
Omer Bahceli
Department of Pharmacology, Faculty of Pharmacy, Mersin University, Mersin 33160, Turkey
Sefika Pinar Senol
Department of Pharmacology, Faculty of Pharmacy, Mersin University, Mersin 33160, Turkey
Meryem Temiz-Resitoglu
Department of Pharmacology, Faculty of Pharmacy, Mersin University, Mersin 33160, Turkey
Mehmet Furkan Horat
Department of Pharmacology, Faculty of Pharmacy, Mersin University, Mersin 33160, Turkey
Seyhan Sahan-Firat
Department of Pharmacology, Faculty of Pharmacy, Mersin University, Mersin 33160, Turkey
Bahar Tunctan
Department of Pharmacology, Faculty of Pharmacy, Mersin University, Mersin 33160, Turkey
LiveDNA: 90.16197
International Journal of Pharmacology
Year: 2022 | Volume: 18 | Issue: 6 | Page No.: 1171-1188







ABSTRACT
Background and Objective: One of the clinically approved retinoid X receptor (RXR) ligands, bexarotene, has been demonstrated to exert analgesic and anti-inflammatory effects in the experimental models of many inflammatory neurological diseases. The previous studies showed that bexarotene ameliorates the lipopolysaccharide (LPS)-induced hyperalgesia by increasing RXR expression associated with suppressed toll-like receptor (TLR)/myeloid differentiation factor (MyD) 88/transforming growth factor beta-activated kinase 1/nuclear factor-κB/cyclooxygenase-2 signaling pathway activity in the central nervous system (CNS). This study tested the hypothesis that the TLR/MyD88-dependent pro-inflammatory and anti-apoptotic signaling pathways mediate the ameliorating effect of bexarotene against LPS-induced inflammatory hyperalgesia. Materials and Methods: Brain and spinal cord tissues of bexarotene-treated mice were used for the measurement of tumor necrosis factor receptor-associated factor (TRAF) 6, inhibitor of IkB kinase (IKK) α, IKKβ, IKKγ, inhibitor of κB (IkB)-α, mitogen-activated protein kinase kinase (MEK), extracellular signal-regulated kinase (ERK) 1/2, c-Jun, phosphoinositide 3-kinase (PI3K) p85α, Akt1, cyclic adenosine monophosphate-response element-binding protein (CREB), B-cell lymphoma (Bcl)-2, peroxisome proliferator-activated receptor (PPAR) α, PPARβ and PPARγ expression and/or activity by using the immunoblotting method. Results: Results showed that diminished protein expression and/or activity of PI3K p85α, Akt1, CREB1, Bcl-2 and PPARα/β/γ was associated with enhanced TRAF6, IKKα/β/γ, IkB-α, MEK1/2, ERK1/2 and c-jun proteins in the tissues of endotoxemic mice. These changes were ameliorated following bexarotene treatment. Conclusion: Thus, decreased activity of pro-inflammatory TLR4/MyD88-dependent TRAF6/IKKα/β/γ/IκB-γ and TRAF6/MEK1/2/ERK1/2/AP-1 signaling pathways associated with an increase in the anti-apoptotic PI3K p85α/Akt1/CREB1/Bcl-2 signaling pathway activity and anti-inflammatory PPARα/β/γ proteins in the CNS contributes to the ameliorating effect of bexarotene against LPS-induced inflammatory hyperalgesia in mice.
PDF Abstract XML References Citation
Copyright: © 2022. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Omer Bahceli, Sefika Pinar Senol, Meryem Temiz-Resitoglu, Mehmet Furkan Horat, Seyhan Sahan-Firat and Bahar Tunctan, 2022. Bexarotene Ameliorates LPS-Induced Hyperalgesia: Contribution of TLR4/MyD88-Dependent Pro-Inflammatory, Anti-Apoptotic and Anti-Inflammatory Signaling Pathways. International Journal of Pharmacology, 18: 1171-1188.
DOI: 10.3923/ijp.2022.1171.1188
URL: https://scialert.net/abstract/?doi=ijp.2022.1171.1188
DOI: 10.3923/ijp.2022.1171.1188
URL: https://scialert.net/abstract/?doi=ijp.2022.1171.1188
INTRODUCTION
Retinoid X receptor (RXRs), as members of the nuclear receptor superfamily of ligand-activated transcription factors, are participated in the nuclear receptors that regulate various physiological cellular events, including cell survival1. The only food and drug administration-approved rexinoid, RXR agonist bexarotene, has been reported to have analgesic and anti-inflammatory effects in the experimental models of many inflammatory neurological diseases2,3. Bexarotene has also been shown to have beneficial effects in the treatment of neurodegenerative diseases characterized by systemic inflammation (e.g., Alzheimer's disease, Parkinson's disease, neuroinflammation and neuropathic pain) in animal models4-6. However, the mechanisms underlying these effects are still not fully understood, but may involve regulation of the toll-like receptor (TLR4)/myeloid differentiation factor (MyD) 88-mediated inflammatory pathway in the central nervous system (CNS).
