
Dandong Hu
School of Life Science, Northwest Normal University, Lanzhou, Gansu 730070, China
Yujuan Cui
School of Life Science, Northwest Normal University, Lanzhou, Gansu 730070, China
Ji Zhang
School of Life Science, Northwest Normal University, Lanzhou, Gansu 730070, China
LiveDNA: 86.37984
International Journal of Pharmacology
Year: 2022 | Volume: 18 | Issue: 6 | Page No.: 1309-1321







ABSTRACT
Background and Objective: Parkinson’s disease (PD) is currently the most common disease with a high incidence worldwide. However, there are few studies regarding the role of neuronic acid (NA) in PD. So this study aimed to explore the intervention effect of NA and further understand its influence on the disease to provide a reliable theoretical basis for future clinical practice. Materials and Methods: Human neuroblastoma cells (SH-SY5Y) and adrenal pheochromocytoma (PC-12) were induced to establish PD cell models, which were assigned to high- (20 μmol L–1 NA), medium- (10 μmol L–1 NA) and low-dose (5 μmol L–1 NA) groups, as well as model group (normal saline intervention) respectively. Normal SH-SY5Y and PC-12 were used as controls. Levels of oxidative stress (OS) and inflammation in cells were measured by PCR and lactate dehydrogenase (LDH) release was analyzed. Western blot was used to determine Ras/MEK/ERK axis-related protein expression and apoptosis. Results: Inflammatory factors (IFs) and OS were inhibited and LDH release decreased in the three groups of cells intervened by NA. In addition, the model group showed markedly enhanced apoptosis, while NA intervened cells presented reduced apoptosis and increased Ras/MEK/ERK axis-related protein expression. After inhibiting the Ras/MEK/ERK axis, the oxidative stress response (OSR), IFs, LDH release and apoptosis of PD cells increased obviously. In the rescue experiment, inhibiting the Ras/MEK/ERK axis completely reversed NA’s effects on PD model cells. Conclusion: NA suppresses PD cell apoptosis via activating the Ras/MEK/ERK axis and alleviates the OS injury and inflammatory damage of cells.
PDF Abstract XML References Citation
Copyright: © 2022. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Dandong Hu, Yujuan Cui and Ji Zhang, 2022. Influence of Nervonic Acid on Parkinson’s Disease Model Cells through Ras/MEK/ERK Axis. International Journal of Pharmacology, 18: 1309-1321.
DOI: 10.3923/ijp.2022.1309.1321
URL: https://scialert.net/abstract/?doi=ijp.2022.1309.1321
DOI: 10.3923/ijp.2022.1309.1321
URL: https://scialert.net/abstract/?doi=ijp.2022.1309.1321
INTRODUCTION
At present, cardio-cerebrovascular diseases have become the number one killer endangering the normal living ability and life safety of the middle-aged and elderly. The typical feature of cardio-cerebrovascular diseases is vascular malignant lesions, which oppress and affect peripheral nerve tissues, causing a series of adverse reactions1. Among them, Parkinson’s disease (PD) is a very common type of cerebrovascular neurodegeneration and also the most commonly seen disease that affects the normal life of the elderly at present2. PD manifests as characteristic motor symptoms, including static tremor, bradykinesia, muscle rigidity and balance disorder, while patients with severe diseases may develop neurasthenia and cognitive impairment3. On average, there are 1-3 PD patients in every 10,000 elderly people over the age of 60, according to the study4. And in some countries with serious ageing and high population density (such as China and India), the incidence of PD among people aged 60 even reaches about 1-2%5. Combined with previous investigations, it is found that the incidence of PD has shown an obvious upward trend in recent years, which has increased by about 6-10 times compared with a decade ago6. Although the specific pathogenesis of PD is not clear, it is considered clinically that degeneration and death of dopaminergic neurons in substantia nigra is the key to the occurrence of PD, while ageing, genetic susceptibility and environmental toxins may be the predisposing factors7. At this stage, the clinical treatment can only ameliorate the symptoms of PD, but can't hinder the progression of the disease, let alone completely cure PD8. And though PD does not pose a great death threat, it has a particularly serious impact on the normal life of patients. Consequently, it is particularly important to fully understand the pathogenesis of PD and seek new and effective treatment methods.
Neuronic acid (NA), a monounsaturated fatty acid, was first found in mammalian nerve tissue9. NA, rich in brain and nerve tissue, is a critical component of biofilm, which is usually used as a marker of the medulla (white matter) in the cerebroside and is an essential nutrient for the growth and redevelopment of nerve cells and the maintenance of physiological functions10. It is also recognized as the first and only double-effect magical substance that can repair and dredge the neural pathway of the damaged brain and promote the regeneration of nerve cells11. Previous evidence has pointed out that NA, which can significantly ameliorate neurological diseases such as encephalomyelitis and leukoaraiosis, is considered a breakthrough in the future treatment of nervous system diseases12,13. However, the research on the effect of NA on PD is difficult to track in the existing literature, in addition to the study by Hu et al in May 2021 showing that NA can effectively relieve the symptoms of PD mice14 and its specific mechanism remains to be further defined.
Reviewing previous studies, we found that Ras/MEK/ERK is an axis that plays an important role in nervous system diseases, showing abnormal activation in Alzheimer’s disease and schwannoma15. Furthermore, this axis is also reported as one of the key links in promoting nerve injury in PD, which is of great significance16. Therefore, we believe that NA may participate in PD via the Ras/MEK/ERK axis.
To confirm our ideas and further understand NA’s influence on PD and the mechanism, this research explored the intervention effect of NA through the preparation of PD cell models, to provide a reliable theoretical basis for future clinical practice.
MATERIALS AND METHODS
Study area: The study was carried out at the School of Life Science, Northwest Normal University, China, from January, 2021-February, 2022.
Cell data: Human neuroblastoma cells (SH-SY5Y) and adrenal pheochromocytoma (PC-12) both offered by ATCC were cultivated (37°C) in the medium (MEM/F12+10% FBS+1% P/S) in a 5% CO2 incubator and the culture medium was changed every 3 days.
PD cell model establishment: PD cell models were established by inducing SH-SY5Y and PC-12 with 1-methyl-4-phenyl pyridine ion (MPP+). Referring to the research of liping Bai et al.17, SH-5Y5Y and PC-12 in a logarithmic growth period were intervened with MMP+ of 250 μmol L–1 for 72 hrs to obtain PD model cells SH-SY5Y-MPP+ and PC-12-MPP+.
NA intervention: SH-SY5Y-MPP+ and PC-12-MPP+ were divided into high-, medium- and low-dose groups, as well as model group and control group respectively. Referring to the research by Vozella et al.18, the high-, medium- and low-dose groups were intervened by 20, 10 NA and 5 μmol L–1 NA, respectively, while the control group and the model group were treated with the same amount of normal saline. All groups were treated for 24 hrs and then the medium was replaced for further culture.
PCR detection: Total RNA of cells, extracted by Trizol, was reverse transcribed into cDNA for PCR amplification. The primer sequences were detailed in Table 1.
| Table 1: | Primer sequences |
| F (5'-3') | R (5'-3') | |
| IL-6 | GATGTTGCTGCTTCACTTC | CCTTGTTGGCTTATGTTCTG |
| IL-8 | GGGCTGCATCTAAAGTAAATGG | CAGAACACTGCTGTAGAAGGTA |
| TNF-α | CTCTTCTCATTCCTGCTTG | CTCCACTTGGTGGTTTGCT |
| SOD | CACAACTGGTTCACCGCTTG | GCCCAACCAGACAGAGAATGA |
| MDA | CGTGCAATCAGTTCGGACC | CCAGGCATCTCCCTTCCATTC |
| β-actin | CTAAGGCCAACCGTGAAAAG | ACCAGAGGCATACAGGGACA |
The amplification conditions were 94°C for 30 sec, 94°C for 5 sec and 60°C for 30 sec and 2–ΔΔct was used to calculate the relative expression.
Lactate dehydrogenase (LDH) release rate detection: LDH-Cytotoxicity Assay Kit (Beyotime, Shanghai, China) was used to detect LDH release and a microplate reader (Thermo Fisher Scientific, Beijing, China) to determine absorbance (490 nm).
