
Jianming Dai
Department of Orthopedics, The people’s Hospital of Zhongjiang, Deyang Sichuan 618100, China
Zaifeng Wu
Department of Pathology, The First Hospital of Qiqihar Affiliated Qiqihar Hospital, Southern Medical University, Qiqihar, Heilongjiang 161000, China
LiveDNA: 86.37375
International Journal of Pharmacology
Year: 2022 | Volume: 18 | Issue: 6 | Page No.: 1189-1198







ABSTRACT
Background and Objective: Mitochondrial improvement is the central player of neuroprotection following spinal cord injury (SCI). The present study evaluated the neuroprotective and anti-inflammatory effects of a new mitochondrial-acting drug, ginsenoside compound Mc1 (GCM), in rats with traumatic SCI, by focusing on the role of mitochondrial ATP-sensitive potassium (ATP-KCh) channels and mitochondrial biogenesis. Materials and Methods: The SCI was modelled in Sprague Dawley rats (n = 60) through laminectomy and placing an extradural aneurysm clip at the T10 vertebral level. The GCM at the concentrations of 20 and 40 μM was intraperitoneally administered to rats at 1, 6 and 12 hrs after laminectomy. Locomotor activity, spinal cord histology, mitochondrial function and biogenesis and the levels of pro-inflammatory cytokines were quantified. Results: Administration of GCM significantly reduced SCI-induced neurological locomotor deficits and histopathological findings in a dose-dependent manner, as compared with the untreated control group (p<0.01). The GCM (40 μM) significantly reversed the SCI-induced changes in mitochondrial membrane depolarization, mitochondrial reactive oxygen species (mitoROS), ATP levels, superoxide dismutase (mnSOD) and pro-inflammatory cytokines TNF-α, IL-1β, IL-8 (p<0.01). As well, the expression of mitochondrial biogenesis proteins PGC-1α, NRF1 and TFAM was upregulated following GCM 40 μM treatment. Importantly, the inhibition of mitochondrial ATP-KCh through hydroxydecanoate significantly eliminated the neuroprotective, anti-inflammatory and mitochondrial impacts of GCM. Conclusion: The GCM post-treatment had a significant neuroprotective effect on SCI which was mediated through increasing mitochondrial ATP-KCh activity and subsequent improvement of mitochondrial biogenesis and function and reduction of inflammatory responses.
PDF Abstract XML References Citation
Copyright: © 2022. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Jianming Dai and Zaifeng Wu, 2022. Mitochondrial Potassium Channel-Dependent Anti-Inflammatory Effects of Ginsenoside Mc1 in Rat Spinal Cord Injury. International Journal of Pharmacology, 18: 1189-1198.
DOI: 10.3923/ijp.2022.1189.1198
URL: https://scialert.net/abstract/?doi=ijp.2022.1189.1198
DOI: 10.3923/ijp.2022.1189.1198
URL: https://scialert.net/abstract/?doi=ijp.2022.1189.1198
INTRODUCTION
Injuries to the spinal cord caused by a variety of factors, result in diverse complications including sensory and motor dysfunction, neurological reflex problems and sphincter disorders the organs that lower the level of injury1. Facing such problems and how to treat and manage the patients with them is considered as one of the important clinical challenges. Due to the industrialization of daily life, the incidence of traumatic spinal cord injuries has increased due to falls from heights or traffic accidents, which has increased the inability to live a normal life as well as deaths due to spinal cord injury (SCI)1,2. Therefore, exploring pathophysiological mechanisms of SCI and discovering more effective treatments is clinically valuable.
The pathophysiology of SCI is numerous and includes tissue destruction, inflammatory and oxidative reactions, apoptosis and necrosis3. Inflammatory responses play a very critical role in exacerbating SCI. According to the results of previous studies, the occurrence of oxidative stress following traumatic SCI provides the conditions for the production of proinflammatory cytokines such as Tumour Necrosis Factor-alpha (TNF-α) and interleukins and thus exacerbates the injury and its complications4. Mitochondria, where O2 is consumed and where important biological reactions and ATP production take place in neurons, become dysfunctional due to SCI3,4. As a result, the expression of proteins involved in mitochondrial biogenesis and its dynamics is severely reduced. These changes intensify the production of inflammatory cytokines and oxidative responses5. Studies have shown that ATP-dependent potassium channels (ATP-KCh) located in mitochondrial membranes play a central role in promoting normal mitochondrial function and enhancing its biogenesis6. Opening of these channels has significant neuroprotective effects in neurological diseases7 and therefore, targeting mitochondrial ATP-KCh can reduce the consequences of SCI and can yield promising results.
The use of various promising research strategies in the development of effective and safe therapeutic interventions for serious neurological disorders including SCI is still an unmet need. Ginseng-derived compounds have shown numerous anti-inflammatory, anti-apoptotic, antioxidant and mitochondrial improving effects in neurodegenerative diseases8,9. Ginsenoside compound Mc1 (GMC) is much more pharmacologically active than other compounds due to its non-glycosylated structure10. Recent limited studies have reported that this newly constructed compound may play a significant role in reducing the oxidative and apoptotic responses of heart and liver cells by amplifying the AMPK signal pathway and reducing endoplasmic reticulum stress through the JAK2/STAT3 pathway11,12. Due to the supportive effects of ginsenoside compounds on mitochondrial targeting pathways in neurodegenerative disorders13, GMC can be more useful in subsiding the complications of SCI. To our knowledge, there is no information about the neurological effects of GMC. Therefore, in this study, we aimed to investigate the protective effects of GMC on SCI complications by evaluating it’s anti-inflammatory and boosting effects on mitochondrial function/biogenesis and to explore the possible role of mitochondrial ATP-KCh.
MATERIALS AND METHODS
Study area: The present work was performed at the Department of Pathology, Affiliated Qiqihar Hospital, at Southern Medical University, China from August, 2020 to December, 2021.