The lipid A part of lipopolysaccharide (LPS), endotoxin, has been shown to enhance pain sensation to thermal stimuli at supraspinal and spinal levels as determined by the hot plate test, which is considered to integrate supraspinal pathways and to be a supraspinally controlled acute pain test7. We have previously demonstrated that nitric oxide (NO) reverses inflammatory hyperalgesia induced by LPS through reducing prostacyclin formation and also participates in the antinociceptive effect of cyclooxygenase (COX), nuclear factor-κB (NF-κB), or poly-adenosine diphosphate ribose synthase inhibitors8. Furthermore, nucleotide-binding domain and leucine-rich repeat protein (NLRP3) inflammasome inhibitor, MCC950, or soluble epoxide hydrolase inhibitor, trifluoromethoxyphenyl-3-(1-propionylpiperidin-4-yl)urea, prevented not only the inflammatory hyperalgesia but also LPS-induced (1) Enhanced activity of the NLRP3/apoptosis-associated speck-like protein containing a caspase activation and recruitment domain (ASC)/pro-caspase-1, NLRC4/ASC/ pro-caspase-1 and caspase-11 inflammasomes, TLR4/MyD88/ transforming growth factor-activated kinase/NF-κB/COX-2 pathway and nicotinamide adenine dinucleotide phosphate oxidase associated with enhanced pro-inflammatory cytokine formation, (2) Reduced levels of anti-inflammatory cytochrome P450 (CYP) epoxygenase-derived epoxyeicosatrienoic acids and (3) Diminished expression of inducible NO synthase (iNOS), neuronal NO synthase, NLRC3 and peroxisome proliferator-activated receptor (PPAR) α/β/γ in the various tissues of mice including brains and spinal cords8-10. In the recent study11, we demonstrated that treatment of mice with bexarotene ameliorated the lipopolysaccharide (LPS)-induced hyperalgesia by increasing protein expression of RXR and suppressing NF-κB-mediated COX-2 expression and prostaglandin E2 formation, which was regulated by TLR4/MyD88-dependent transforming growth factor beta-activated kinase (TAK) 1 signaling pathway in the CNS. As a continuation of our previous work, we aimed to test the hypothesis that RXR agonists such as bexarotene may be useful in preventing the events that may also result in inflammatory hyperalgesia accompanied by the increase in the TLR4/MyD88-dependent pro-inflammatory signaling pathway activity associated with suppressed activity of anti-apoptotic phosphoinositide 3-kinase (PI3K)/Akt1/cyclic adenosine monophosphate-response element-binding protein (CREB) 1/B-cell lymphoma (Bcl)-2 pathway caused by LPS in the CNS.
MATERIALS AND METHODS
Animals and tissues: The brain and spinal cord tissues of male Balb/c mice (20-40 g, n = 20) used in the experiments were housed under standard conditions with free access to food and water and a 12 h light-dark cycle. The animals were obtained from the Research Center of Experimental Animals, Mersin University, Mersin, Turkey. The experimental protocol was approved by the Mersin University Experimental Animals Local Ethics Committee (decision numbers: 2017/17 and 2018/20). All experiments were carried out according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals and the American Veterinary Medical Association Guidelines for the Euthanasia of Animals 2013 Edition. The study was carried out from March to October, 2021.