Western blot detection: RIPA lysed cells to extract the total protein, which was then transferred to a PVDF membrane by SDS-PAGE (12%), sealed with nonfat-dried milk (5%) for 2 hrs and closed overnight (4°C) after adding the I antibody of the protein to be tested. After the removal of the I antibody, the membrane was immersed in the horseradish peroxidase-labelled goat anti-rabbit secondary antibody for 1 hr cultivation (37°C) and 18 luminescence reagent (Shanghai Youmede Technology Co., Ltd., Shanghai, China).
Flow cytometry: The trypsin digested cells were immersed in 100 μL binding buffer to prepare a cell suspension of 1×106 cells mL–1, which was then added with Annexin V-FITC and PI in turn for a 5 min light-tight incubation (indoor temperature). The apoptosis rate was determined by flow cytometry.
Intervention of Ras/MEK/ERK axis: MEK/ERK selective inhibitor U0126 (20 μM) was used to treat SH-SY5Y-MPP+ and PC-12-MPP+ for 48 hrs as the intervention group and a blank group intervened with the same amount of normal saline was set up. Cell activity was detected according to the above method.
Rescue experiment: SH-SY5Y-MPP+ and PC-12-MPP+ were treated with NA and U0126 simultaneously (the dosage of NA used was the dose with the best intervention effect) as group A and the cells intervened by NA alone and normal saline was set as groups B and C, respectively. The cellular activity was measured according to the above-mentioned method.
Statistical processing: Statistical processing was conducted with the use of SPSS 22.0 software. All tests were repeated 3 times and the results were averaged as (Mean±Standard deviation). Independent t-test was used for inter-group comparison, repeated analysis of variance and LSD posthoc test was used for multi-group comparison, with the difference deemed remarkable when p<0.05.
RESULTS
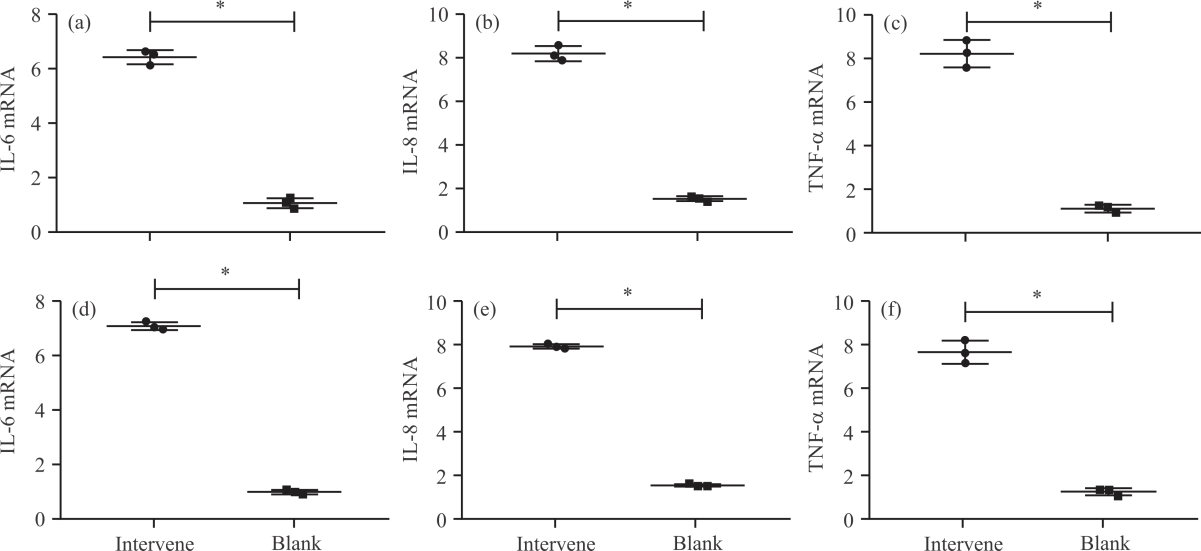
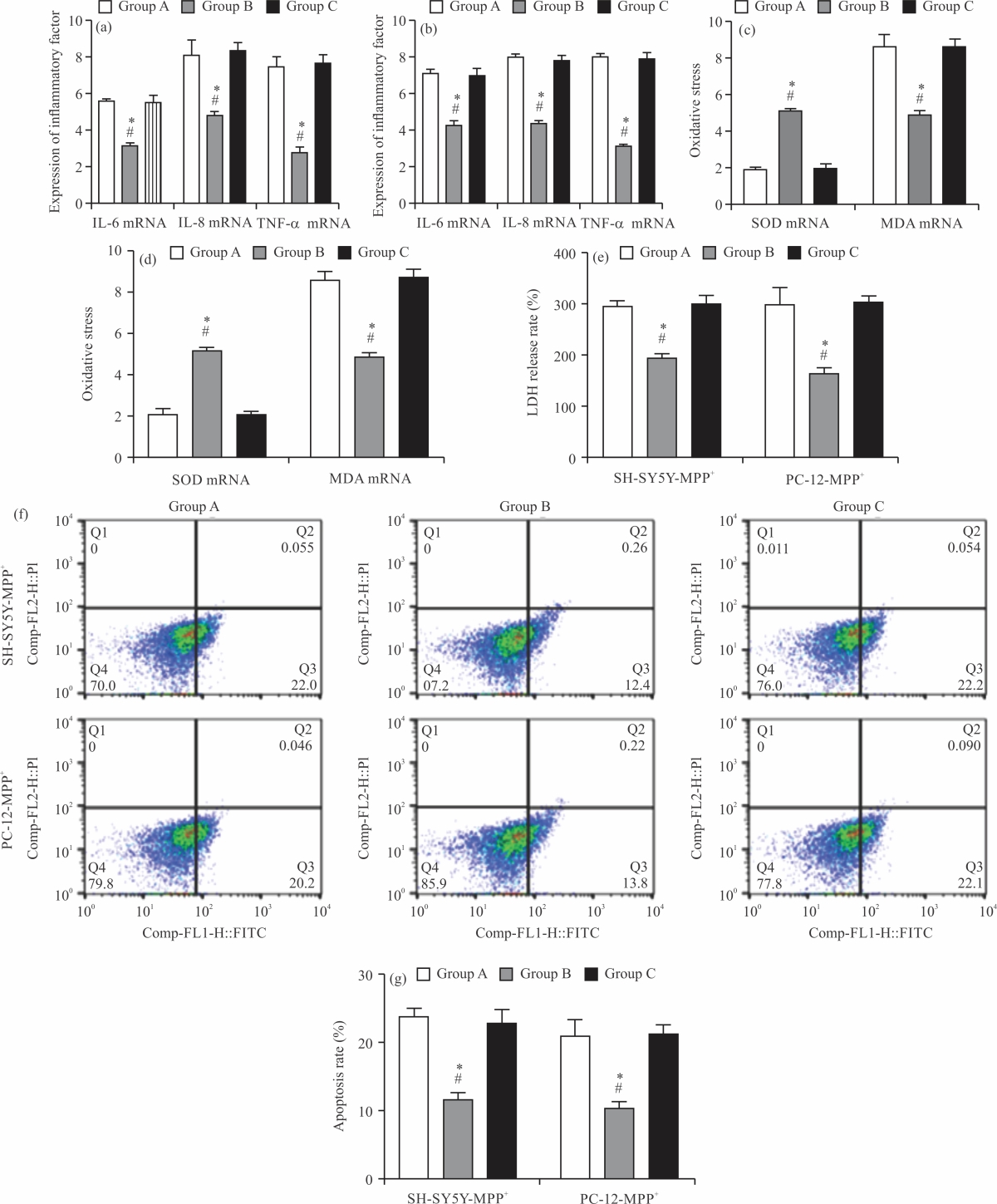
Impact of NA on inflammatory responses in PD cell models: First, the levels of inflammatory factors (IFs) in the PD cell model were examined. The results showed that the mRNA levels of IL-6, IL-8 and TNF-α were the highest in the model group among the five groups (p<0.05), with their levels in SH-SY5Y-MPP+ cells being (5.71±0.25, Fig. 1a), (8.03±0.66, Fig. 1b) and (7.47±0.58, Fig. 1c) respectively and those in PC-12-MPP+ cells being (7.16±0.29, Fig. 1d), (7.90±0.23, Fig. 1e) and (7.81±0.68, Fig. 1f) respectively, while these IFs showed the lowest levels in SH-SY5Y-MPP+ and PC-12-MPP+ cells in the control group (p<0.05). Among the three groups intervened by NA, the levels of IFs were reduced to varying degrees compared with the model group. The levels of IL-6, IL-8 and TNF-α were the lowest in the high-dose group and the highest in the low-dose group, with those in the medium-dose group in between (p<0.05). It can be seen that there are obvious inflammatory responses in the PD cell model and the use of NA can inhibit the inflammatory process.