Animals: Sixty male Sprague-Dawley rats with 3-4 months old and 250±20 g weights were kept in an animal room with the conditions of 23±2°C and a diurnal cycle of 12/12 hrs lightness/darkness. The animals had free access to food and water. The animal handling and experiments were performed based on the international ethics protocols and approved by the animal ethics committee of the university (approval number: CBETH2020).
Animal grouping: After 10 days acclimation period, the animals were divided randomly into six groups as follow: Group I: Control (intact animals), Group II: SCI (injury-model group), Group III: SCI+G20 (treatment group with GMC 20 μm), Group IV: SCI+G40 (treatment group with GMC 40 μm), Group V: SCI+HD (ATP-KCh inhibitor group) and Group VI: SCI+HD+G40 (treatment group with GMC 50 μm in the presence of ATP-KCh inhibitor). N number for each group was 10. The animals in the control group experienced laminectomy without SCI challenge. In the G20 and G40-receiving groups, the rats were treated with GCM at the dosages of 20 and 40 μm i.p., respectively, 1, 6 and 12 hrs after laminectomy and SCI induction. The drug was diluted in 1% DMSO (vehicle) and the non-treated SCI rats were given the same volumes of vehicle, i.p. For blockade of mitochondrial ATP-KCh, 100 μm hydroxydecanoate (HD) was intraperitoneally injected before administration of GCM at 40 μM. Alone administration of HD in SCI rats had no significant effect on neurological and histological findings and therefore the data of this group were excluded from the results section.
Induction of spinal cord injury: The induction of traumatic SCI in rats was accomplished as reported earlier1. Following 12 hrs fasting, the rats were anaesthetized with sodium pentobarbital (40 mg kg–1, i.p.). Body temperature was monitored and kept at 37±0.5°C throughout the surgery. The Rats were placed in the prone position and a complete single laminectomy at T10 level was performed through a 20 mm incision under a dissecting microscope and aseptic condition. After revealing the spinal cord, the SCI was induced by the extradural application of a temporary aneurysm clip with a constant closing force of 24 g on the spinal cord for 60 sec1. The injury was done without damaging the dura mater. After surgery and suturing the skin incision, the rats were maintained warm and allowed to be recovered fully from anaesthesia. Until the recovery of urinary activity following SCI induction, bladder massage was employed two times a day to ease urination.
Assessment of motor function: Locomotor activity of rats was recorded on the 7th day after laminectomy by employing the basso, beattie and bresnahan (BBB) open-field locomotor scores scale. The rats were assessed for BBB score on a scale ranging from 0, indicating complete paralysis of hind limbs, to 21, indicating almost normal locomotor function. Two blind observers to study groups scored each animal and the average values were considered as the final result to reduce the experiment bias.
Samples collection: Collection of tissue samples was done 7 days after interventions. Under pentobarbital-induced anaesthesia, the lesioned part of dorsal horns of spinal cord tissue from six rats in each group was collected and kept at -70°C for biochemical and molecular measurements. Also, five rats per group were perfused with paraformaldehyde. Thereafter, the spinal cord tissues were taken, fixed in 10% buffered formalin and sectioned for histological evaluation. The motor function test was recorded by an observer blind to the study.
Histological evaluation: The spinal cord tissue from 4 rats was fixed with 10% buffered formalin, then embedded in paraffin and cut in 5 μm-thick horizontal sections. After deparaffinization, the samples were stained with Haematoxylin/Eosin (H&E) to quantify the morphological assessment of tissue. Photomicrographs of an area containing the spinal cord lesions were captured under a light microscope (Dialux 22 Leitz, Milan Italy) at 40× magnification. The histopathologic changes of neuronal degeneration, necrosis and inflammatory infiltration were scored according to a 5-degree grading system to verify the severity of spinal cord damage. The scores of all the sections were averaged to give a final score for an individual rat.
Measurement of mitochondrial function indices: Damaged spinal sections (5 mm) were isolated and homogenized at 4°C in a lysate buffer solution consisting of protease and phosphatase inhibitors. The mitochondrial isolation kit (Biochain. Newark, CA, USA) was employed to isolate the spinal mitochondria. In addition, the Pierce BCA protein assaying kit (Thermo Fisher Scientific, Waltham, USA) was used to quantify the protein contents of mitochondrial and tissue homogenates. For detecting mitochondrial ROS levels, the resulting supernatant, which is the mitochondrial fraction, was incubated for 30 min at 37°C in a phosphate-buffered solution (PBS) containing 2 μmol DCFDA dye (Sigma-Aldrich, USA). Then, using a fluorimeter, the amount of excitation and its emission were measured at 480 and 530 nm, respectively. The obtained values were calibrated based on the protein concentration of the samples and reported as mitochondrial ROS levels. Another amount of mitochondrial supernatant was incubated in 2 mL of PBS containing 2 μL of JC-1 dye (Sigma-Aldrich, USA) for 30 min at 37°C in a dark environment. After washing with PBS, the fluorescence intensity of JC-1 in each sample at red and green wavelengths was measured by a fluorimeter and the red to green ratio was used as an indicator to assess the degree of mitochondrial membrane depolarization. The ATP content of the samples was also measured using an ATP assay kit based on the instructions provided (Sigma-Aldrich, USA). Finally, mitochondrial manganese superoxide dismutase (MnSOD) was measured from the supernatants using an ELISA kit, according to instructions of the kit (Sigma-Aldrich, USA). The relative absorbance was read at 450 nm spectrophotometrically.
Measurement of proinflammatory cytokines: The ipsilateral side of the spinal cord centred on the impact site was removed for ELISA measurements. The samples were homogenized in RIPA buffer solution and after centrifugation and preparation of their supernatant, the contents of proinflammatory cytokines including TNF-α, interleukin-8 (IL-8) and IL-1β were measured using specific ELISA kits according to kit instructions (MyBioSource, Inc., USA). The protein content of the samples was also determined by the Bradford technique. The concentration of cytokines in each sample was standardized and reported based on the protein concentration of the samples.
