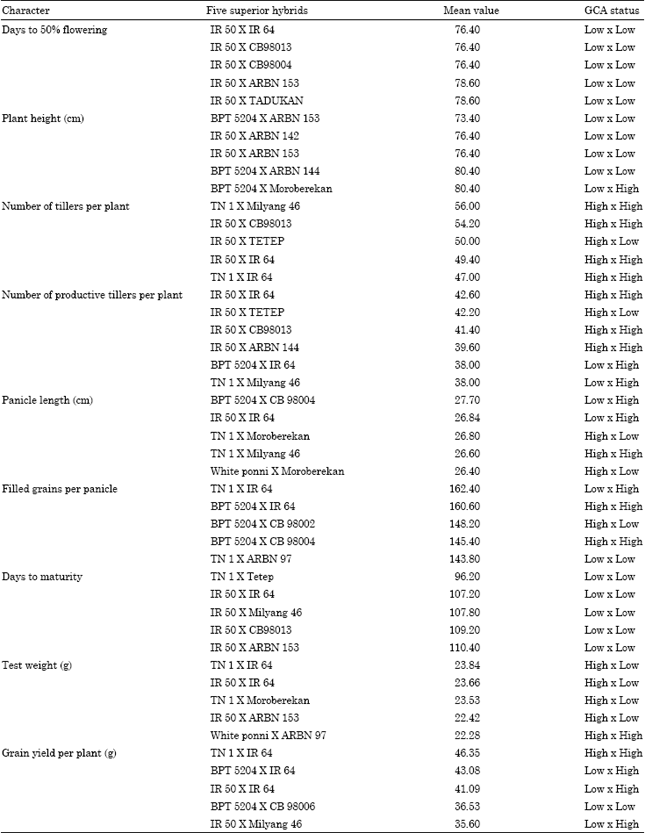
| Table 8: | Top five hybrids related with per se performance and GCA status |
 | |
The parents with high gca effect can be utilized in the hybridization programme for selection of superior recombinants in segregating progenies as suggested by Rojas and Sprague (1952).
The knowledge of combining ability coupled with per se performance of parents would be of great value in selecting suitable parents for hybridization program. In the present study, association between per se performance and gca effects was evident for most of the traits except number of tillers and number of productive tillers per plant (Table 7, 8).
Among the lines, IR 50 exhibited high per se performance along with high gca effects for days to flowering, days to maturity and plant height in the negative direction. The varieties BPT 5204 and TN I recorded high gca along with high per se performance for filled grains per panicle and grain yield per plant respectively.
Among the testers, IR 64 recorded high per se performance along with high gca effects for panicle length, filled grains per panicle, 1000 grain weight and grain yield per plant. Earlier studies also indicated that the parallelism between per se performance and gca effects does not always existed (Radhidevi et al., 2002; Suresh and Reddy, 2002; Selvaraj et al., 2006).
Generally parents with high order of per se performance for characters resulted in hybrids with high expression (Gilbert, 1958). None of the hybrid showed superior mean performance for all the traits studied. However, the hybrid IR 50xIR 64 ranked first by recording first two high mean values for 5 characters in viz., days to flowering, number of tillers per plant, panicle length, days to maturity (early) and test weight followed by TN 1xIR 64 for 3 characters with top rank viz., filled grains per panicle, test weight and grain yield per plant. The remaining hybrids showed superior performance for either one or two characters only (Table 8).
The hybrids with high mean performance for more number of traits can be utilized in hybrid breeding programmes. Hybrids with high mean performance for number of productive tillers, panicle length, test weight and grain yield per plant were observed (Radhidevi et al., 2002; Suresh and Reddy, 2002).
Thus, a high order of expression of a character by a hybrid might be attributed by the high degree of expression of either one or both of its parents in respect to the traits concerned. The sca is the deviation from the predicted value of hybrid on the basis of gca of their respective parents. The sca effect may be due to non-additive gene action.
In the present study, the first five top ranking hybrids that exhibited maximum sca effects are presented in Table 9. The ranking based on sca effects of the hybrids showed reasonable deviation from that based on the magnitude of heterosis for most of the traits. This could be expected, since the sca effects are only estimates. The large sca effect need not necessarily result in exceptional performance of a cross.
