
S. Kundagrami
Plant Molecular and Cellular Genetics Section, Bose Institute, P 1/12 CIT Scheme VII-M, Kolkata 700054, India
J. Basak
Plant Molecular and Cellular Genetics Section, Bose Institute, P 1/12 CIT Scheme VII-M, Kolkata 700054, India
S. Maiti
Plant Molecular and Cellular Genetics Section, Bose Institute, P 1/12 CIT Scheme VII-M, Kolkata 700054, India
A. Kundu
Plant Molecular and Cellular Genetics Section, Bose Institute, P 1/12 CIT Scheme VII-M, Kolkata 700054, India
B. Das
Department of Botany, Bose Institute, 93/1 APC Road, Kolkata-700009, India
T.K. Ghose
Department of Botany, Bose Institute, 93/1 APC Road, Kolkata-700009, India
A. Pal
Plant Molecular and Cellular Genetics Section, Bose Institute, P 1/12 CIT Scheme VII-M, Kolkata 700054, India
International Journal of Plant Breeding and Genetics
Year: 2009 | Volume: 3 | Issue: 1 | Page No.: 1-10







ABSTRACT
The Mungbean Yellow Mosaic India Virus (MYMIV), transmitted through Bemisia tabaci causes severe damage in several grain legumes. Three mutant MYMIV-tolerant lines, namely, VM 1, VM 4 and VM 6, along with the susceptible Vigna mungo cultivar T9 were characterized. The objective of this study was to evaluate these three MYMIV-tolerant lines in comparison to the susceptible cv. T9 on the performance of eight agro-morphological traits. The four genotypes were grown in a randomized complete block design and combined analysis of variance over two years was carried out. The analysis of variance for individual years showed significant differences between these two genotypes for two traits in two consecutive years. However, combined analysis of variance over two years showed that except for few traits, there were no significant differences in other major traits amongst the genotypes. Genetic control of MYMIV-resistance was re-evaluated and confirmed a monogenic recessive nature. The molecular analysis revealed defect in the NB-ARC domain of putative disease resistance (R) gene in the susceptible cv.T9. While NB-ARC domains of all the MYMIV-tolerant mutant lines have common functional motifs. Presumably, the susceptibility of cultivar T9 is due to the limitation in transcript formation for the R-gene, which otherwise is a high yielding superior cultivar. Therefore, MYMIV-tolerant lines may prove useful to the plant breeders for further improvement towards sustainable agriculture.
PDF Abstract XML References Citation
How to cite this article
S. Kundagrami, J. Basak, S. Maiti, A. Kundu, B. Das, T.K. Ghose and A. Pal, 2009. Agronomic, Genetic and Molecular Characterization of MYMIV-Tolerant
Mutant Lines of Vigna mungo. International Journal of Plant Breeding and Genetics, 3: 1-10.
DOI: 10.3923/ijpbg.2009.1.10
URL: https://scialert.net/abstract/?doi=ijpbg.2009.1.10
DOI: 10.3923/ijpbg.2009.1.10
URL: https://scialert.net/abstract/?doi=ijpbg.2009.1.10
INTRODUCTION
Vigna mungo (urdbean) is one of the most popular pulses in South East Asia and a substantial source of dietary protein. Mungbean yellow mosaic India virus (MYMIV; Mayo, 2005) is a Begmovirus transmitted through the white fly, Bemisia tabaci Genn. (Nariani, 1960; Honda et al., 1983). It causes significant yield loss for many legume seeds, not only, Vigna mungo, but also, V. radiata and Glycine max throughout the South-Asian countries. Depending on the severity of the disease the yield penalty may reach up to cent percent (Basak et al., 2004). And an annual loss of 300 million US$ due to this viral infection to the leguminous crop has been projected.