Inflammatory hyperalgesia model: In the previous study11, the inflammatory hyperalgesia model was induced by the injection of LPS intraperitoneally (i.p.) into mice and the latency of pain to nociceptive response was measured by the hot plate test as described previously8,10,12. The mice were injected with saline (10 mL kg–1, i.p.) or LPS (10 mg kg–1, i.p.) (Escherichia coli. LPS, O111:B4) 6 hrs before the thermal stimulus. Dimethyl sulfoxide (DMSO) (4 mL kg–1) or bexarotene (4 mL kg–1) (Sigma Chemical Co., St., Louis, MO, USA) was administered subcutaneously at the same time with saline or LPS. The LPS (L4130) and bexarotene (SML0282) were purchased from Sigma Chemical Co. (St., Louis, MO, USA). DMSO was purchased from Applichem Inc. Maryland Heights, MO, USA. Saline and DMSO (a 1% [v/v] dilution of DMSO in saline) were used to dissolve LPS and bexarotene, respectively. Mice treated with saline, LPS, or DMSO were used as control, inflammatory hyperalgesia and vehicle control groups, respectively. To observe pain behavior, a hot plate apparatus (AHP 9601, Commat Ltd., Ankara, Turkey) maintained at temperature of 55±0.2°C was used to determine paw licking time within 30 sec after 6 hrs of drug administration. Euthanasia was carried out by cervical dislocation of mice after the test and brains and spinal cords were collected from all animals.
Immunoblotting: The immunoblotting method was used according to the protocol as described in detail previously with minor modifications10,11,13,14. Briefly, tissue homogenates (30 mg of protein) were subjected to a 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. After electrophoresis was complete, proteins were transferred from the gel to nitrocellulose membranes. The membranes were blocked with non-fat dry milk in Tris-buffered saline and incubated overnight with the following primary antibodies in bovine serum albumin (BSA) at 4°C against (1) Tumor necrosis factor (TNF) receptor-associated factor (TRAF) 6 (sc-8409, Santa Cruz Biotechnology, Santa Cruz, CA, USA), (2) IKKα antibody (2682, Cell Signaling, Danvers, MA, USA), (3) Inhibitor of κB kinase (IKK) β (8943, cell signaling), (4) Phosphorylated IKKα/IKKβ (p-IKKα/IKKβ) (2078, cell signaling), (5) IKKγ (sc-71331, Santa Cruz), (6) phosphorylated IKKγ (p-IKKγ) (sc-293135, Santa Cruz), (7) inhibitor of κB (IkB)-α antibody (sc-1643, Santa Cruz), (8) Phosphorylated IkB-α (p-IkB-α) (sc-7977, Santa Cruz), (9) mitogen-activated protein kinase (MEK1/2) (sc-81504, Santa Cruz), (10) phosphorylated MEK1/2 (p-MEK1/2) (sc-81503, Santa Cruz), (11) extracellular signal-regulated kinase (ERK) 1/2 (sc-514302, Santa Cruz), (12) phosphorylated ERK1/2 (p-ERK1/2) (sc-136521, Santa Cruz), (13) c-Jun (sc-1694, Santa Cruz), (14) phosphorylated c-Jun (p-c-Jun) (sc-16312, Santa Cruz), (15) PI3K p85α (OAAF05915, Aviva Systems Biology, San Diego, CA, USA), (16) phosphorylated PI3K p85α (p-PI3K p85α) (OAEC00301, Aviva Systems Biology), (17) Akt1 (sc-271149, Santa Cruz), (18) phosphorylated Akt1 (p-Akt1) (sc-52940, Santa Cruz), (19) CREB1 (sc-271, Santa Cruz), (20) phosphorylated CREB1 (p-CREB1) (sc-81486, Santa Cruz), (21) Bcl-2 (sc-7382, Santa Cruz), (22) PPARα (sc-9000, Santa Cruz), (23) PPARβ (sc-74517, Santa Cruz) and (24) PPARγ (sc-7273, Santa Cruz). The membranes were subsequently incubated for 1 h at room temperature with an appropriate secondary antibody solution in BSA containing a sheep anti-mouse IgG-horseradish peroxidase or a goat anti-rabbit IgG-horseradish peroxidase (RPN4201 or RPN4301, respectively, Amersham Life Sciences, Cleveland, OH, USA). The membranes were then incubated wit an ECL Prime Western Blotting Detection Reagent (RPN2232, Amersham). A gel-imaging system (EC3-CHEMI HR imaging system, Ultra-Violet Products, UVP, Cambridge, UK) was use to image specific bands for the immunoreactive proteins on the membranes. To determine the relative immunoreactive bands densities, Image J densitometry analysis software (Image J 1.46r, Wayne Rasband, National Institute of Health, Bethesda, MD, USA) was used. The membranes were reused for anti-β-tubulin antibody (sc-5274, Santa Cruz) as a loading control. The ratio of each band/β-tubulin was taken into account for the expression level of specific proteins.