The sca effects along with per se performance of hybrids gave an idea about the practical utility of hybrid combinations for heterosis breeding. Out of 64 hybrids, significant negative sca effects were recorded for days to 50% flowering, plant height and days to maturity respectively. Negative sca for the above mentioned characters were reported earlier by Sampoornam and Thiyagarajan (1998) Radhidevi et al. (2002) and Suresh and Reddy, (2002). Totally, 16 hybrids for number of tillers and productive tillers, six hybrids for panicle length, 10 hybrids for test weight, 19 hybrids for filled grains per panicle, 10 hybrids for 1000 grain weight and 18 hybrids for yield per plant recorded significantly positive sca effects. Similar type of postive sca effects were reported by Ramalingam et al. (1997), Sampoornam and Thiyagarajan (1998) and Radhidevi et al. (2002).
Among the top five ranking hybrids, the hybrid White ponni/Tadukan showed higher sca for days to flowering, plant height, days to maturity and filled grains per panicle. The hybrid TN 1/ARBN 97 showed higher sca effects for number of tillers, number of productive tillers and panicle length.
| Table 9: | Hybrids with high sca effects |
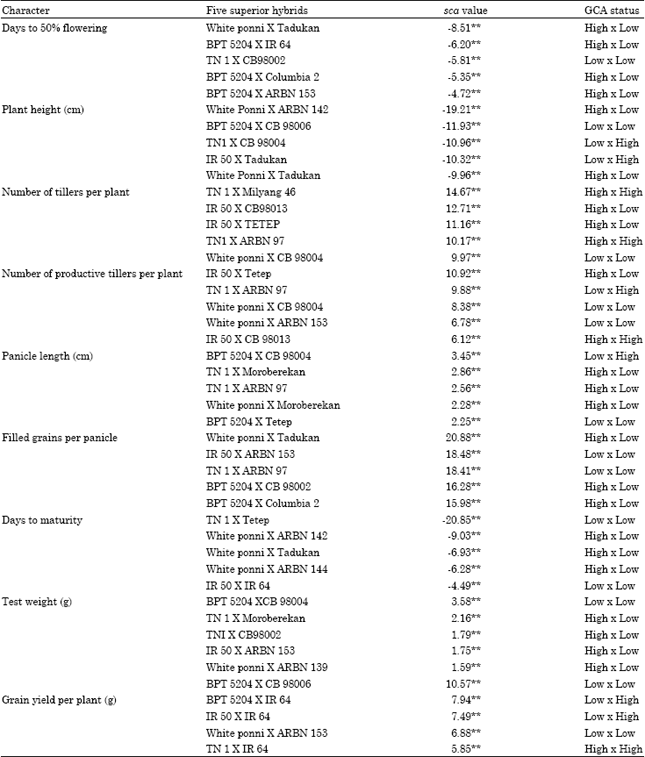 | |
The hybrids TN 1/Tetep and BPT 5204 recorded higher sca effects for test weight and grain yield per plant (Table 9).
An examination of gca effects of parents and the sca effects of the resultant hybrids revealed that, it may not be possible to find a definite trend for all the traits in all the hybrids. However, in the present study when both the parents possessed significant positive gca effects, positive sca effects were discernible in the hybrids involving them in at least a few hybrids (data not shown).
Selection of hybrids combining superior per se performance, sca and heterosis is desirable in breeding programme to get satisfactory results. In the present investigation, among the top five ranking hybrids, IR 50/IR 64 was identified as a superior hybrid as it recorded higher magnitude of standard heterosis for maximum number of characters viz., days to flowering, number of tillers per plant, number of productive tillers per plant, panicle length, test weight and grain yield per plant. It also expressed superior performance (lesser) for duration. It also had high sca for most of the important traits viz., grain yield per plant and days to maturity. This hybrid also involved parents with high gca effects. The other promising hybrids which showed superior mean performance, sca effects and standard heterosis for most of the characters studied are TN 1/ARBN 97, White ponni/Taddukan, TN 1/IR 64, BPT 5204/IR 64, BPT 5204/ CB 98006 and IR 50/Milyang 46. Parents with high x high gca effects indicating the presence of additive x additive type of gene action between favourable alleles contributed by the two parents which was considered to be of fixable nature (Subbarao and Aruna, 1997). Thus these hybrids would be very much useful for further testing.
The magnitude of heterosis is a prerequisite for development of any hybrid. Before selecting a cross on the basis of per se performance it would be worthwhile to evaluate them for hybrid vigour for various characters. Knowledge on the extent of heterosis would help in the choice of the best crosses for commercial exploitation.