Several attempts have been made in improving this leguminous crop in India. At the G.B. Plant University of Agriculture and Technology, V. radiata var. sublobata and V. mungo var. silvistris, the wild progenitors of mungbean and urdbean, were used to improve yield components and to incorporate resistance to MYMV (Singh, 1981). A genetic study involving crosses of MYMV-resistant and susceptible urdbean and mungbean (Singh, 1980, 1981; Verma and Singh, 1986) and soybean (Singh, 1988) showed two recessive genes were governed disease resistance and susceptibility to be dominant over resistance. Interspecific transfer of MYMV-resistance from V. mungo to V. radiata was initiated at the Punjab Agriculture University, India (Gill et al., 1983). The resistant character is inherited independently of the seed color and maturity (Singh, 1988). Dana (1966) attempted crossing between V. mungo (Synonym. Phaseolus mungo) and V. radiata (Synonym. Phaseolus aureus) to induce MYMV resistance in V. mungo (P. mungo), but none of the resistant progeny lines sustained the resistance trait. According to him MYMV-resistance breaks down within 10 years in the gangetic plains of Bengal, India (Dana S, personal communication). On the contrary, we are maintaining the mutant MYMIV (previously known as MYMV)-tolerant lines for over 24 years. However, considering the importance of Vignas as a pulse crop, development of MYMIV-resistant varieties is of prime importance for stabilizing the yield levels for sustainable agriculture. Additionally, introduction of virus resistant pulses in the farmer`s field would also reduce the insecticide application. In our laboratory we are maintaining six MYMIV-resistant mutant lines (VM1 to VM6) of V. mungo through selfing since 1984. These mutant lines were derived from a MYMIV-susceptible cultivar of V. mungo, T9, which has wide adaptability and higher agronomic yield, but, the crop production often suffers due to the prevalence of the viral disease. These mutant line-seeds were inbred for five generations prior to any experimentation. Of these, VM 1, VM 4 and VM 6 were tested at Indian Agricultural Research Institute (New Delhi, India) through the Agroinfection method (by infecting with T-DNA containing viral coat protein genes; Jacob et al., 2003) and the virus-tolerant nature of these lines were confirmed. The objective of this study was to evaluate performance of the three MYMIV- tolerant lines in comparison to the MYMIV-susceptible cultivar T9 based on eight agro-morphological traits under the prevailing climatic condition of Madhyamgram, North 24-paraganas, Kolkata, India; to determine inheritance pattern of MYMIV resistance and to investigate probable cause of gain-in-function mutants from the MYMIV-susceptible cultivar T9 at the molecular level.
MATERIALS AND METHODS
Plant Materials and Experimental Design
Vigna mungo L. Heppar cv. T9, a MYMIV-susceptible but agronomically superior cultivar, was collected from the Behrampur Pulses and Oilseeds Research Station (West Bengal, India). Three MYMIV-tolerant lines, VM 1, VM 4 and VM 6 and T9 were grown at the Madhyamgram Experimental Farm (22°41` N, 88° 27` E, Bose Institute, Kolkata, India) for two years (February to May, 2000 and 2001) following the Randomized Complete Block Design (RCBD) with three replications/year and five plants/replication. Combined analysis of variance of the RCBD experiments over two years was carried out with data collected on eight agro-morphological and yield traits following the procedure given by Gomez and Gomez (1984). The traits were: plant height, branches/plant, pods/plant, pod length, seeds/pod, 100-seed weight, seed yield/plant and seeds/plant.
Development of Segregating Populations for MYMIV-Reaction
The susceptible cultivar T9 (female) was crossed with the resistant line, VM6 (male), F1s produced and F2 populations were raised along with the parental lines and a resistant check.
Phenotyping the Population Segregating for MYMIV-Reaction
The lines involved in the crosses and the F1s were screened for MYMIV-reaction under field epiphytotic condition with abundant white fly population during February to May 2003.