Statistical analysis: The sample size determined for each treatment group in each experiment was based on previous studies from our laboratory10-12 and complied with power analysis15. The results are expressed as Means±Standard Error of the Mean (SEM). Since the data were normally distributed, parametric statistical analysis was conducted using the Student's t test. Statistical analysis was performed using GraphPad Prism version 7.04 for Windows (GraphPad Software, San Diego California USA, http://www.graphpad.com). A p<0.05 was considered to be statistically significant.
RESULTS
LPS-induced increase in the TRAF6 expression was prevented by bexarotene treatment: First, we tested whether bexarotene affects the expression of TRAF6, which is an important adaptor molecule in the pro-inflammatory TLR4/MyD88-dependent IKK/IκB-α/NF-κB and MAPK/AP-1 signaling pathways in the CNS, in response to LPS. The brain and spinal cord tissue samples of saline-, LPS- and/or bexarotene-treated mice were immunoblotted with a specific TRAF6 antibody. Enhanced expression of TRAF6 was observed in the tissues of LPS-treated mice compared with the control group values (p<0.05) (Fig. 1). Treatment with bexarotene inhibited the LPS-induced increase in TRAF6 expression in the tissues compared with LPS-injected mice (p<0.05). Expression of TRAF6 in the tissues of bexarotene-treated mice was not different from the control group values (p>0.05).
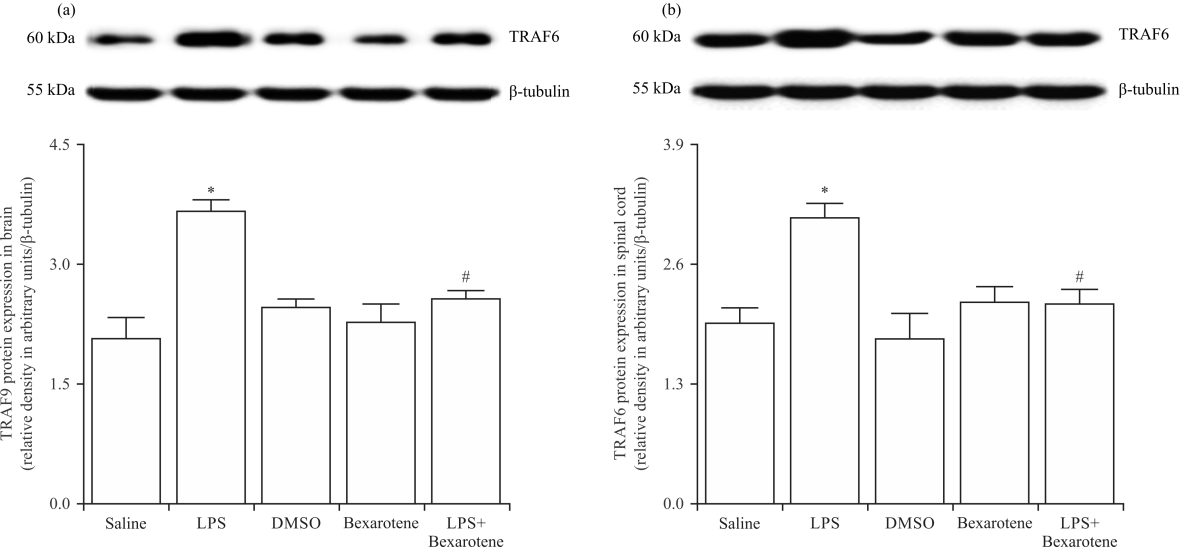 |
| Fig. 1(a-b): | Bexarotene ameliorated the LPS-induced increase in TRAF6 protein expression in the (a) Brain and (b) Spinal cord tissues of control and endotoxemic mice |
Protein expression of TRAF6 in the tissues was measured by immunoblotting, data are expressed as the Mean±SEM of 4 animals, *p<0.05 vs. saline-injected group and #p<0.05 vs. LPS-injected group
LPS-induced increase in the activity of IKK/IκB-α signaling pathway was prevented by bexarotene treatment: In the second experimental approach, we tested whether bexarotene affects the expression and activity of IKK/IkB-α signaling pathway in the CNS, the brain and spinal cord tissue samples of saline-, LPS- and/or bexarotene-treated mice were immunoblotted with specific antibodies for IKKα, IKKβ, p-IKKα/IKKβ (at Ser176 and Ser177, respectively), IKKγ, p-IKKγ (at Ser376), IkB-α and p-IkB-α (at Ser32). As shown in Fig. 2, expression of the phosphorylated, but not unphosphorylated, catalytic (IKKα and IKKβ) and regulatory (IKKγ) subunits of the IKK complex proteins was enhanced in the tissues of LPS-injected mice compared with the control group values (p<0.05). Reduced expression of IkB-α was also associated with the increased p-IkB-α expression in the tissues of LPS-injected mice compared with the control group values (p<0.05). The changes in the expression of these proteins in the mice injected with LPS were inhibited by bexarotene (p<0.05). Expression of unphosphorylated and phosphorylated proteins in the tissues of bexarotene-treated mice was not different from the control group values (p>0.05).