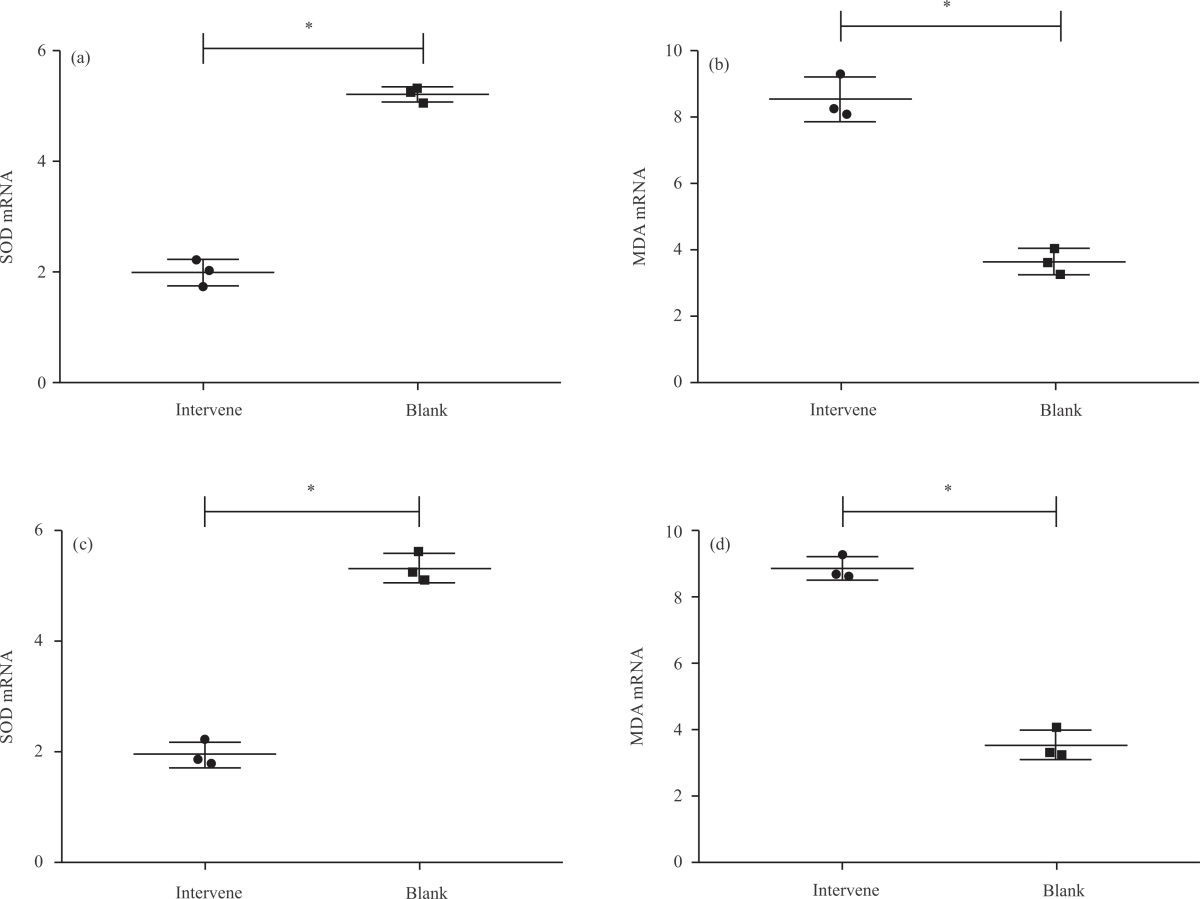
Impact of NA on oxidative stress (OS) in PD cell models (Fig. 2a-d): Subsequently, we detected the oxidative stress response (OSR) indexes SOD and MDA in the PD cell model to evaluate the OSR of cells. After detection, it was found that SOD mRNA levels in SH-SY5Y-MPP+ and PC-12-MPP+ cells in the model group were the lowest among the five groups (p<0.05), followed in ascending order by low- and medium-dose groups (p<0.05), The SOD mRNA levels in the high-dose group were not different from those in the control group (p>0.05), higher than the other three groups (p<0.05, Fig. 2a-c). The MDA mRNA levels in SH-SY5Y-MPP+ and PC-12-MPP+ cells were the highest in the model group [(1.72±0.46), (2.05±0.34)] among the five groups and the lowest in the control group (p<0.05).
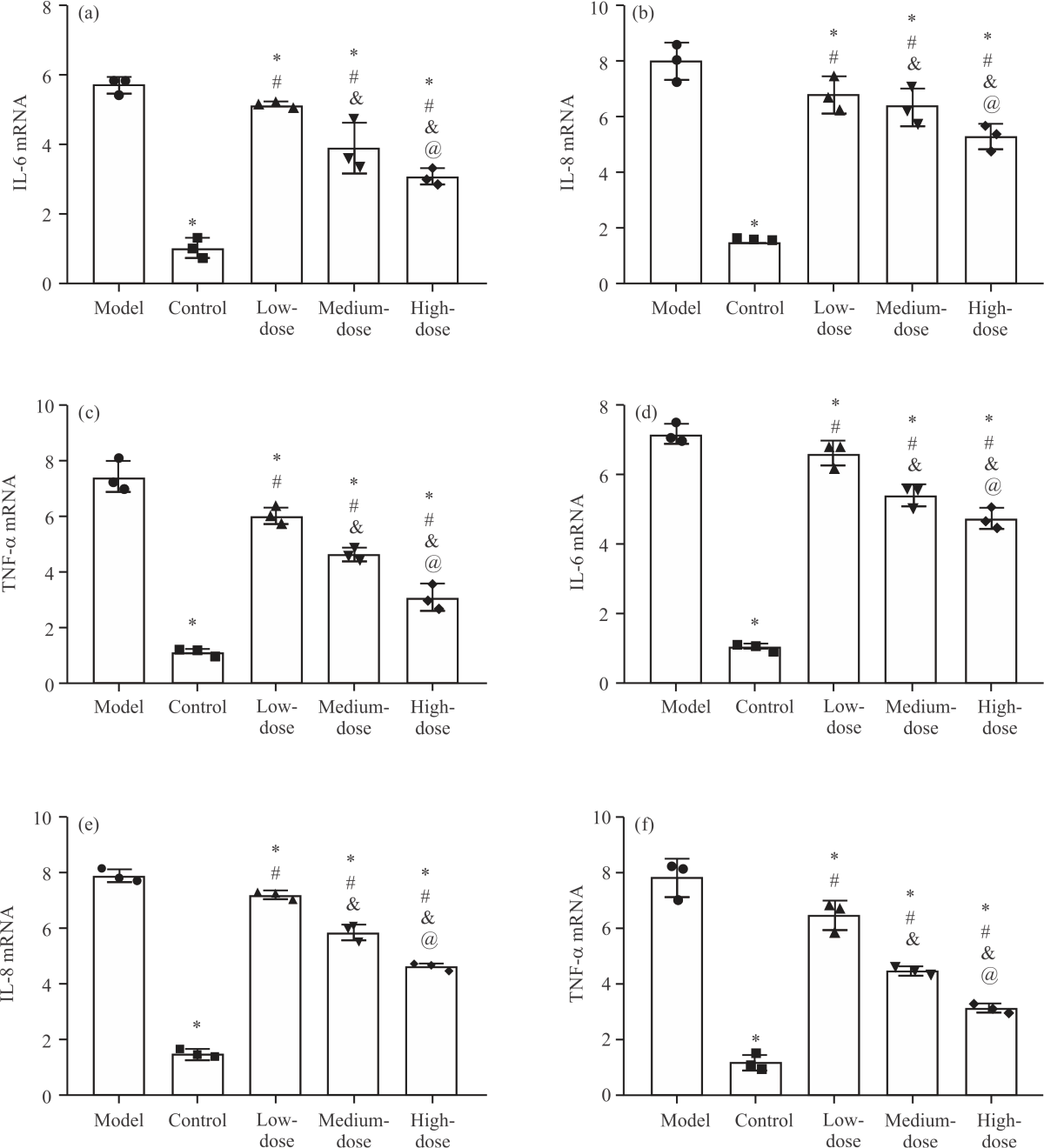 |
| Fig. 1(a-f): | Impact of NA on inflammatory responses in PD cell models, (a) IL-6 mRNA level in SH-SY5Y-MPP+, (b) IL-8 mRNA level in H-SY5Y-MPP+, (c) Comparison of TNF-α mRNA levels in H-SY5Y-MPP+, (d) IL-6 mRNA level in PC-12-MPP+, (e) IL-8 mRNA level in PC-12-MPP+ and (f) Comparison of TNF-α mRNA levels in PC-12-MPP+ *Model group, p<0.05, #Control group, p<0.05, &Low-dose group, p<0.05, @Medium-dose group, p<0.05 and X-axis: Groups |
Among the three NA intervention groups, MDA mRNA levels were the lowest in the high-dose group, followed in ascending order by medium- and low-dose groups (p<0.05, Fig. 2b-d). The results suggest that there is also obvious oxidative stress in the PD cell model, which is significantly inhibited by the intervention of NA.