The parental lines and the F2 populations were screened from July-September 2003, both under natural field condition and artificial/forced feeding conditions. Data on MYMIV-reaction under natural epiphytotic condition was obtained from 312 F2 plants and analysed. For forced feeding, white flies were collected from the plants and confined on a susceptible plant showing typical MYMIV symptoms for 24 h using a small, transparent glass cage with a spring cap. The same cage with the flies was attached to a healthy plant and the viruliferous insects were allowed to feed on a leaf for 24 h. After acquisition feeding, the flies were used for 3-5 transfers for inoculation feeding. Following this protocol a total number of 484 F2 plants were forced inoculated and the MYMIV-reaction data analyzed.
Statistical Analysis
Mean value of each character over five randomly selected plants in each replication was computed. The Analysis of Variance (ANOVA) table was prepared following statistical analysis (Gomez and Gomez, 1984) to find out the mean sum of square from which the Least Significance Difference (LSD) of different genotypes for each character was computed. F-tests of homogeneity of error variance of all traits were applied to combine the values of two consecutive years. For the calculation of segregation pattern of MYMIV-reaction on F2 individuals under natural condition and forced inoculated condition Chi-square test was employed to determine the probability (P) in accepting the hypothesis (expected ratio).
Genomic DNA Isolation and Detection of Polymorphism Between MYMIV-Tolerant Lines, VM1-VM6 and T9
The genomic DNA were isolated from six MYMIV-tolerant lines and cv. T9 following the method described by Basak et al. (2004). Initially several operon primers were tried but most of the cases monomorphic profiles generated. Subsequently, we have designed degenerate primers from the conserved motifs of NB-ARC domain of plant disease resistance gene in members of Fabaceae (Pal et al., 2007), referred to as Resistance Gene Analog (RGA). Isolated genomic DNA were PCR amplified with 175 RGA primer combinations (Table 1) and the amplification conditions were followed as described by Basak et al. (2004).
Cloning of Polymorphic Fragments and Sequence Analysis
Two amplified polymorphic fragments were cloned separately using pGEM-T easy vector kit (Promega, USA) following the supplied protocol and sequenced using a ABI Prism 3100 automated DNA sequencer. The polymorphic markers were named as Vigna mungo resistance gene homolog of VM6 (VMYR6) and Vigna mungo susceptible allele 1 (VMYS1). Nucleotide sequence and in silico translated peptide-sequence similarities between the VMYR6 and VMYS1 and other published sequences were determined by screening the GenBank non-redundant database using the computer program NBLAST (Basic Local Alignment Search Tool, Altschul et al., 1997). To ascertain the presence of ORF and conserved domain(s) in VMYR6 and VMYS1, the NCBI ORF finder (http://www.ncbi.nlm.nih.gov/projects/gorf/) and conserved domain (http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml) databases were also searched.
Sequence Alignment and Database Search
Sequences (VMYR6 and VMYS1) were aligned using CLUSTALW (ver 1.83) (Thompson et al., 1994) with the default settings of gap opening penalty (10.0) and gap extension penalty (0.1). The Gannet 250 protein weight matrix has been used.
| Table 1: | RGA primers and respective annealing temperatures |
 | |
RESULTS
Agro-Morphological Traits of MYMIV-Susceptible and -Tolerant Genotype
Significant differences between the MYMIV-susceptible and -tolerant genotypes for characters like, branches/plant, pods/plant and seeds/pod were evident, while no significant differences were noted for rest of the traits among individuals of these genotypes (Table 2). The mean and LSD values for the eight agro-morphological traits are shown in Table 2. The results of the analysis of variance of the RCBD experiments for individual years and the combined analysis of variance over two years are shown in Table 3. The analysis of variance for individual years showed significant differences amongst the genotypes for two different traits in two subsequent years (Table 3). In the first year, seeds/plant was significantly different at 1% level. In the second year, branches/plant was significantly different at the 5% level. However, combined analysis by F-test for homogeneity of error variances revealed that all traits, except for branches/plant (significant at 5 and 1% due to the genotype), pods/plant and seeds/pod (significant at 5% due to the genotype), were homogeneous amongst the MYMIV-tolerant lines and the susceptible cultivar T9 (Table 3). The tolerant lines and the susceptible T9 cultivar have since been used for crossing to generate populations segregating for MYMIV-reaction (Basak et al., 2004).