Statistical analysis: The data were reported as Mean±SD. After confirming the normal distribution of data, the differences between the groups were analyzed by one-way analysis of variance (ANOVA) and Tukey post hoc test. The minimum level of significance was considered at the alpha level of 0.05.
RESULTS
Effect of GMC on the locomotor activity: Neurological locomotor activity was assessed using a BBB scoring system in rats with SCI. A greater neurological deficit was observed in SCI-receiving rats in comparison to the control group, indicating the successful induction of SCI. After SCI induction, GMC-treated rats showed higher degrees of locomotor activity in comparison to the SCI group, but the effect of the higher
dose of GMC (p<0.001) was greater than that of the low dose (p<0.05) (Fig. 1a). Alone administration of HD to inhibit mitochondrial ATP-KCh in SCI rats has no significant influence on the locomotor activity as compared with the SCI group (data were not shown). However, inhibition of these channels by HD significantly abolished the protective effect of the drug on neurological findings as compared with SCI rats receiving GMC 40 μM (p<0.01). To confirm GMC-protective effects, spinal cord histopathological changes were quantified using the H&E staining method and the results showed that administration of GMC at both doses significantly reduced SCI-induced spinal cord pathology in a dose-dependent manner (Fig. 1b). Blocking of ATP-KCh considerably abolished the GMC effects on locomotor dysfunction and histopathology changes compared to the SCI+G40 group (p<0.05).