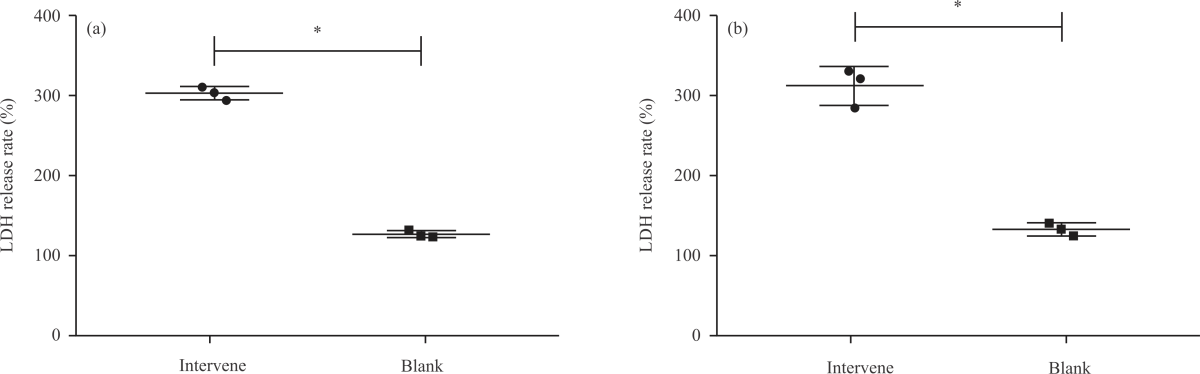
Impact of NA on LDH release rate in PD cell models: Then, we detected the LDH release rate of SH-SY5Y-MPP+ (Fig. 3a) and PC-12-MPP+ (Fig. 3b) cells. It was found that the LDH release rates of SH-SY5Y-MPP+ and PC-12-MPP+ in the model group were (292.75±11.26%, Fig. 3a) and (308.32±46.75%, Fig. 3b) respectively, which were the highest among the five groups (p<0.05), while those of the control group were the lowest [(128.00±7.17%), (135.71±6.86)%] (p<0.05). Among the three groups of cells treated with NA, the LDH release rate decreased significantly and the order from high to low was low-dose group, medium-dose group and high-dose group (p<0.05). It can be seen that NA can also effectively inhibit the LDH release rate of the PD cell model.
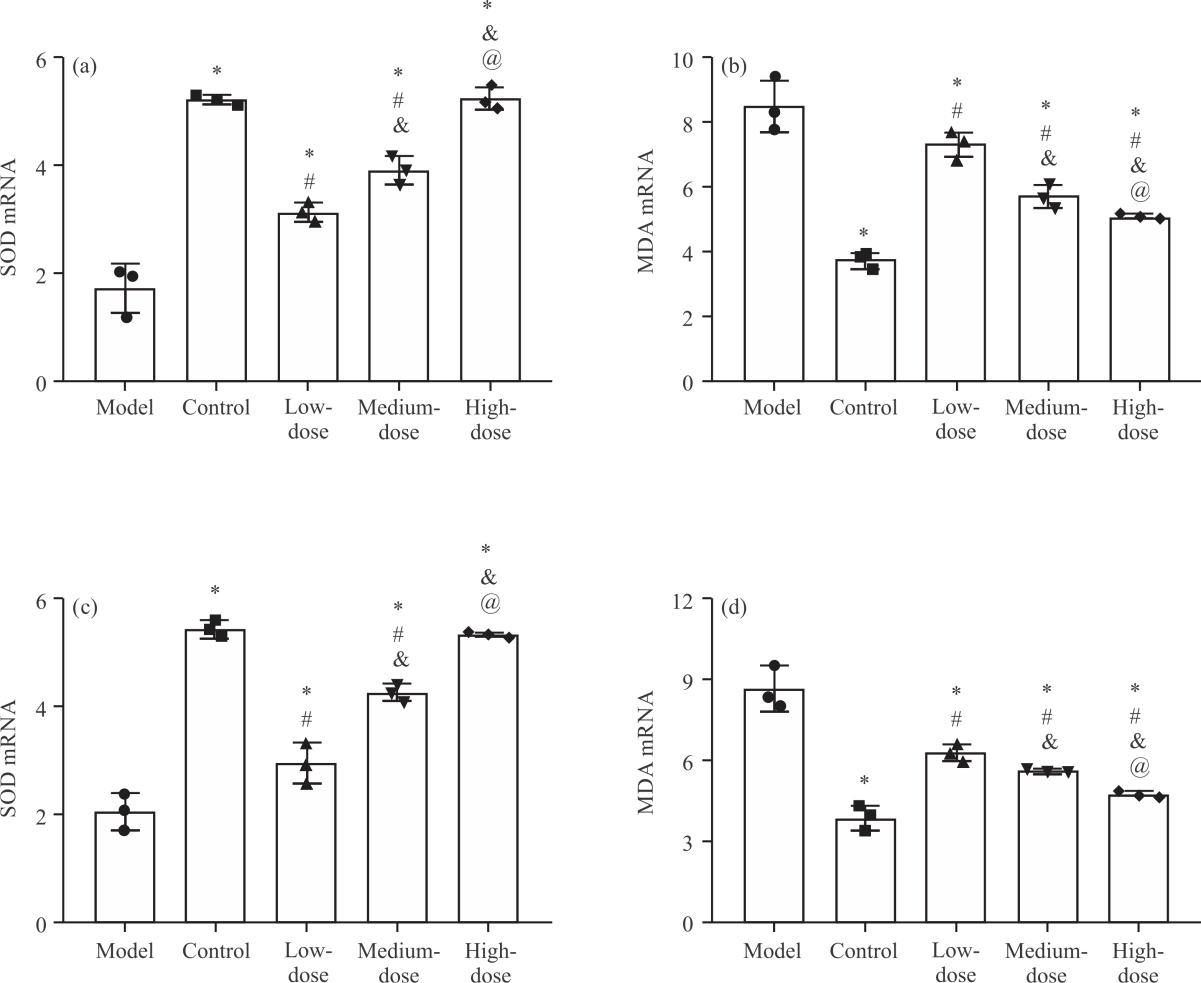 |
| Fig. 2(a-d): | Impact of NA on oxidative stress in PD cell models, (a) SOD mRNA level in SH-SY5Y-MPP+, (b) MDA mRNA level in SH-SY5Y-MPP+, (c) SOD mRNA level in PC-12-MPP+ and (d) MDA mRNA level in PC-12-MPP+ *Model group, p<0.05, #Control group, p<0.05, &Low-dose group, p<0.05, @Medium-dose group, p<0.05 and X-axis: Groups |
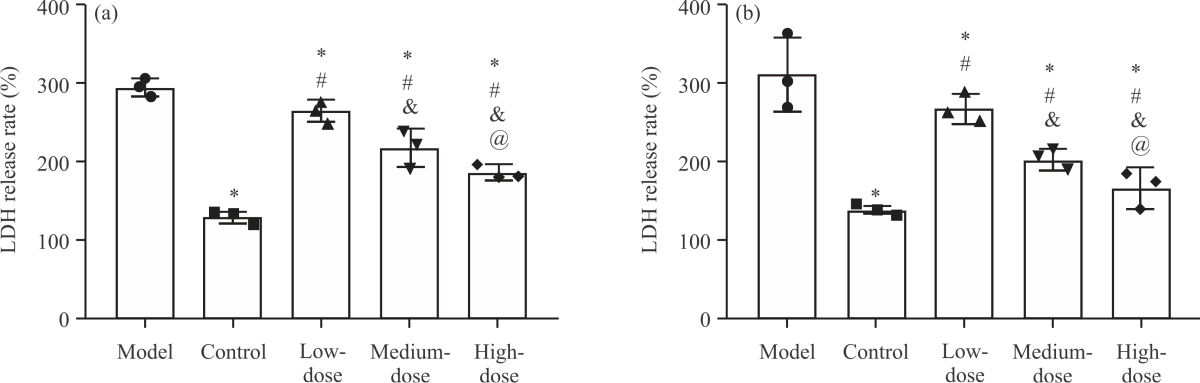 |
| Fig. 3(a-b): | Impact of NA on LDH release rate in PD cell models, (a) LDH release rate of SH-SY5Y-MPP+ and (b) LDH release rate of PC-12-MPP+ *Model group, p<0.05, #Control group, p<0.05, &Low-dose group, p<0.05, @Medium-dose group, p<0.05 and X-axis: Groups |
Impact of NA on Ras/MEK/ERK axis in PD cell models: Western blot analysis showed that the protein levels of Ras, p-MEK1/2/MEK1/2 and p-ERK1/2/ERK1/2 were the lowest in SH-SY5Y-MPP+ (Fig. 4a-b) and PC-12-MPP+ (Fig. 4c-d) cells in the model group (p<0.05), indicating that the Ras/MEK/ERK signalling pathway was inhibited in the PD cell model.