LPS-induced increase in the activity of MEK1/2, ERK1/2 and c-Jun was prevented by bexarotene treatment: To further test whether bexarotene affects the expression and activity of MEK1/2, ERK1/2 and c-Jun (a key subunit of the activator protein [AP]-1) in the CNS, the brain and spinal cord tissue samples of saline-, LPS- and/or bexarotene-treated mice were immunoblotted with specific antibodies for MEK1/2, p-MEK1/2 (at Ser218/Ser222 for MEK1 and Ser222/Ser226 for MEK2), ERK1/2, p-ERK1/2 (at Thr203/Tyr205 for ERK1 and (at Thr183/Tyr185 for ERK2), c-Jun and p-c-Jun (at Ser63/Ser73). Expression of the phosphorylated, but not unphosphorylated (with the exception of c-Jun), proteins was enhanced in the tissues of LPS-injected mice compared with the control group values (p<0.05) (Fig. 3). The enhanced expression of these proteins in the mice injected with LPS was inhibited by bexarotene (p<0.05). Expression of unphosphorylated and phosphorylated proteins in the tissues of bexarotene-treated mice was not different from the control group values (p>0.05).
LPS-induced decrease in the activity of PI3K/Akt1/CREB1/ Bcl-2 pathway was prevented by bexarotene treatment: To test the hypothesis that bexarotene also prevents LPS-induced decrease in the anti-apoptotic PI3K/Akt1/CREB1/Bcl-2 pathway activity in the CNS, the brain and spinal cord tissue samples of saline-, LPS- and/or bexarotene-treated mice were immunoblotted with specific antibodies for PI3K p85α, p-PI3K p85α (at Tyr467), Akt1, p-Akt1 (at Ser473), CREB1, p-CREB1 (at Ser133) and Bcl-2. As shown in Fig. 4, expression of p-PI3K p85α, p-Akt1, p-CREB1 and Bcl-2 proteins was diminished in the tissues of LPS-injected mice compared with the control group values (p<0.05). The decrease in the p-PI3K p85α, p-Akt1, p-CREB1 and Bcl-2 expression in the mice injected with LPS was inhibited by bexarotene (p<0.05). Expression of unphosphorylated and/or phosphorylated PI3K p85α, Akt1, CREB1 and Bcl-2 proteins in the tissues of bexarotene-treated mice was not different from the control group values (p>0.05).
LPS-induced reduction in the expression of PPARα/β/γ was prevented by bexarotene treatment: To further investigate whether bexarotene also affects the expression of PPARα/β/γ, an important family of ligand-activated nuclear transcription factors, in the CNS, the brain and spinal cord tissue samples of saline-, LPS- and/or bexarotene-treated mice were immunoblotted with specific antibodies for PPARα, PPARβ and PPARγ.