IR 50/CB 98013 was the most promising combination since it showed highly significant negative relative heterosis for days to 50% flowering. IR 50/CB 98013 recorded highly significant negative heterobeltiosis and IR 50/CB 98013 and IR 50/IR 64 recorded higher standard heterosis over ASD 16. Negative heterosis for days to 50% flowering is a desirable feature as confers earliness. Negative heterosis for this trait was reported by Thirumeni and Subramanian (2000) and Radhidevi et al. (2002).
Negative heterosis for plant height is highly desirable as it confers resistance to lodging. White ponni/ARBN 142 had shown highly significant negative relative heterosis and heterobeltiosis. BPT 5204/ARBN 153 had shown the highest value of negative standard heterosis over ASD 16.
Days to maturity determines the earlier harvest of the produces and earlier marketing. Highest value of significant positive relative heterosis, heterobeltiosis and standard heterosis was recorded by the hybrid TN 1/Te-tep over the standard check ASD 16. Positive heterosis for this trait had been reported earlier by Joshi et al. (2004). Test weight is one of the most important components of yield that influences the yield conspicuously. The highest value of relative heterosis (BPT 5204/CB 98013), heterobeltiosis (TN 1/Moroberekan) and standard heterosis (TN 1/IR 64) was recorded over the standard check ASD 16. The present observations are in accordance with the findings of Yolanda and Vijendra Das (1996), Seetharamiah et al. (1999) and Thirumeni and Subramanian (2000).
Most of the hybrids recorded positive significant standard heterosis values for grain yield per plant. IR 50/IR 64 recorded highest heterosis values for relative heterosis and TN 1/IR 64 recorded high heterobeltiosis and standard heterosis over the standard check ASD 16. The present observations are in accordance with the findings of Souframanien et al. (1998) and Radhidevi et al. (2002). The hybrids with higher standard heterosis were listed in Table 10.
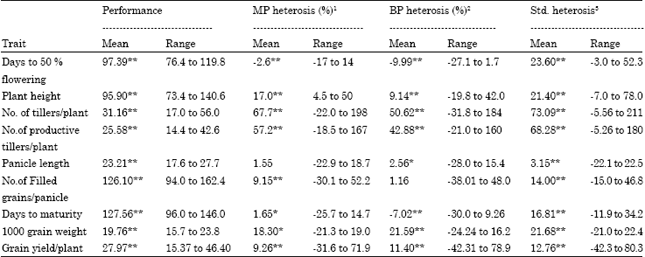
The mean values, ranges of performance and heterosis of the 64 F1 hybrids was given in Table 11. The degree of heterosis showed variation from trait to trait. For heterobeltiosis (heterosis over the better parent), No. of tillers/plant showed the highest heterosis (50.62%), followed by no.of productive tillers / plant (42.88 %), 1000 grain weight (21.59 %) and grain yield / plant (11.40%).
| Table 10: | Hybrids with desirable standard heterotic expression for different traits |
 | |
| Table 11: | Mean values, ranges of performance and heterosis among the 64 F1 hybrids |
 | |
| *,**Significant at p = 0.05 and p = 0.01, respectively. 1-Mid-parent heterosis. 2-Better-parent heterosis. 3-Standard heterosis | |
Days to flowering (-9.99 %) and days to maturity (-7.02 %) exhibited significant negative heterosis. However, better-parent heterosis of filled grains/panicle was not statistically significant although it ranged from -38 to 48%, depending on the crosses. Mid-parent heterosis for yield varied from-31.9 to 71.9 % in the hybrids. Better-parent heterosis for yield ranged between -42.31 and 78.9%. Days to 50 % flowering, when compared with other traits, exhibited a low level of heterosis. Standard heterosis was obtained by using the standard check (ASD 16) as control, which is a ruling variety for grain yield in Tamilnadu, India. Standard heterosis for yield varied from -42.3 to 80.3% in the hybrids.