However, the protein expression of Ras, p-MEK1/2/MEK1/2 and p-ERK1/2/ERK1/2 in the low- and medium-dose groups was higher than that in the model group and there was no difference between the high-dose group and the control group (p>0.05), suggesting that NA can activate the Ras/MEK/ERK signal pathway in the PD cell model.
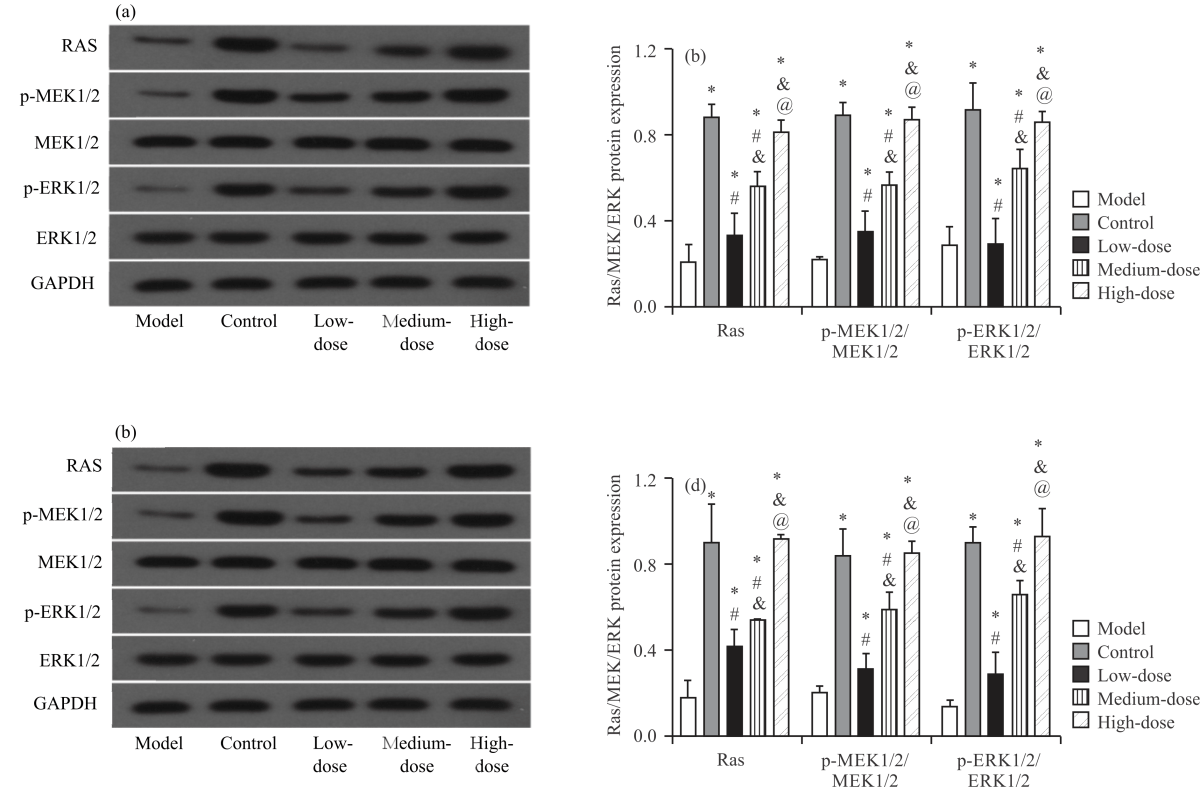 |
| Fig. 4(a-d): | Impact of NA on Ras/MEK/ERK axis in PD cell models, (a) Western blot diagram of SH-SY5Y-MPP+, (b) Ras/MEK/ERK pathway protein expression in SH-SY5Y-MPP+, (c) Western blot diagram of PC-12-MPP+ and (d) Ras/MEK/ERK pathway protein expression in PC-12-MPP+ *Model group, p<0.05, #Control group, p<0.05, &Low-dose group, p<0.05, @Medium-dose group, p<0.05 and X-axis: Protein expression groups |
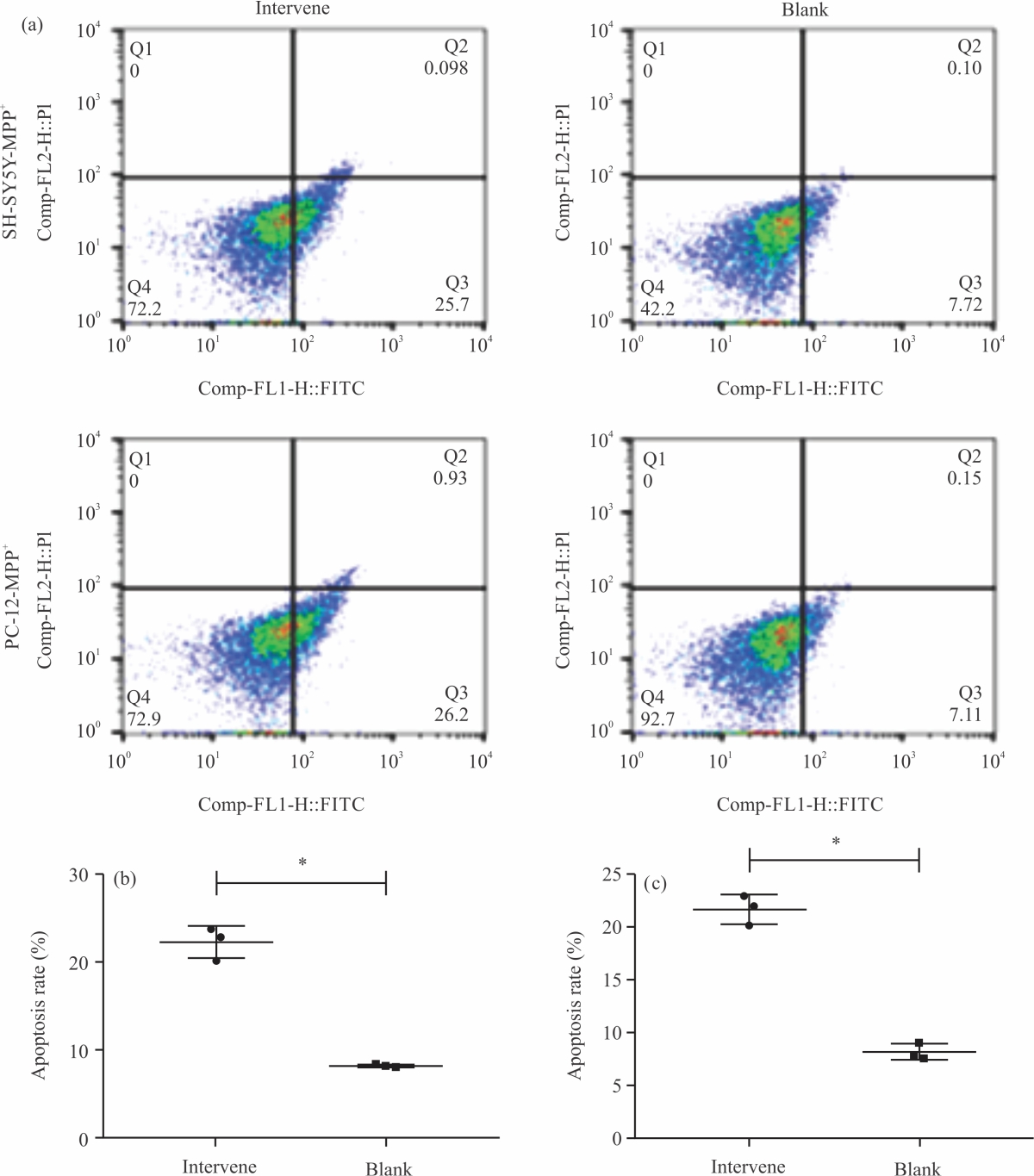
Impact of NA on the activity of PD modelled cells: The results of flow cytometry (Fig. 5a) showed that the apoptosis rates of SH-SY5Y-MPP+ and PC-12-MPP+ cells in the model group were (23.76±0.76%, Fig. 5b) and (19.56±2.08%, Fig. 5c), respectively, which were the highest among the 5 groups (p<0.05), While the lowest apoptosis rates of SH-SY5Y-MPP+ and PC-12-MPP+ cells were determined in the control group (p<0.05), which were 8.19±0.36 and 7.98±0.71%, respectively. Among the three groups intervened by NA, the apoptosis rate was the highest in the low-dose group, followed in descending order by medium- and high-dose groups (p<0.05). The above data indicate that NA can effectively alleviate the apoptosis of PD model cells.
Impact of Ras/MEK/ERK pathway on inflammatory responses in PD cell models: To further confirm the effect of the Ras/MEK/ERK signalling pathway on the PD cell model, we measured the expression of IFs in the intervention group treated with U0126. The results showed that the mRNA levels of IFs IL-6, IL-8 and TNF-α in SH-SY5Y-MPP+ (Fig. 6a-c) and PC-12-MPP+ (Fig. 6d-f) cells in the intervention group were significantly higher than those in the blank group (p<0.05), indicating that inhibiting Ras\/MEK/ERK signalling pathway can activate inflammatory responses in the PD cell model.