| Table 2: | The mean of the eight agro-morphological traits of the MYMIV-susceptible cultivar T9 and -tolerant lines, VM 1, VM 4 and VM 6 |
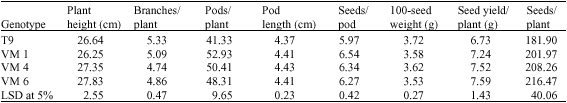 | |
| Table 3: | Analysis of variance of the RCBD experiments for individual years and the combined analysis of variance over two years |
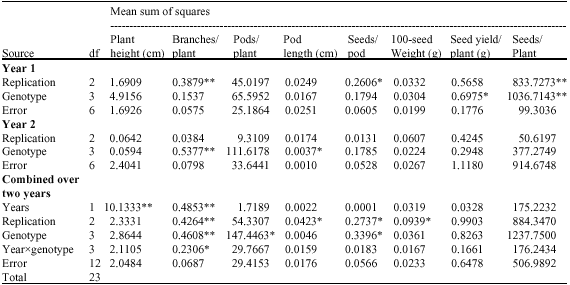 | |
| *and **Significant at 5 and 1% levels, respectively | |
Inheritance of MYMIV-Resistance Trait
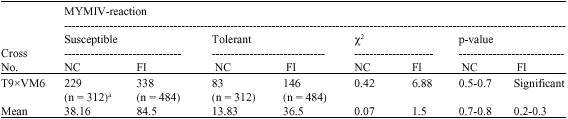
In this research, the inheritance of MYMIV-resistance in crosses of V. mungo was studied. Segregation into tolerant and susceptible individuals in progenies of the cross between tolerant line, VM6, with the susceptible cv. T9 showed expression of one R gene in the tolerant individuals studied both under natural and artificial screening conditions and the data indicates that segregation fits into 3:1 ratio. The segregating pattern showed MYMIV-resistance to be monogenic recessive (Table 4). Data on MYMIV-reaction under natural epiphytotic condition from F2 plants are shown in Table 4 and analyzed, which corroborated with monogenic recessive control.
Generation of Polymorphism Between MYMIV-Tolerant and -Susceptible Genotypes
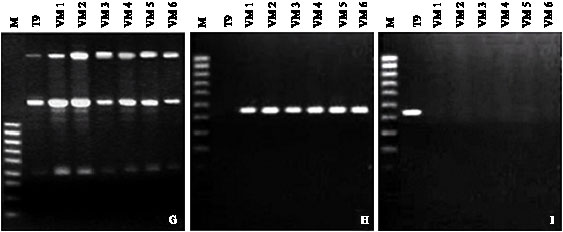
Out of 175 pairs of RGA primer combinations used so far, most of the primer combinations produced monomorphic amplification profiles (Fig. 1A-G). While, only the combination of RGA-1F-CG (5`-AGTTTATAATTCGATTGCT-3`) and RGA-1R (5`-ACTACGATTCAAGACG TCCT-3`) generated one polymorphic fragment in all the 6 MYMIV-tolerant lines (Fig. 1H), whereas, no amplification product was obtained in the MYMIV-susceptible cultivar T9 of V. mungo. While, a degenerate primer RGA-1F-TG (5`-AGTTTATAATTTGATTGCT-3`) with RGA-1R generated one polymorphic fragment only in cv. T9 of V. mungo but not in MYMIV-tolerant genotypes (Fig. 1I).