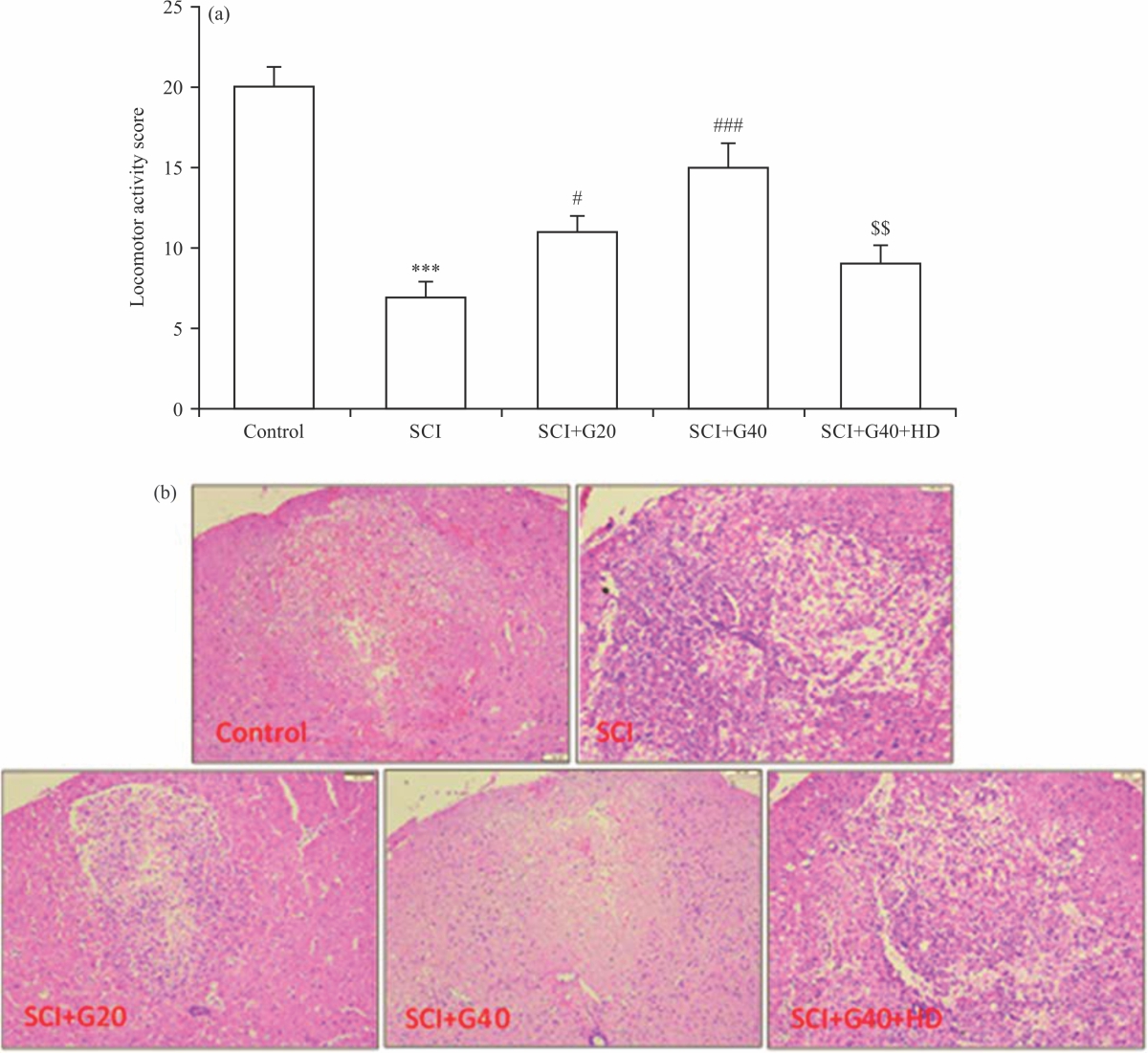 |
| Fig. 1(a-b): | Protective effects of GMC against spinal cord injury, (a) Locomotor activity score (n = 10) and (b) Histopathological changes (n = 4) ***p<0.001 vs. control group, #p<0.05, ###p<0.001 vs. SCI group, $$p<0.01 vs. SCI+G40 group, SCI: Spinal cord injury, G: Ginsenoside Mc1, GMC and HD: Hydroxydecanoate |
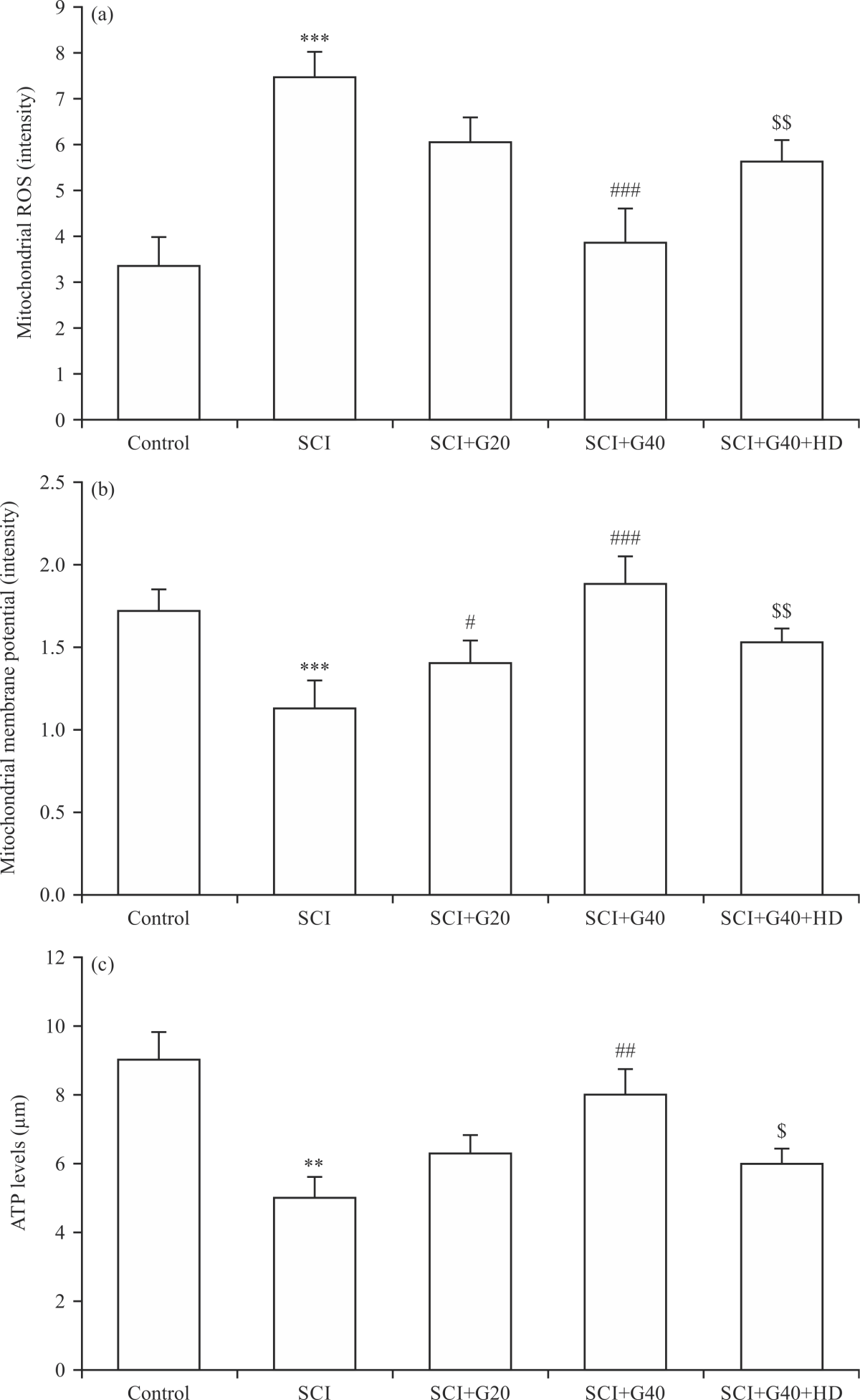 |
| Fig. 2(a-c): | Effects of GMC on mitochondrial function parameters in SCI model, (a) Mitochondrial ROS levels, (b) Mitochondrial membrane potential changes and (c) ATP levels (n = 6), ***p<0.001 vs. control group, #p<0.05, ###p<0.001 vs. SCI group, $$p<0.01 vs. SCI+G40 group, SCI: Spinal cord injury, G: Ginsenoside Mc1, GMC and HD: Hydroxydecanoate |
Effect of GMC on the mitochondrial activity: Mitochondrial activity was estimated through the assessment of mitochondrial ROS levels, mitochondrial membrane potential changes and ATP levels. The SCI significantly augmented the levels of mitochondrial ROS (Fig. 2a) and depolarization of mitochondrial membrane potential (reduced potential) (Fig. 2b) and reduced ATP levels as compared with the control group (p<0.01 and p<0.001)
(Fig. 2c). Nevertheless, GMC administration in rats after SCI significantly inverted SCI-induced changes in mitochondrial parameters, dose-dependently (p<0.05 and p<0.001). Furthermore, concomitant administration of HD significantly abolished the beneficial effects of GMC on the SCI-induced changes of mitochondrial ROS and membrane potential (p<0.01) as well ATP levels (p<0.05) (Fig. 2).