Impact of Ras/MEK/ERK on OS in PD cell models: Then, the OSR of cells under the intervention of U0126 was also detected. The results showed that the SOD mRNA levels of SH-SY5Y-MPP+ (Fig. 7a-b) and PC-12-MPP+ (Fig. 7c-d) cells in the intervention group were (2.00±0.24) and (1.95±0.23), respectively, lower than those in the control group (p<0.05), While higher MDA mRNA levels were determined in the intervention group (8.53±0.66), (8.84±0.36) compared with the control group (p<0.05). It is suggested that inhibition of the Ras/MEK/ERK signal pathway can also aggravate oxidative stress in the PD cell model.
Impact of Ras/MEK/ERK on LDH release rate in PD cell models: Similarly, the LDH release results showed that in the intervention group, the LDH release rate of SH-SY5Y-MPP+ was (303.37±8.17)% (Fig. 8a) and that of PC-12-MPP+ was (312.35±24.16)% (Fig. 8b), both of which were higher compared with the blank group (p<0.05).
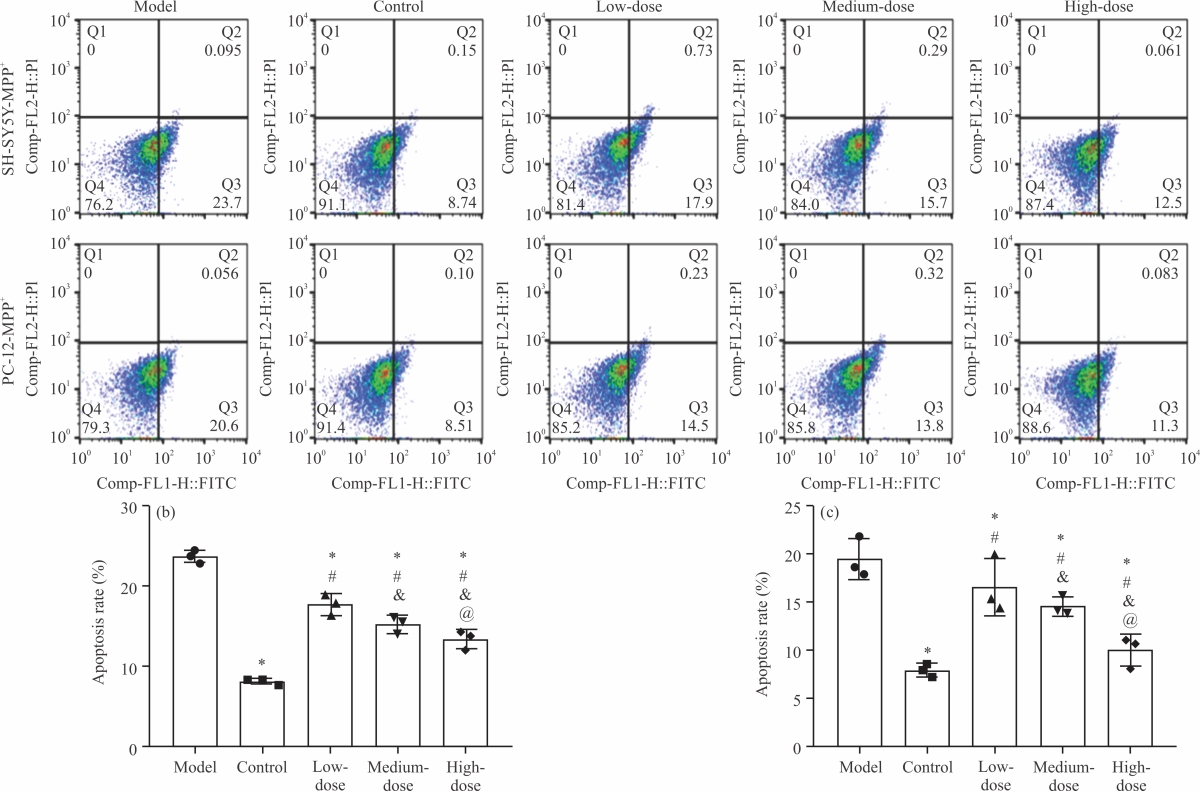 |
| Fig. 5(a-c): | Impact of NA on the activity of PD modelled cells, (a) Flow cytometry, (b) SH-SY5Y-MPP+ apoptosis rate and (c) PC-12-MPP+ apoptosis rate *Model group, p<0.05, #Control group, p<0.05, &Low-dose group, p<0.05, @Medium-dose group, p<0.05 and X-axis: Groups |
 |
| Fig. 6(a-f): | Impact of RAS/MEK/ERK pathway on inflammatory responses in PD cell models, (a) IL-6 mRNA level in SH-SY5Y-MPP+, (b) IL-8 mRNA level in H-SY5Y-MPP+, (c) Comparison of TNF-α mRNA levels in H-SY5Y-MPP+, (d) IL-6 mRNA level in PC-12-MPP+, (e) IL-8 mRNA level in PC-12-MPP+ and (f) Comparison of TNF-α mRNA levels in PC-12-MPP+ *Comparison between the two groups was p<0.05 and X-axis: Groups |
 |
| Fig. 7(a-d): | Impacts of RAS/MEK/ERK on oxidative stress in PD cell models, (a) SOD mRNA level in SH-SY5Y-MPP+, (b) MDA mRNA level in SH-SY5Y-MPP+, (c) SOD mRNA level in PC-12-MPP+ and (d) MDA mRNA level in PC-12-MPP+ *Comparison between the two groups was p<0.05 and X-axis: Groups |
 |
| Fig. 8(a-b): | Impact of Ras/MEK/ERK on LDH release rate in PD cell models, (a) LDH release rate of SH-SY5Y-MPP+ and (b) LDH release rate of PC-12-MPP+ *Comparison between the two groups was p<0.05 and X-axis: Groups |
 |
| Fig. 9(a-c): | Impact of Ras/MEK/ERK on the activity of PD modelled cells, (a) Flow cytometry, (b) SH-SY5Y-MPP+ apoptosis rate and (c) PC-12-MPP+ apoptosis rate *Comparison between the two groups was p<0.05 and X-axis: Groups |
The data suggest that the Ras/MEK/ERK signalling pathway is also closely related to the LDH release rate of the PD cell model.