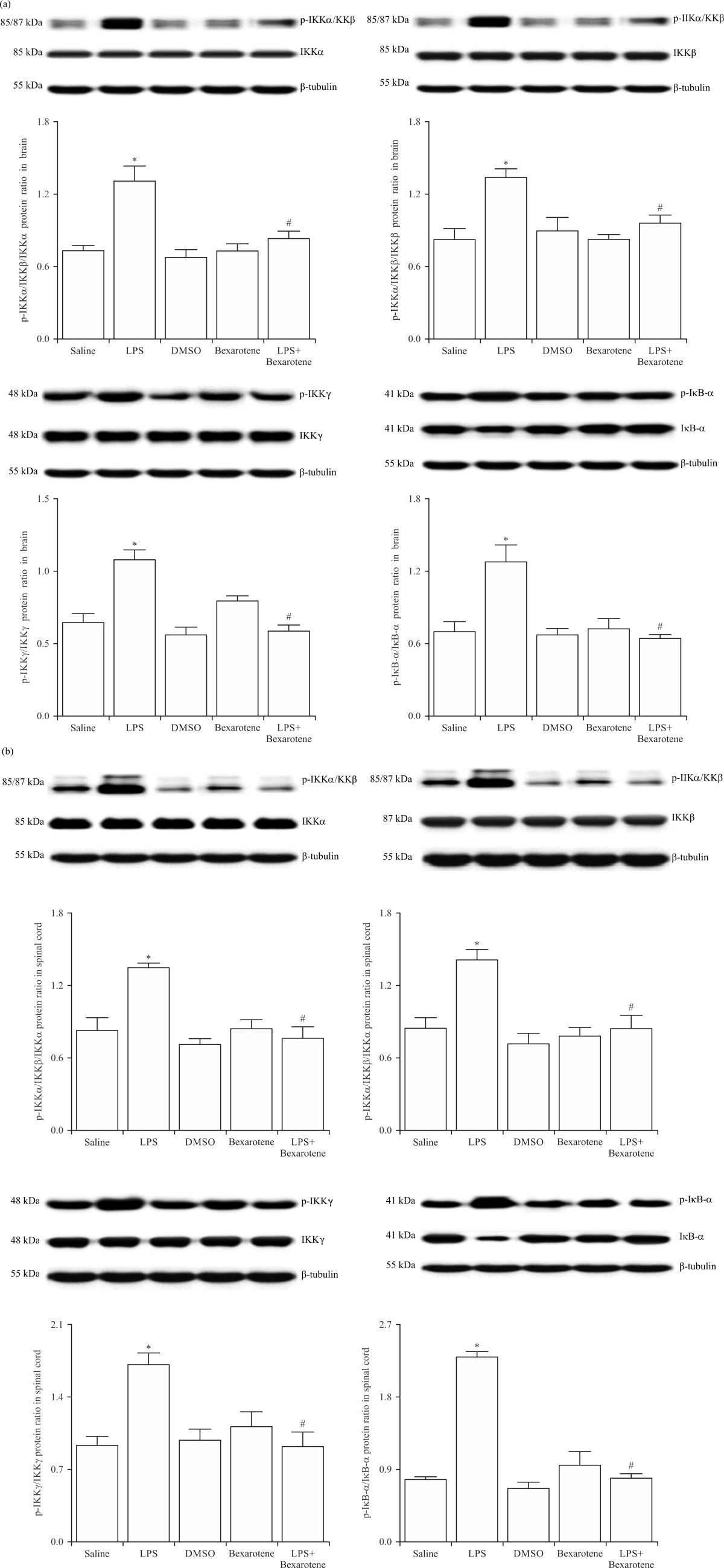 |
| Fig. 2(a-b): | Bexarotene ameliorated the LPS-induced changes in IKKα, IKKβ, IKKγ and IκB-α protein phosphorylation in the (a) Brain (b) Spinal cord tissues of control and endotoxemic mice Expression of unphosphorylated and phosphorylated IKKα, IKKβ, IKKγ and IkB-a proteins in the tissues was measured by immunoblotting, data are expressed as the Mean±SEM of 4 animals, *p<0.05 vs. saline-injected group and #p<0.05 vs. LPS-injected group |
Expression of PPARα, PPARβ and PPARγ proteins was diminished in the tissues of LPS-injected mice compared with the control group values (p<0.05) (Fig. 5). The reduction in the expression of PPARα, PPARβ and PPARγ proteins in the mice injected with LPS was inhibited by bexarotene (p<0.05). Expression of PPARα, PPARβ and PPARγ in the tissues of bexarotene-treated mice was not different from the control group values (p>0.05).