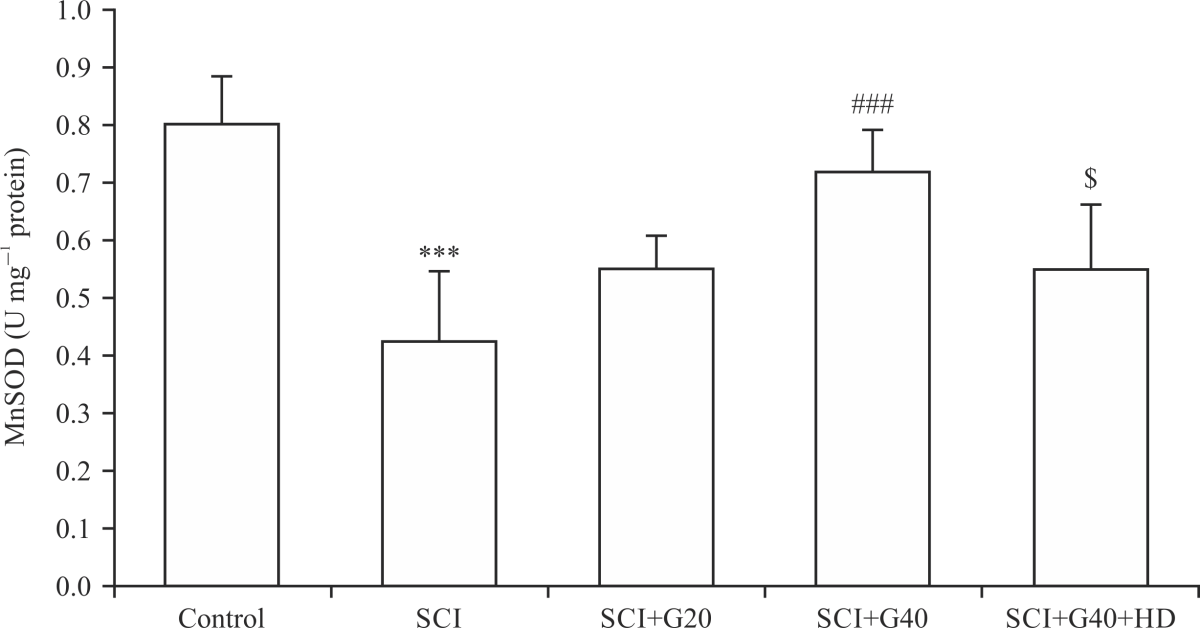 |
| Fig. 3: | Effect of GMC on mitochondrial MnSOD level in SCI model (n = 6), ***p<0.001 vs. control group, ###p<0.001 vs. SCI group, $p<0.05 vs. SCI+G40 group, SCI: Spinal cord injury, G: Ginsenoside Mc1, GMC and HD: hydroxydecanoate |
Effect of GMC on the MnSOD levels: The spinal cord levels of MnSOD, as an endogenous potent antioxidative enzyme, in the SCI group was significantly lower than that of the control group (p<0.001) (Fig. 3). The GMC at 40 μM but not 10 μM significantly elevated SCI-induced reduction of MnSOD (p<0.001). However, administration of HD to inhibit mitochondrial ATP-KCh significantly hindered the effect of the drug on the MnSOD level, as compared with the SCI+G40 group (p<0.05).
Effect of GMC on the expression of mitochondrial biogenesis proteins: Following in vivo SCI in rats, the protein expression of proteins regulating mitochondrial biogenesis (PGC-1α, NRF1 and TFAM) decreased significantly (p<0.001) (Fig. 4a-d). Administration of GMC at 20 μM was not capable to reduce SCI-induced downregulation of proteins. However, the expression levels of PGC-1α (p<0.001), NRF1 and TFAM (p<0.01) were significantly upregulated after administration of GMC at 40 μM, compared to the SCI group. Blocking of ATP-KCh by HD significantly prevented the effects of GMC at 40 μM on the expression of these proteins as compared with the SCI+G40 group (p<0.05) (Fig. 4).
Effect of GMC on the proinflammatory cytokines: As shown in Fig. 5, the production of proinflammatory cytokines TNF-α, IL-8 and IL-1β was significantly amplified after induction of SCI in rats compared with that in the control group (p<0.001). GMC at 20 μM did not affect the level of TNF-α, but it significantly reduced the production of IL-8 and IL-1β, in comparison to the SCI group (p<0.05) (Fig. 5a-c). Compared with the SCI group, a significant downregulation of the proinflammatory cytokines was observed in the spinal tissues of the rats after administration of GMC at 40 μM (p<0.01 and p<0.001). Furthermore, inhibition of mitochondrial ATP-KCh remarkably eliminated the anti-inflammatory actions of GMC 40 μM in the SCI setting (p<0.05 and p<0.001).
DISCUSSION
Post-treatment of rats with GMC significantly diminished the SCI-induced neurological motor scoring and histological findings in a dose-dependent manner. The GMC at 40 μm markedly improved mitochondrial function and upregulated the expression of PGC-α, NRF1 and TFAM biogenesis proteins. It also reduced the production of spinal inflammatory cytokines and inhibited mitochondrial oxidative stress and augmented ATP production. Blockade of mitochondrial ATP-KCh by HD significantly suppressed the neuroprotective, anti-inflammatory and mitochondrial boosting effects of GMC. These findings provided evidence that strengthening mitochondrial activity in spinal cord cells by GMC may be a promising strategy to prevent SCI.
Owing to extensive function, mitochondria in neural tissues have a special role in protecting them against fatal traumatic or exocytotoxic injury outcomes14. Under normal activity of mitochondria, cellular oxygen is used mostly for mitochondrial reduction-oxidation processes, reducing the likelihood of high mitochondrial ROS production and increasing the expression of endogenous antioxidative enzymes like MnSOD. Otherwise, mitochondria become dysfunctional with the sudden SCI and now they act as the main producer of ROS and free radicals in the cells, instead of ATP production14. Following amplification of mitochondrial oxidative stress and disruption of mitochondrial function, the production of pro-inflammatory cytokines is also accelerated15.