Resistance to rice varieties to blast is governed mostly by dominant genes, but in few cases by recessive genes (Marchetti et al., 1987; Padmanabhan et al., 1973). An inhibitor gene was also reported by Woo (1965). Twenty six hybrids which excelled in yield parameters were screened in natural condition including the high yielding hybrids for partial leaf blast disease at Hybrid Research Evaluation Centre, Gudalur, India. Most of the hybrids were resistant except the combinations IR 50/ARBN 138 and TN 1/ARBN 138 which was susceptible. The hybrids IR 50/ARBN 97 and TN 1/ARBN 97 was moderately resistant for leaf blast reaction. The resistance in the hybrids might be due to the dominance reaction (Table 12). The parents had one or more resistance genes and the resistant gene expresses its dominance in the F1 generation. Kiyosawa and Shiyomi (1970) reported that resistance of Toride 1 was controlled by a dominant gene Pi-z (t). Yamada et al. (1976) identified a single resistant gene for blast fungus in nine cultivars. Padmanabhan et al. (1973) indicated that three dominant genes govern the resistance of the variety Zenith of which the two are complementary (Pi-z and Pi-a). The two complementary genes found in Zenith and in S.67 were allelic or same. Inheritance of resistance in Te-tep and Tadukan was found to be tri-genic. An inhibitory gene was found in the variety C.I. 5309. Kiyosawa (1981) identified 13 genes for complete resistance, none of which were recessive. Some studies, however, have identified recessive genes for blast resistance. Marchetti et al. (1987) identified a single recessive gene in the cultivar Gulfrose.
Two hybrids recorded moderate resistance reaction. Parlevliet (1988) described moderate resistance as an incomplete quantitative resistance based on minor genes.
| Table 12: | Disease reaction of high yielding hybrids to leaf blast incidence under natural screening at HRE,Gudalur |
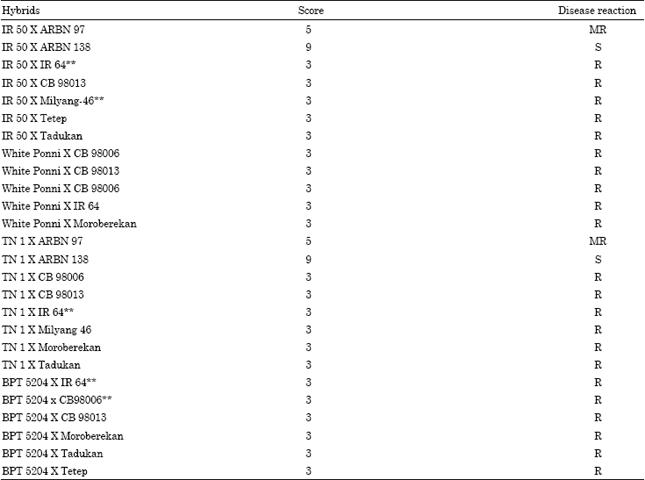 | |
| ** - indicating the crosses with high heterotic value for grain yield per plant | |
It is characterized by compatibility between the pathogen and plant with the reduced incidence of the disease. Genetic studies indicate that the partial resistance is under oligo or polygenic control and can be affected by the environment. Several researchers had also reported the role of minor genes in conferring disease resistance (Babugee and Gnanamanickam, 2000).
In conclusion, Incorporation of resistance to leaf blast is one of the important objectives of hybrid rice breeding without compromising the yield. For the success of such a breeding programme it is essential to know the variability in the disease expression of the resistant parents under varying environmental conditions and to know their genetic constitution.
REFERENCES
- Ahn, S.W. and S.H. Ou, 1982. Epidemiological implications of the spectrum of resistance to rice blast. Phytopathology, 12: 282-284.
Direct Link - Berruyer, R., H. Adreit, J. Milazzo, S. Gaillard and A. Berger et al., 2003. Identification and fine mapping of Pi33, the rice resistance gene corresponding to the Magnaporthe grisea avirulence gene ACE1. Theor. Applied Gene., 107: 1139-1147.
CrossRefPubMedDirect Link - Sallaud, C., M. Lorieux, E. Roumen, D. Tharreau and R. Berruyer et al., 2003. Identification of five new blast resistance genes in the highly blast-resistant rice variety IR64 using a QTL mapping strategy. Theor. Applied Genet., 106: 794-803.
CrossRefPubMedDirect Link - Chen, D.H., M.D. Vina, J. Inukai, D.J. Mackill, P.C. Ronald and R.J. Nelson, 1999. Molecular mapping of the blast resistance gene, Pi-44(t), in a line derived from a durable resistant cultivar. Theor. Applied Genet., 98: 1046-1053.