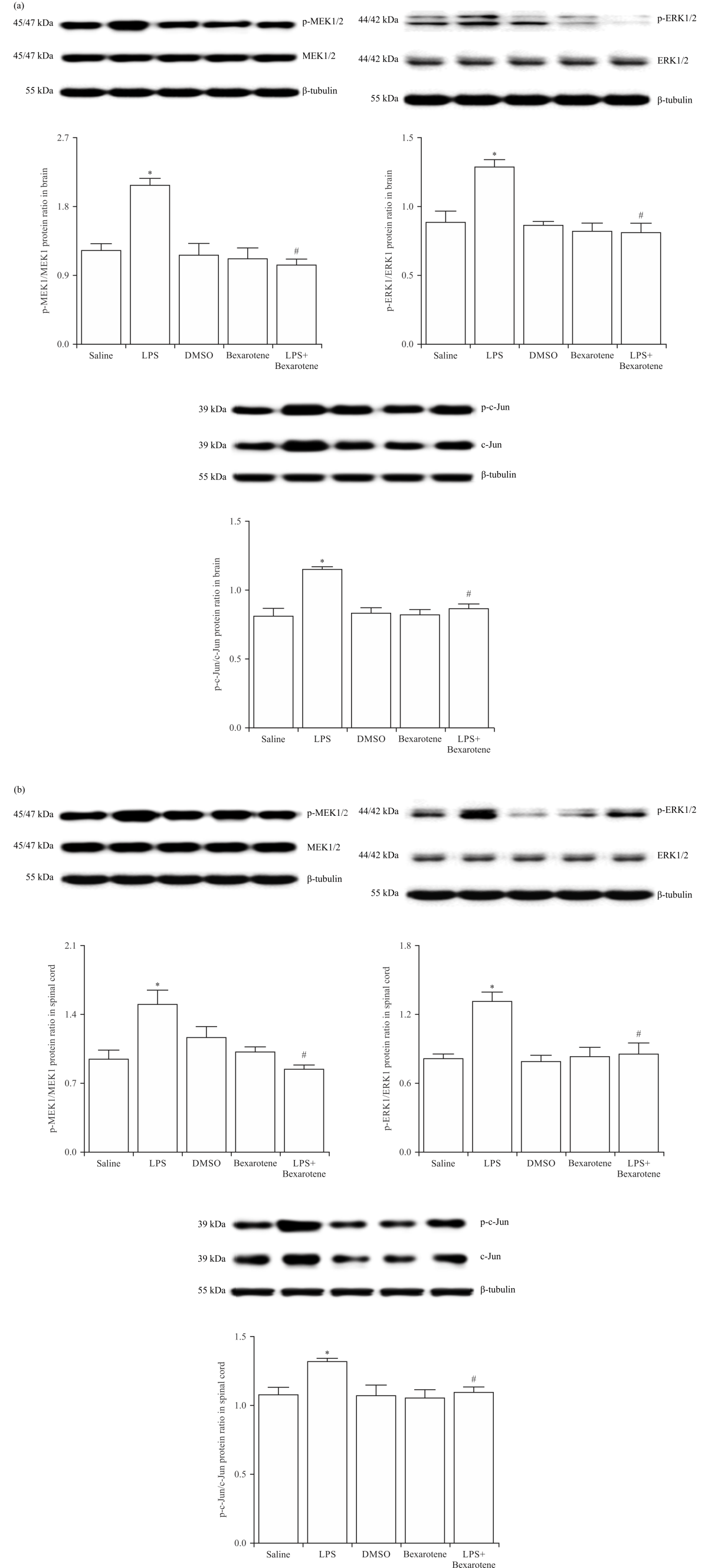 |
| Fig. 3(a-b): | Bexarotene ameliorated the LPS-induced changes in MEK1/2, ERK1/2 and c-Jun protein expression and/or phosphorylation in the (a) Brain and (b) Spinal cord tissues of control and endotoxemic mice Expression of unphosphorylated and phosphorylated MEK1/2, ERK1/2 and c-Jun proteins in the tissues was measured by immunoblotting, data are expressed as the Mean±SEM of 4 animals, *p<0.05 vs. saline-injected group and #p<0.05 vs. LPS-injected group |
DISCUSSION
The findings of the present study ensure the first evidence that suppression of the pro-inflammatory TLR4/MyD88-dependent TRAF6/IKKα/β/γ/IκB-α and TRAF6/MEK1/2/ERK1/2/AP-1 signaling pathways as well as the increase in the activity of anti-apoptotic PI3K p85α/Akt1/CREB1/Bcl-2 signaling pathway in addition to anti-inflammatory PPARα/β/γ proteins in the CNS of mice involves in the ameliorating effect of bexarotene against inflammatory hyperalgesia induced by LPS (Fig. 6).