| Fig. 1: | Monomorphic (A-G) and polymorphic (H, I) amplification profiles generated from genomic DNA of susceptible cv. T9 and six MYMIV-tolerant mutant lines VM1-VM6, employing different RGA primer combinations: RGA 11 F-G/RGA 10 R-AA, RGA 2 F-TC/RGA 2 R, RGA 8 F-G/RGA 8 R, RGA 11 F-G/RGA 10 R-AG, RGA 10 F-G/RGA 10 R-GA, RGA 6 F/RGA 6 R, RGA 11 F-G/RGA 10 R-GG, RGA 1 F-CG/RGA 1 R and RGA 1 F-TG/RGA 1 R |
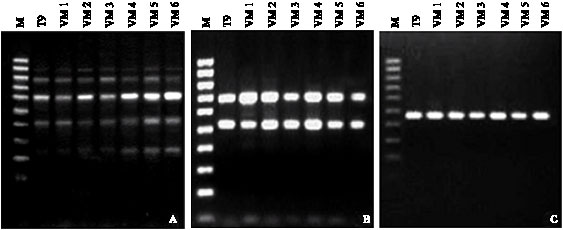
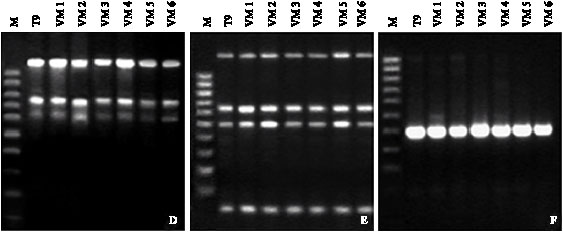
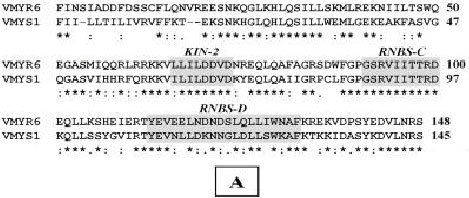
| Fig. 2: | (A) Pairwise nucleotide sequence alignment between VMYR1 and VMYS6 using CLASTALW software. Reported conserved motifs of NB-ARC domain are highlighted and (B) Nucleotide sequence of VMYS1 and in silico translated aminoacid sequences showing the presence of stop codons within the ORF. * = Denotes to the identity, : = Strongly similar, . = Weakly similar |

| Table 4: | MYMIV-reaction of individuals of F2 segregating population under Natural Condition (NC) and Forced-Inoculat (FI) conditions |
 | |
| an: No. of samples | |
Analysis of Nucleotide Sequences of Marker Fragments
Sequence analysis followed by sequence alignment of VMYR6 and VMYS1 showed sequence similarity (Fig. 2A) and these sequences also have homology with other NB-ARC domains (results not shown). Majority of these accessions were either plant R genes or R gene homologues. Both of these sequences also have high sequence similarity with the MYMIV-resistance linked marker VMYR1 of V. mungo (Accession No. AY297425, Basak et al., 2004). Similarities between VMYR1 and VMYR6 and between VMYR1 and VMYS1 represented by E-value (expected frequency) = 0 and 4e-90, respectively. The conserved domain search revealed that VMYR6 sequence is a part of the NB-ARC domain containing conserved reported motifs (Pal et al., 2007). In silico translated amino acid sequence obtained from the nucleotide sequence of VMYS1 revealed the presence of stop codons within the ORF (Fig. 2B).
DISCUSSION
Agro-Morphological Traits of MYMIV-Susceptible and -Tolerant Genotype
The MYMIV-tolerant lines produced more seeds/pod than those produced by the T9. The results also showed that the variation was due to year (environment) and the year X genotype interactions were insignificant (Table 3). These results clearly show that in terms of agro-morphology and yield traits the MYMIV-tolerant lines were statistically nearly identical to the MYMIV-susceptible T9 cultivar.