CrossRef - Ganesh, S.K., P. Vivekanandaan, N. Nadarajan, R.C. Babu, P. Shanmugasundaram, P.A. Priya and A. Manickavelu, 2004. Genetic improvement for drought tolerance in rice (Oryza sative L.). Proceedings of the Workshop Resilient Crops for Water Limited Environments, (WRCWLE`04), CIMMYT, Cuernavaca, Mexico, pp: 98-99.
- Hittalmani, S., M.R. Foolad, T.V. Mew, R.L. Rodriguez and N. Huang, 1995. Development of PCR-based marker to identify rice blast resistance gene Pi-2(t), in a segregating population. Theor. Applied Genet., 91: 9-14.
CrossRefDirect Link - Inukai, T., R.J. Nelson, R.S. Zeigler, S. Sarkarung and D.J. Mackill et al., 1994. Allelism of blast resistance genes in near- isogenic lines of rice. Phytopathology, 84: 1278-1283.
CrossRef - Jeon, J.S., D. Chen, G.H. Yi, G.L. Wang and P.C. Ronald, 2003. Genetic and physical mapping of Pi5(t), a locus associated with broad-spectrum resistance to rice blast. Mol. Genet. Genomics, 269: 280-289.
CrossRefPubMedDirect Link - Joshi, S.K., S.N. Sharma, D.L. Singhania and R.S. Sain, 2004. Combining ability in the F1 and F2 generations of diallel cross in hexaploid wheat (Triticum aestivam L. em. Thell). Heriditas, 141: 115-121.
CrossRef - Lawrence, I.I., R.C. Yadav, N. Huang, P.C. Ronald and F.M. Ausubel, 2000. Isolation and characterization of disease resistance gene homologous from the rice cultivar, IR 64. Gene, 255: 245-255.
PubMed - Marchetti, M.A., X. Lai and C.N. Bollich, 1987. Inheritance of resistance to P. oryzae in rice cultivars grown in United States. Genetics, 77: 799-804.
CrossRef - Padmavathi, G., T. Ram, K. Sathyanarayana and B. Mishra, 2005. Identification of blast (Magnaporthe grisea) resistance genes in rice. Curr. Sci., 88: 628-630.
Direct Link - Parlevliet, J.E., 1988. Identification and Evaluation of Quantitative Resistance. In: Plant Disease Epidemiology: Genetics, Resistance and Management, Leonard, K.J. and W.E. Fry (Eds.). McGraw-Hill, New York, pp: 215-248.
Direct Link - Sarker, U., P.S. Biswas, B. Prasad and M.A.K. Mian, 2002. Heterosis and genetic analysis in rice hybrids. Pak. J. Biol. Sci., 5: 1-5.
CrossRefDirect Link - Wang, G.L., D.J.K. Mackill, J.M. Bonman, S.R. McCouch, M.C. Champoux and R.J. Nelson, 1994. RFLP mapping of genes conferring complete and partial resistance to blast in a durably resistance rice cultivars. Genetics, 136: 1421-1434.
Direct Link - Wu, J.L., P.K. Sinha, M. Variar, K.L. Zheng, J.E. Leach, B. Courtis and H. Leung, 2004. Association between molecular markers and blast resistance in an advanced backcross population of rice. Theor. Applied Genet., 108: 1024-1032.
CrossRef - Yamada, M., S. Hiyosawa, T. Yamaguchi, T. Hirana, T. Kobayashi, K. Kushibuchi and S. Watanabe, 1976. Proposal for new method of differentiating races of Pyricularia oryzae in Japan. Ann. Phytopathol. Soc. Japan, 42: 216-219.
Direct Link - Toenniessen, G.H., J.C. O'Toole and J. DeVries, 2003. Advances in plant biotechnology and its adoption in developing countries. Curr. Opin. Plant Biotechnol., 6: 191-198.
CrossRef - Rashid, M., A.A. Cheema and M. Ashraf, 2007. Line x Tester analysis in Basmati rice. Pak. J. Bot., 39: 2035-2042.
Direct Link - Hittalmani, S., A. Parco, T.V. Mew, R.S. Zeigler and N. Huang, 2000. Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice. Theor. Applied Genet., 100: 1121-1128.
CrossRefDirect Link - Er-Ming, R., X. Yi-Long, Y. You-Jin, Z. Jie-Yun, Z. Kang-Le and L. Feng, 2005. Relationship between blast resistance phenotypes and resistance gene analogue profiles in rice. Rice Sci., 12: 75-82.
Direct Link