Impact of Ras/MEK/ERK on the activity of PD modelled cells: Finally, the results of flow cytometry (Fig. 9a) showed that the apoptosis rate of SH-SY5Y-MPP+ in the intervention group was (22.23±1.87)%, which was higher than that in the blank group (p<0.05, Fig. 9b). Besides, a higher apoptosis rate of PC-12-MPP+ cells was also determined in the intervention group (21.64±1.39)% compared with the blank group (p<0.05, Fig. 9c). It can be seen that inhibition of the Ras/MEK/ERK signal pathway also significantly increases the apoptosis rate of the PD cell model.
Rescue experiment: To confirm that NA affects the PD cell model through the Ras/MEK/ERK signal pathway, we intervened SH-SY5Y-MPP+ and PC-12-MPP+ cells with NA and U0126 at the same time and detected the IFs (Fig. 10a-b), oxidative stress (Fig. 10c-d), LDH release rate (Fig. 10e) and apoptosis rate (Fig. 10f-g) again.
 |
| Fig. 10(a-g): | Rescue experiment, (a) SH-SY5Y-MPP+ inflammatory factor level, (b) PC-12-MPP+ inflammatory factor level, (c) SH-SY5Y-MPP+ oxidative stress response, (d) PC-12-MPP+ oxidative stress response, (e) LDH release rate, (f) Flow cytometry and (g) Apoptosis rate *Group A, p<0.05, #Group C, p<0.05 and X-axis: Expression types |
The results showed that groups A and C were not significantly different in levels of Ifs, OSR, LDH release rate and apoptosis rate (p>0.05), higher than those in group B (p<0.05). It indicates that the effect of NA on SH-SY5Y-MPP+ and PC-12-MPP+ can be completely reversed by U0126, which confirmed the relationship between the two cells.
DISCUSSION
In this experiment, researchers used MPP+ to induce SH-SY5Y and PC-12 to establish PD cell models, which is also the most commonly used PD cell model building scheme in clinical practice and has been proved to be of value in several studies19,20. Researchers detected the IFs and OSR in PD cell models and found elevated IL-6, IL-8, TNF-α and MDA while decreased SOD in the model group were all increased, which was consistent with the previous research on the PD cell model, that is, there were obvious OSR and inflammatory injury in PD cells21. In the three groups of cells intervened by NA, the IFs and OS were inhibited, indicating that NA could effectively intervene in the process of PD injury, which was consistent with our expectation. And in this study, it was explored the influence of NA on PD cell models, which has important reference significance for the future clinical application of NA. NA, as a key substance for nerve growth and structural stability, has shown excellent therapeutic effects in various nervous system diseases22. As Altinoz et al.23 pointed out, NA renders stable benefits for the immunity and metabolism of senile dementia patients. Lewkowicz et al.24 suggested that NA can improve oligodendrocyte activity. At present, the incidence of PD is increasing year by year, which brings increasing challenges to the clinic25. For PD, the application of NA may become a new direction for future treatment that can guarantee the normal living standard of patients.
In our previous research, also found that NA can alleviate the oxidative damage and inflammatory reaction of PD mouse model26, which can also testify to our experimental results and further confirm the value of NA in nervous system diseases. In addition, LDH, as an extremely important enzyme in the process of energy metabolism of the human body, is found in almost all tissues. When tissue necrosis occurs, LDH can be released into the blood in large quantities, so it is often applied to the diagnosis of various tissue injury diseases. In PD, the LDH release rate also shows an obvious trend of increase27. In our study, the release rate of LDH in PD modelled cells decreased remarkably after NA intervention, which once again emphasized that NA could repair tissue damage and can alleviate PD. Looking up relevant studies, we conclude that NA has the following effects28,29: (1) It can repair and dredge nerve fibres, improve cranial nerve activity and restore nerve terminal viability, Moreover, it is capable of promoting nerve cell growth and development, preventing neurofibroatrophy and facilitating the recovery of function after brain injury, (2) It can suppress the degenerative changes of nerve fibres and nerve cells in the brain and prevent the collapse, atrophy and hardening of brain tissue, (3) It can reduce the accumulation of lipofuscin in brain cells and prevent brain cells from ageing while improving brain blood circulation, ensuring an adequate supply of blood oxygen in brain cells and promoting metabolism, (4) It can provide nutrition for cranial nerves, enhance brain cell energy, promote cell proliferation and differentiation and improve brain functions such as cognition, discrimination, learning, thinking and memory. As for the pathogenesis of PD, it is precisely due to the degeneration and death of neurons caused by dopamine in the substantia nigra of the midbrain, obvious inflammatory damage in the cytoplasm of neurons, as well as dopamine ability system disorder of different degrees, resulting in a series of endocrine disorders30. NA can not only fundamentally alleviate the pathological changes of PD, but also further regulate the stability of multiple internal functions, which is especially suitable for the treatment of PD. The results of this experiment also preliminarily confirmed this view.
Subsequently, to further understand NA’s influence on PD, we detected the cell activity under the intervention of NA. The results identified enhanced apoptosis in the model group while decreased apoptosis in cells intervened by NA, suggesting that NA can effectively inhibit the death of PD cells. Furthermore, we found an activated Ras/MEK/ERK axis under NA intervention. The role of the Ras/MEK/ERK axis as a promoter of cell activity has been confirmed in many diseases including stroke and this pathway is also vital in mediating cell activity31. Moreover, inhibition of the Ras/MEK/ERK pathway increased the OSR, IFs, LDH release rate and apoptosis rate of PD model cells, which can also fully verify its important role in PD. Finally, through the rescue experiment, we found that the influence of NA on PD model cells could be completely reversed by inhibiting the Ras/MEK/ERK axis. The above experimental results confirmed our conjecture that NA influenced PD model cells by regulating the Ras/MEK/ERK axis, which can lay a reliable foundation for the future clinical application of NA.
However, this experiment also has many shortcomings to be addressed. For example, the absence of in vivo experiments failed to clarify how NA affects PD in vivo. Furthermore, NA may take a part in PD cell models through some other pathways rather than the one we studied here, which needs further exploration. Because of the above limitations, we will carry out a more in-depth and comprehensive experimental analysis as soon as possible to provide a more reliable reference for the clinical application of NA.
CONCLUSION
In PD cells, there were obvious inflammatory responses and oxidative stress responses and the LDS release rate and apoptosis rate of the cells increased. NA inhibits the apoptosis of PD cells via activating the Ras/MEK/ERK axis and alleviates the OS injury and inflammatory injury of cells, thus achieving the goal of treating PD, which may be a breakthrough in future PD treatment, to solve the current situation of poor clinical effect on PD treatment.
SIGNIFICANCE STATEMENT
This study discovered the role of Neuronic acid in Parkinson’s disease that can be beneficial for these patients. This study will help the researchers to uncover the critical areas of Neuronic acid that affect Parkinson’s disease therapy that many researchers were not able to explore. Thus a new theory on Neuronic acid regulating the Parkinson’s cell apoptosis and inflammatory damage of cells may be arrived at.