Inheritance of MYMIV-Resistance Trait
Assuming Mendelian inheritance, the almost perfect fit to a ratio of 3:1 (susceptible: resistant) for segregating progenies under natural condition suggest the monogenic recessive control of MYMIV-resistance in V. mungo mutant line, VM6 (Table 4). Inheritance of resistance to MYMIV was studied in crosses of mungbean, blackgram and their interspecific crosses with V. sublobata (Singh, 1980, 1988). Resistance to MYMV was recessive in the three Vigna species. The segregation ratios in F2 and back crosses indicated that the resistance was digenic recessive in the crosses of mungbean and in interspecific crosses of mungbean with blackgram and V. sublobata but MYMV-resistance was monogenic recessive in blackgram crosses. Frisch and Melchinger (2001) reported that several important genes in breeding for resistance and quality traits are inherited recessively. Especially, resistance traits for plant viruses has been reported to be recessive in crop plants (Park and Tu, 1991; Pal et al., 1991; Miklas et al., 2000; Diaz-Pendon et al., 2004; Hayes et al., 2004; Ritzenthaler, 2005). The significant p-value from the expected 3:1 segregation for MYMIV-reaction in the F2-population under forced feeding condition (Table 4) was probably due to the following reasons:
| • | Experimental error due to the non-viruliferous nature of the white flies |
| • | Vectors may not feed on the plant during the feeding period |
| • | Vectors were weak and incapable of transmit the virus |
Monogenic recessive nature of the genetic control for MYMIV-tolerance was also reported earlier and that was re-confirmed by phenotypic segregating-F2 progenies of a third cross in the present investigation. By understanding the genetic basis of the MYMIV-reaction trait and the allelic variation at the locus, the breeder would be able to design superior genotypes of V. mungo.
Probable Cause of Gain-in-Function Mutants from the MYMIV-Susceptible cv. T9
The findings of the present study corroborate with our contention that the MYMIV-tolerant plant arose due to the natural mutation/s in the susceptible T9 genome. Firstly, the collection and cultivation history of the T9 genotype was in favour of the natural mutation hypothesis (Basak et al., 2004). The combined analysis of variance over two years presented have shown that the tolerant and susceptible genotypes are homogeneous with respect to most of the yield related traits. Secondly, except two RGA primer pairs, all the rest primers tried so far produced monomorphic banding profiles from the genomes of the MYMIV-tolerant lines and the susceptible cv. T9. It indicates that most probably the tolerant lines arose through a natural mutation in the genome of the susceptible cultivar T9 and have high genomic homology except for a small portion of the genome including R gene/s; which is evident from the differential MYMIV disease reaction.
It is further evident from the analysis of marker fragment (VMYS1), generated from within the NB-ARC domain (which is a novel signaling-motif shared by plant R gene products and regulators of cell death in animals) of the susceptible genotype, that it is a pseudo-ORF and such no transcripts was found even after challenging with the virus (result not shown). Whereas, perhaps due to the spontaneous mutation, the tolerant genotypes salvaged the function; which is also evident from the presence of the transcripts of VMYR6 after challenging with the virus. Therefore, it is assumed that the tolerant genotypes are gain-in-function mutants.
CONCLUSION
Chemical pesticides and insecticides are commonly applied in the farmer`s field to protect crop plants from the attack of pathogens. The extensive use of these toxic chemicals not only forced the insects to build up resistance and new biotypes, but also adversely affects the ecological balance and natural pest controlling agents. Cultivation of biotic stress tolerant varieties endowed with favoured allele, like R-gene are globally preferred to keep the environment free from chemical and toxic pollutants and to sustain ecological balance. The V. mungo cultivar T9 is a superior genotype with high agronomic yield and cultivated at different states of India. MYMIV-tolerant lines derived from the T9 genotype would prove useful for farming and also for further improvement of V. mungo.
ACKNOWLEDGMENTS
We thank the Department of Biotechnology, India, for financial assistance (BT/PRO 0689/AGR/07/27/97 and BT/01/COE/06/03) and research fellowships to J.B, SK and A.K. We are thankful to Dr. Aparajita Mitra for the identification of the mutant lines from the MEF, Bose Institute.