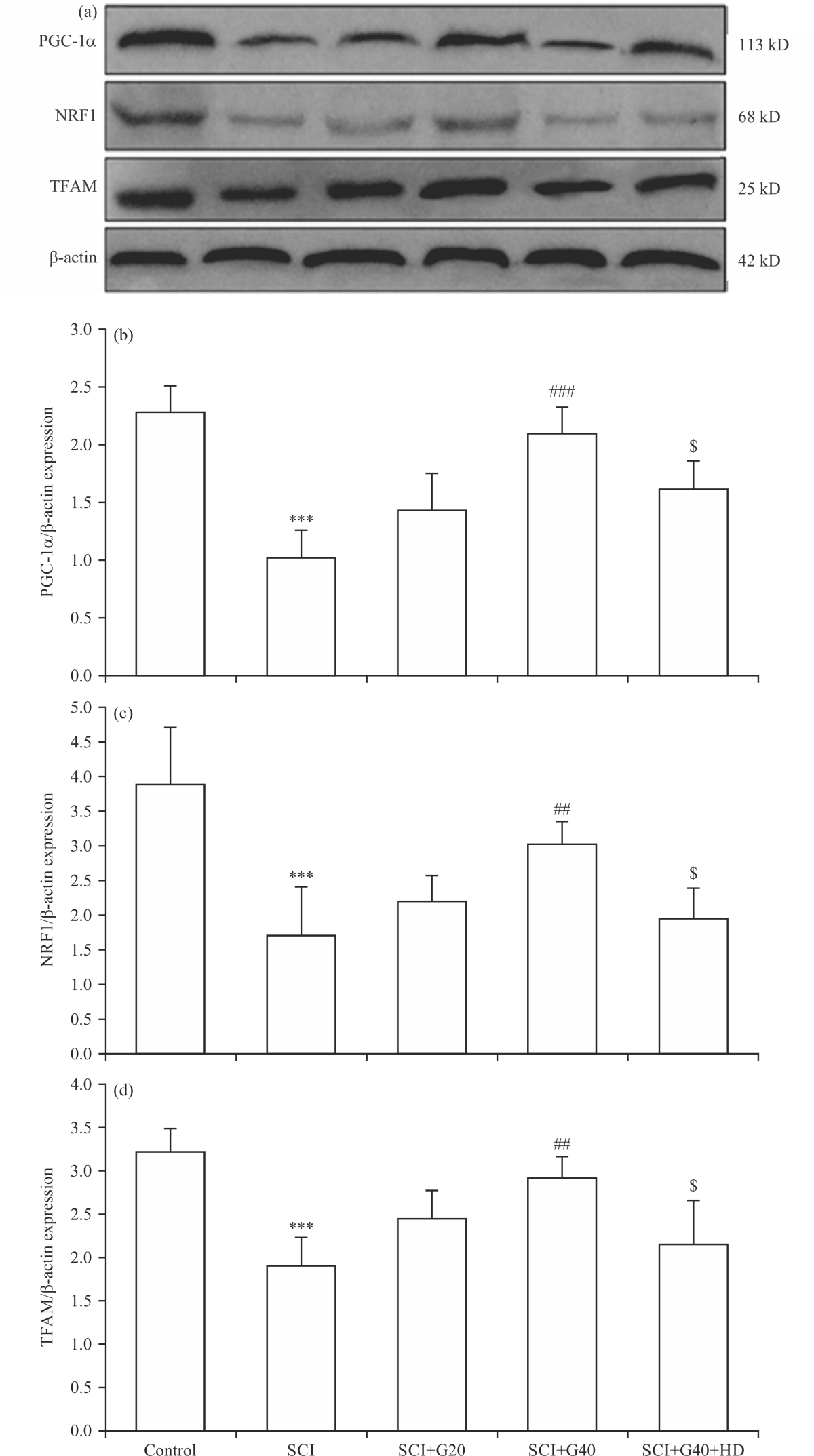 |
| Fig. 4(a-d): | Effects of GMC on the expression of proteins regulating mitochondrial biogenesis in SCI model, (a) Representative Immunoblots, (b) PGC-1α, (c) NRF1 and (d) TFAM (n = 4), ***p<0.001 vs. control group, #p<0.05, ##p<0.01, ###p<0.001 vs. SCI group, $p<0.05 vs. SCI+G40 group, SCI: Spinal cord injury, G: Ginsenoside Mc1, GMC and HD: Hydroxydecanoate |
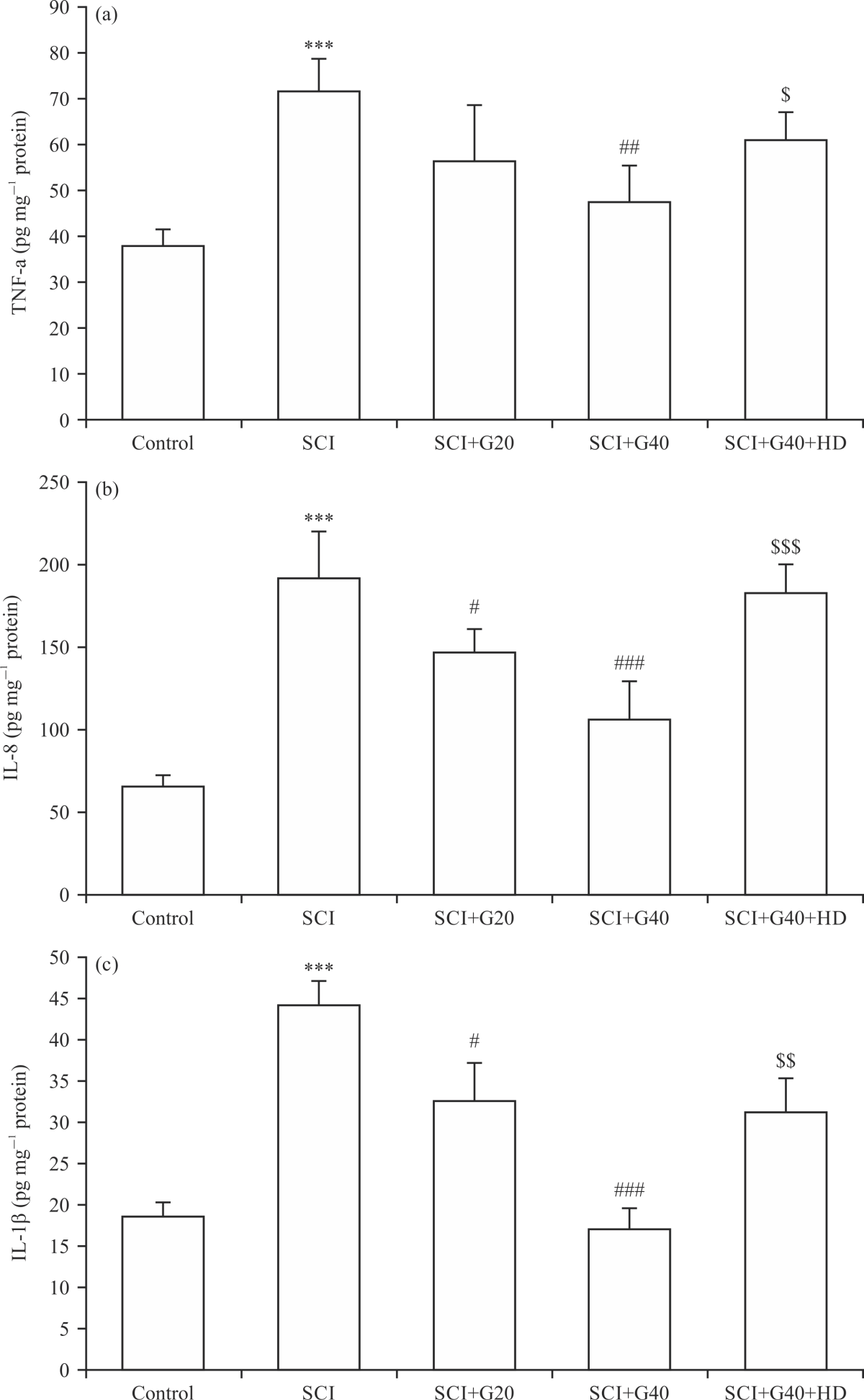 |
| Fig. 5(a-c): | Effects of GMC on the production of proinflammatory cytokines in SCI model, (a) TNF-α, (b) IL-8 and (c) IL-1β (n = 6), ***p<0.001 vs. control group, #p<0.05, ##p<0.01, ###p<0.001 vs. SCI group, $p<0.05, $$p<0.01, $$$p<0.001 vs. SCI+G40 group, SCI: Spinal cord injury, G: Ginsenoside Mc1, GMC and HD: Hydroxydecanoate |
The result of this situation is the exacerbation of SCI. In the present study, administration of GMC limited the production of pro-inflammatory TNF-α and other interleukins and suppressed mitochondrial oxidative stress (as assessed by the changes of mitochondrial ROS and MnSOD). These effects of the drug as well as its inhibitory effect against mitochondrial membrane depolarization indicate that this drug can modify mitochondrial internal homeostasis to a high degree in SCI conditions. Similarly, it has been reported that ginsenosides have beneficial impacts in patients with mitochondrial diseases including neuropathy13. It also interacts with peroxiredoxins, increasing their peroxidase activity and reducing ROS-induced cell death16. The active metabolites of this drug also reduced I/R-induced cardiac damages and prevented oxidative responses by activating the expression of AMPK10. All of these findings, together with our results, suggest that the mitochondrial-directed pathway is an important target for this drug and it is a good candidate for reducing SCI.
Mitochondrial dysfunction and subsequent inflammatory and oxidative responses are associated with alterations in mitochondrial biogenesis. Accumulating evidence shows that promoting mitochondrial biogenesis contributes to neuroprotection17. In the present study, we found that GMC was capable of inducing mitochondrial biogenesis, thereby providing adequate neuroprotection against SCI. Here, expressions of TFAM, NRF1 and PGC-1α were increased following GMC treatment in the spinal cord tissues. The PGC-1α is the initiating factor of mitochondrial biogenesis that induces the expression of