REFERENCES
- Francula-Zaninovic, S. and I.A. Nola, 2018. Management of measurable variable cardiovascular disease' risk factors. Curr. Cardiol. Rev., 14: 153-163.
CrossRefDirect Link - Dorsey, E.R., T. Sherer, M.S. Okun and B.R. Bloem, 2018. The emerging evidence of the Parkinson pandemic. J. Parkinson's Dis., 8: S3-S8.
CrossRefDirect Link - Vignatelli, L., C. Zenesini, L.M.B. Belotti, E. Baldin and G. Bonavina et al., 2021. Risk of hospitalization and death for COVID-19 in people with Parkinson's disease or Parkinsonism. Mov. Disord., 36: 1-10.
CrossRefDirect Link - Puschmann, A., 2017. New genes causing hereditary Parkinson’s disease or Parkinsonism. Curr. Neurol. Neurosci. Rep., Vol. 17.
CrossRefDirect Link - Darweesh, S.K.L., K.G. Raphael, P. Brundin, H. Matthews, R.K. Wyse, H. Chen and B.R. Bloem, 2018. Parkinson matters. J. Parkinson's Dis., 8: 495-498.
CrossRefDirect Link - Timmer, M.H.M., M.H.C.T. van Beek, B.R. Bloem and R.A.J. Esselink, 2017. What a neurologist should know about depression in Parkinson’s disease. Pract. Neurol., 17: 359-368.
CrossRefDirect Link - Lin, G., L. Wang, P.C. Marcogliese and H.J. Bellen, 2019. Sphingolipids in the pathogenesis of Parkinson’s disease and parkinsonism. Trends Endocrinol. Metab., 30: 106-117.
CrossRefDirect Link - Habets, J.G.V., M. Heijmans, M.L. Kuijf, M.L.F. Janssen, Y. Temel and P.L. Kubben, 2018. An update on adaptive deep brain stimulation in Parkinson's disease. Mov. Disord., 33: 1834-1843.
CrossRefDirect Link - Fan, Y., H.M. Meng, G.R. Hu and F.L. Li, 2018. Biosynthesis of nervonic acid and perspectives for its production by microalgae and other microorganisms. Appl. Microbiol. Biotechnol., 102: 3027-3035.
CrossRefDirect Link - Kageyama, Y., T. Kasahara, T. Nakamura, K. Hattori and Y. Deguchi et al., 2018. Plasma nervonic acid is a potential biomarker for major depressive disorder: A pilot study. Int. J. Neuropsychopharmacol., 21: 207-215.
CrossRefDirect Link - Li, Q., J. Chen, X. Yu and J.M. Gao, 2019. A mini review of nervonic acid: Source, production, and biological functions. Food Chem., Vol. 301.
CrossRefDirect Link - Ma, Q., T. Sun, S. Li, J. Wen and L. Zhu et al., 2020. The Acer truncatum genome provides insights into nervonic acid biosynthesis. Plant J., 104: 662-678.
CrossRefDirect Link - Kageyama, Y., Y. Deguchi, K. Hattori, S. Yoshida, Y.I. Goto, K. Inoue and T. Kato, 2021. Nervonic acid level in cerebrospinal fluid is a candidate biomarker for depressive and manic symptoms: A pilot study. Brain Behav., Vol. 11.
CrossRefDirect Link - Umemoto, H., S. Yasugi, S. Tsuda, M. Yoda, T. Ishiguro, N. Kaba and T. Itoh, 2021. Protective effect of nervonic acid against 6-hydroxydopamine-induced oxidative stress in PC-12 cells. J. Oleo Sci., 70: 95-102.
CrossRefDirect Link - Ntoumani, E., B. Strandvik and K.G. Sabel, 2013. Nervonic acid is much lower in donor milk than in milk from mothers delivering premature infants-of neglected importance? Prostaglandins Leukotrienes Essent. Fatty Acids, 89: 241-244.
CrossRefDirect Link - Kim, E.K. and E.J. Choi, 2015. Compromised MAPK signaling in human diseases: An update. Arch. Toxicol., 89: 867-882.
CrossRefDirect Link - Bai, L., F. Yan, R. Deng, R. Gu, X. Zhang and J. Bai, 2021. Thioredoxin-1 rescues MPP+/MPTP-induced ferroptosis by increasing glutathione peroxidase 4. Mol. Neurobiol., 58: 3187-3197.
CrossRefDirect Link - Vozella, V., A. Basit, A. Misto and D. Piomelli, 2017. Age-dependent changes in nervonic acid-containing sphingolipids in mouse hippocampus. Biochim. Biophys. Acta, Mol. Cell. Biol. Lipids, 1862: 1502-1511.
CrossRefDirect Link - Zhao, J., L. Geng, Y. Chen and C. Wu, 2020. SNHG1 promotes MPP+-induced cytotoxicity by regulating PTEN/AKT/mTOR signaling pathway in SH-SY5Y cells via sponging miR-153-3p. Biol. Res., Vol. 53.
CrossRefDirect Link - Zhang, G.F., Y. Zhang and G. Zhao, 2015. Crocin protects PC12 cells against MPP+-induced injury through inhibition of mitochondrial dysfunction and ER stress. Neurochem. Int., 89: 101-110.
CrossRefDirect Link - Trist, B.G., D.J. Hare and K.L. Double, 2019. Oxidative stress in the aging substantia nigra and the etiology of Parkinson's disease. Aging Cell, Vol. 18.
CrossRefDirect Link - Voshavar, C., M. Shah, L. Xu and A.K. Dutta, 2015. Assessment of protective role of multifunctional dopamine agonist D-512 against oxidative stress produced by depletion of glutathione in PC12 cells: Implication in neuroprotective therapy for Parkinson’s disease. Neurotox. Res., 28: 302-318.
CrossRefDirect Link - Altinoz, M.A. and A. Ozpinar, 2019. PPAR-δ and erucic acid in multiple sclerosis and Alzheimer's disease. Likely benefits in terms of immunity and metabolism. Int. Immunopharmacol., 69: 245-256.
CrossRefDirect Link - Lewkowicz, N., P. Piątek, M. Namiecińska, M. Domowicz and R. Bonikowski et al., 2019. Naturally occurring nervonic acid ester improves myelin synthesis by human oligodendrocytes. Cells, Vol. 8.
CrossRefDirect Link - Nassir, C.M.N.C.M., T. Damodaran, S.R. Yusof, A. Norazit and G. Chilla et al., 2021. Aberrant neurogliovascular unit dynamics in cerebral small vessel disease: A rheological clue to vascular Parkinsonism. Pharmaceutics, Vol. 13.
CrossRefDirect Link - Hu, D., Y. Cui and J. Zhang, 2021. Nervonic acid amends motor disorder in a mouse model of Parkinson’s disease. Transl. Neurosci., 12: 237-246.
CrossRefDirect Link - Song, Y., Y. Liu and X. Chen, 2018. MiR-212 attenuates MPP+-induced neuronal damage by targeting KLF4 in SH-SY5Y cells. Yonsei Med. J., 59: 416-424.
CrossRefDirect Link - Xu, C.Q., H. Liu, S.S. Zhou, D.X. Zhang and W. Zhao et al., 2019. Genome sequence of Malania oleifera, a tree with great value for nervonic acid production. GigaScience, Vol. 8.
CrossRefDirect Link - Konstantynowicz-Nowicka, K., K. Berk, A. Chabowski, I. Kasacka, P. Bielawiec, B. Łukaszuk and E. Harasim-Symbor, 2019. High-fat feeding in time-dependent manner affects metabolic routes leading to nervonic acid synthesis in NAFLD. Int. J. Mol. Sci., Vol. 20.
CrossRefDirect Link - Li, B., L. Lv and W. Li, 2020. 1,25-Dihydroxy vitamin D3 inhibits the Ras-MEK-ERK pathway and regulates proliferation and apoptosis of papillary thyroid carcinoma. Steroids, Vol. 159.
CrossRefDirect Link - Yao, W., Z. Lin, P. Shi, B. Chen and G. Wang et al., 2019. Delicaflavone induces ROS-mediated apoptosis and inhibits PI3K/AKT/mTOR and Ras/MEK/Erk signaling pathways in colorectal cancer cells. Biochem. Pharmacol., Vol. 171.
CrossRefDirect Link