REFERENCES
- Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Basak, J., S. Kundagrami, T.K. Ghose and A. Pal, 2004. Development of Yellow Mosaic Virus (YMV) resistance linked DNA marker in Vigna mungo from populations segregating for YMV-reaction. Mol. Breed., 14: 375-383.
CrossRefDirect Link - Dana, S., 1966. Cross between Phaseolus aureus Roxb. and P. mungo L. Genetica, 37: 259-274.
CrossRefDirect Link - Diaz-Pendon, J.A., V. Truniger, C. Nieto, J. Garcia-mas, A. Bendahmane and M. Aranda, 2004. Advances in understanding recessive resistance to plant viruses. Mol. Plant Pathol., 5: 223-233.
CrossRefDirect Link - Frisch, M. and A.E. Melchinger, 2001. Marker-assisted backcrossing for introgression of a recessive gene. Crop Sci., 41: 1485-1494.
Direct Link - Gill, A.S., M.M. Verma, H.S. Dhaliwal and T.S. Sandhu, 1983. Interspecific transfer of resistance to mungbean yellow mosaic virus from Vigna mungo to Vigna radiata. Curr. Sci., 52: 31-33.
Direct Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Hayes, A.J., S.C. Jeong, M.A. Gore, Y.G. Yu, G.R. Buss, S.A. Tolin and M.A. Saghai Maroof, 2004. Recombination within a nucleotide-binding-site/leucine rich-repeat gene cluster produces new variants conditioning resistance to soybean mosaic virus in soybeans. Genetics, 166: 493-503.
Direct Link - Honda, Y., M. Iwaki, Y. Saito, P. Thongmeearkom, K. Kittisak and N. Deema, 1983. Mechanical transmission, purification and some properties of whitefly-borne mung bean yellow mosaic virus in Thailand. Plant Dis., 67: 801-804.
Direct Link - Jacob, S.S., R. Vanitharani, A.S. Karthikeyan, Y. Chinchore, P. Thilaichidambaram and K. Veluthambi, 2003. Mungbean yellow mosaic virus-Vi Agroinfection by codelivery of DNAA and DNAB for one Agrobacterium strain. Plant Dis., 87: 247-251.
CrossRefDirect Link - Miklas, P., R. Larsen, R. Riley and J.D. Kelly, 2000. Potential marker-assisted selection for bc-1 2 resistance to bean common mosaic potyvirus in common bean. Euphytica, 116: 211-219.
CrossRefDirect Link - Pal, A, A. Chakrabarti and J Basak, 2007. New motifs within the NB-ARC domain of R proteins: Probable mechanisms of integration of geminiviral signatures within the host species of Fabaceae family and implications in conferring disease resistance. J. Theor. Biol., 246: 564-573.
CrossRefDirect Link - Pal, S.S., H.S. Dhaliwal and S.S. Bains, 1991. Inheritance of resistance to yellow mosaic virus in some Vigna species. Plant Breed., 106: 168-171.
CrossRefDirect Link - Park, S.J. and J.C. Tu, 1991. Inheritance and allelism of resistance to a severe strain of bean yellow mosaic virus in common bean. Can. J. Plant Pathol., 13: 7-10.
Direct Link - Ritzenthaler, C., 2005. Resistance to plant viruses: Old issue and new answers? Curr. Opin. Biotechnol., 16: 118-122.
CrossRefDirect Link - Singh, D.P., 1980. Inheritance of resistance to yellow mosaic virus in blackgram (Vigna mungo (L.) Hepper). Theor. Applied Genet., 57: 233-235.
CrossRefDirect Link - Singh, D.P., 1981. Breeding resistance to disease in green gram and black gram. Theor. Applied Genet., 59: 1-10.
CrossRefDirect Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Verma, R.P.S. and D.P. Singh, 1986. The allelic relationship of genes giving resistance to mungbean yellow mosaic virus in blackgram. Theor. Applied Genet., 72: 737-738.
CrossRefDirect Link