TFAM in collaboration with NRF1, as its transcriptional partner18,19. The interplay between NRF1 and TFAM induces the transcription of mtDNA and stimulates mitochondrial oxidative/phosphorylation18. Although these findings imply that GMC neuroprotective effects may be achieved through stimulating the entire mitochondrial biogenesis program, the correlation between GMC-induced spinal cord protection and PGC-1α-activated mitochondrial recovery requires further study.
To further explore the role of GMC in controlling mitochondrial activity and biogenesis as well as inflammatory responses under SCI conditions, mitochondrial ATP-KCh were blocked using HD to examine the involvement of these channels in the protective effect of the drug. Blocking of ATP-KCh not only abolished the cord protective effects of GMC but also reversed its anti-inflammatory and mitochondrial actions. These findings highlight that the opening of these channels mediates the beneficial effects of GMC in SCI. In agreement with our results, it has been documented that ATP-KCh is critically involved in the regulation of PGC-1α to influence mitochondrial homeostasis6. The density of these channels in the neuronal cells is greater than other organs20 and this strongly emphasizes the importance of these mitochondrial channels in GMC actions on mitochondrial biogenesis and homeostasis (preventing mitochondrial impairment and membrane potential collapse) as well as anti-inflammation. These channels are considered as one of the main arms of mitochondria in their role as the end effector of neuroprotection7 and thereby this main cellular component mediates the protective influences of GMC in reducing SCI outcomes. Therefore, the neuroprotective effect of GMC in SCI was achieved through modulation of the mitochondrial ATP-KCh/mitochondrial biogenesis/inflammatory pathway. However, the contribution of other important mediators including PKG/cGMP and PI3K/AKT pathways in this effect requires further experimentation.
CONCLUSION
The GMC, as a new non-glycosylated ginsenoside compound, displayed strong spinal cord protection in SCI through anti-inflammatory, anti-oxidative and mitochondrial activity improving effects. Increased activity of mitochondrial ATP-KCh markedly mediated the beneficial actions of GMC. This study highlighted that this drug can be one of the most promising pharmacological approaches to target mitochondrial dysfunction in SCI conditions.
SIGNIFICANCE STATEMENT
This study explores the potential protective effects of GMC on the spinal cord, which could be beneficial in reducing SCI outcomes. This study helps the researcher to uncover the critical area of SCI and the involvement of mitochondrial function/biogenesis in its pathophysiology that can be reversed by GMC and this has not been previously explored. Thereby, a new theory may be developed about this protective treatment and possibly its mechanisms of action.
REFERENCES
- Liu, L., J. Zhou, Y. Wang, T. Qi, Z. Wang, L. Chen and N. Suo, 2020. Imatinib inhibits oxidative stress response in spinal cord injury rats by activating Nrf2/HO‑1 signaling pathway. Exp. Ther. Med., 19: 597-602.
CrossRefDirect Link - Mitchell, R., L. Harvey, R. Stanford and J. Close, 2018. Health outcomes and costs of acute traumatic spinal injury in New South Wales, Australia. Spine J., 18: 1172-1179.
CrossRefDirect Link - Anjum, A., M.D. Yazid, M.F. Daud, J. Idris and A.M.H. Ng et al., 2020. Spinal cord injury: Pathophysiology, multimolecular interactions, and underlying recovery mechanisms. Int. J. Mol. Sci., Vol. 21.
CrossRefDirect Link - Alizadeh, A., S.M. Dyck and S. Karimi-Abdolrezaee, 2019. Traumatic spinal cord injury: An overview of pathophysiology, models and acute injury mechanisms. Front. Neurol., Vol. 10.
CrossRefDirect Link - Cherry, A.D. and C.A. Piantadosi, 2015. Regulation of mitochondrial biogenesis and its intersection with inflammatory responses. Antioxid. Redox Signaling, 22: 965-976.
CrossRefDirect Link - Strickland, M., B. Yacoubi-Loueslati, B. Bouhaouala-Zahar, S.L.F. Pender and A. Larbi, 2019. Relationships between ion channels, mitochondrial functions and inflammation in human aging. Front. Physiol., Vol. 10.
CrossRefDirect Link - Sun, H.S. and Z.P. Feng, 2013. Neuroprotective role of ATP-sensitive potassium channels in cerebral ischemia. Acta Pharmacol. Sin., 34: 24-32.
CrossRefDirect Link - Rahman, M.A., H. Rahman, P. Biswas, M.S. Hossain and R. Islam et al., 2020. Potential therapeutic role of phytochemicals to mitigate mitochondrial dysfunctions in Alzheimer's disease. Antioxidants, Vol. 10.
CrossRefDirect Link - Huang, X., N. Li, Y. Pu, T. Zhang and B. Wang, 2019. Neuroprotective effects of ginseng phytochemicals: Recent perspectives. Molecules, Vol. 24.
CrossRefDirect Link - Hong, S.H., H.J. Hwang, J.W. Kim, J.A. Kim and Y.B. Lee et al., 2020. Ginsenoside compound-Mc1 attenuates oxidative stress and apoptosis in cardiomyocytes through an AMP-activated protein kinase–dependent mechanism. J. Ginseng Res., 44: 664-671.
CrossRefDirect Link - Roh, E., H.J Hwang, J.W Kim, S.H. Hong and J.A. Kim et al., 2020. Ginsenoside Mc1 improves liver steatosis and insulin resistance by attenuating ER stress. J. Ethnopharmacol., Vol. 259.
CrossRefDirect Link - Wang, H., J. Zhu, Z. Jia and S. Lu, 2022. Pretreatment of diabetic aged rats with combination of ginsenoside-Mc1 and silibinin protects liver from ischemia-reperfusion injury through an AMPK-dependent mechanism. Turk. J Biochem., 47: 23-32.
CrossRefDirect Link - Huang, Q., S. Gao, D. Zhao and X. Li, 2021. Review of ginsenosides targeting mitochondrial function to treat multiple disorders: Current status and perspectives. J. Ginseng Res., 45: 371-379.
CrossRefDirect Link - Cheng, G., R.H. Kong, L.M. Zhang and J.N. Zhang, 2012. Mitochondria in traumatic brain injury and mitochondrial-targeted multipotential therapeutic strategies. Br. J. Pharmacol., 167: 699-719.
CrossRefDirect Link - Liu, X., X. Gu, M. Yu, Y. Zi and H. Yu et al., 2018. Effects of ginsenoside Rb1 on oxidative stress injury in rat spinal cords by regulating the eNOS/Nrf2/HO‑1 signaling pathway. Exp. Ther. Med., 16: 1079-1086.
CrossRefDirect Link - Cong, L. and W. Chen, 2016. Neuroprotective effect of ginsenoside Rd in spinal cord injury rats. Basic Clin. Pharmacol. Toxicol., 119: 193-201.
CrossRefDirect Link - Stetler, R.A., R.K. Leak, W. Yin, L. Zhang, S. Wang, Y. Gao and J. Chen, 2012. Mitochondrial biogenesis contributes to ischemic neuroprotection afforded by LPS pre-conditioning. J. Neurochem., 123: 125-137.
CrossRefDirect Link - Gureev, A.P., E.A. Shaforostova and V.N. Popov, 2019. Regulation of mitochondrial biogenesis as a way for active longevity: Interaction between the Nrf2 and PGC-1α signaling pathways. Front. Genet., Vol. 10.
CrossRefDirect Link - Nanjaiah, H. and B. Vallikannan, 2019. Lutein upregulates the PGC‐1α, NRF1, and TFAM expression by AMPK activation and downregulates ROS to maintain mtDNA integrity and mitochondrial biogenesis in hyperglycemic ARPE‐19 cells and rat retina. Biotechnol. Appl. Biochem., 66: 999-1009.
CrossRefDirect Link - Cui, J., J. Fan, H. Li, J. Zhang and J. Tong, 2021. Neuroprotective potential of fisetin in an experimental model of spinal cord injury: Via modulation of NF-κB/IκBα pathway. Neuro Rep., 32: 296-305.
CrossRefDirect Link