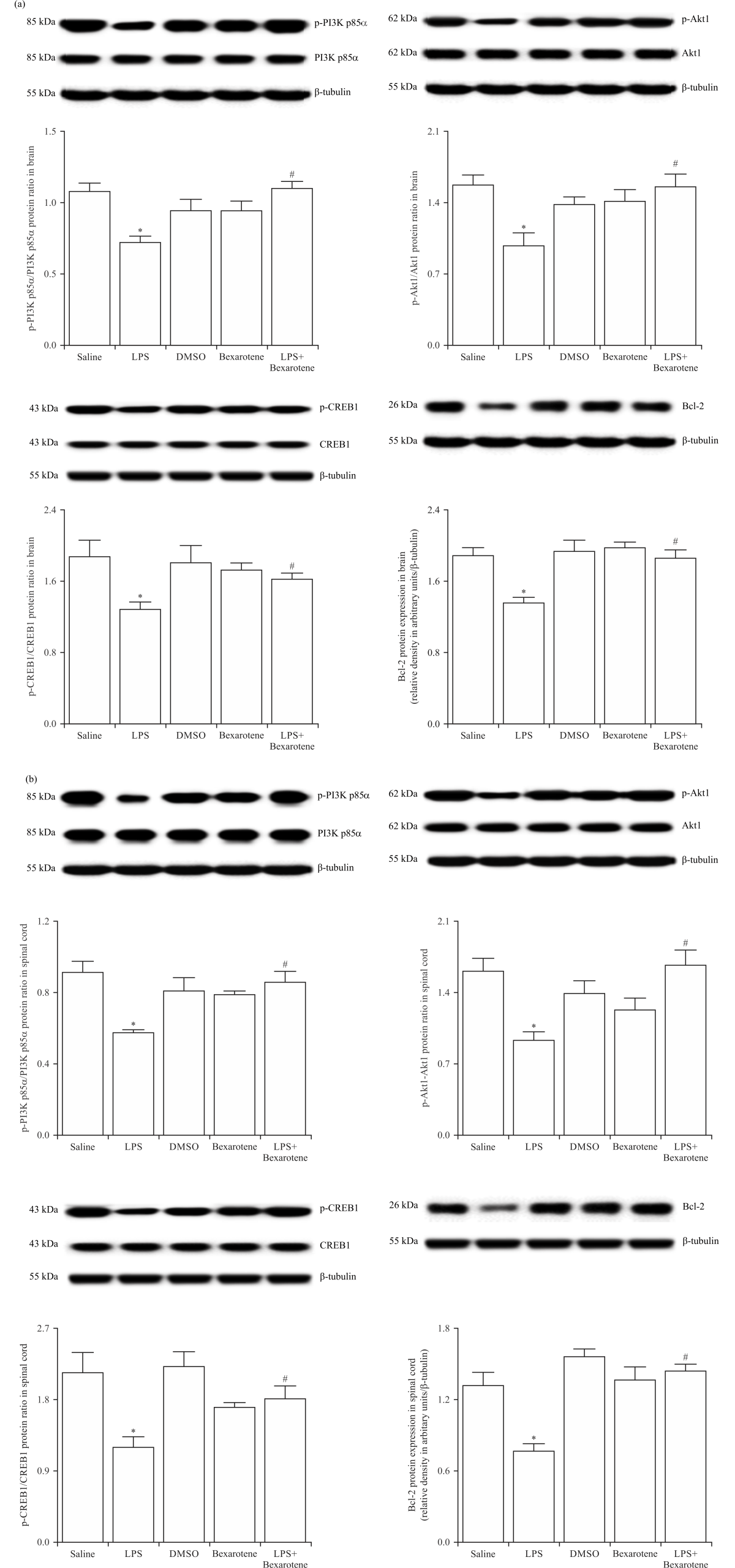 |
| Fig. 4(a-b): | Bexarotene ameliorated the LPS-induced decrease in PI3K p85α, Akt1, CREB1 and Bcl-2 protein expression and/or phosphorylation in the (a) Brain and (b) Spinal cord tissues of control and endotoxemic mice Expression of unphosphorylated and phosphorylated PI3K p85α, Akt1, CREB1 and Bcl-2 proteins in the tissues was measured by immunoblotting, data are expressed as the Mean±SEM of 4 animals, *p<0.05 vs. saline-injected group and #p<0.05 vs. LPS-injected group |
Recent studies have indicate that TRAF6 is an important adaptor molecule in the pro-inflammatory TLR4/MyD88-dependent IKK/IκB-α/NF-κB and MAPK/AP-1 signaling pathways in response to LPS16,17. There is also accumulating evidence suggesting that a reduction in the activity of the anti-apoptotic PI3K/Akt1/CREB1/Bcl-2 pathway associated with the enhanced activity of the TLR4/MyD88 inflammatory pathway, decreased cell survival and increased apoptosis18,19. As a family of ligand-activated nuclear transcription factors, PPARs have also been shown to have a special role in the regulation of the neuroinflammatory process20,21. Although there are other in vivo studies in the literature investigating the contribution of TAK1, TLR4, MyD88, NF-κB, ERK1/2 and RXRa to the pathogenesis of inflammatory hyperalgesia caused by LPS22-24, no studies were found regarding the contribution of TRAF6, IKK complex, IkB-α, MEK1/2, AP-1, Akt1, CREB1, Bcl-2 and PPARα/β/γ.
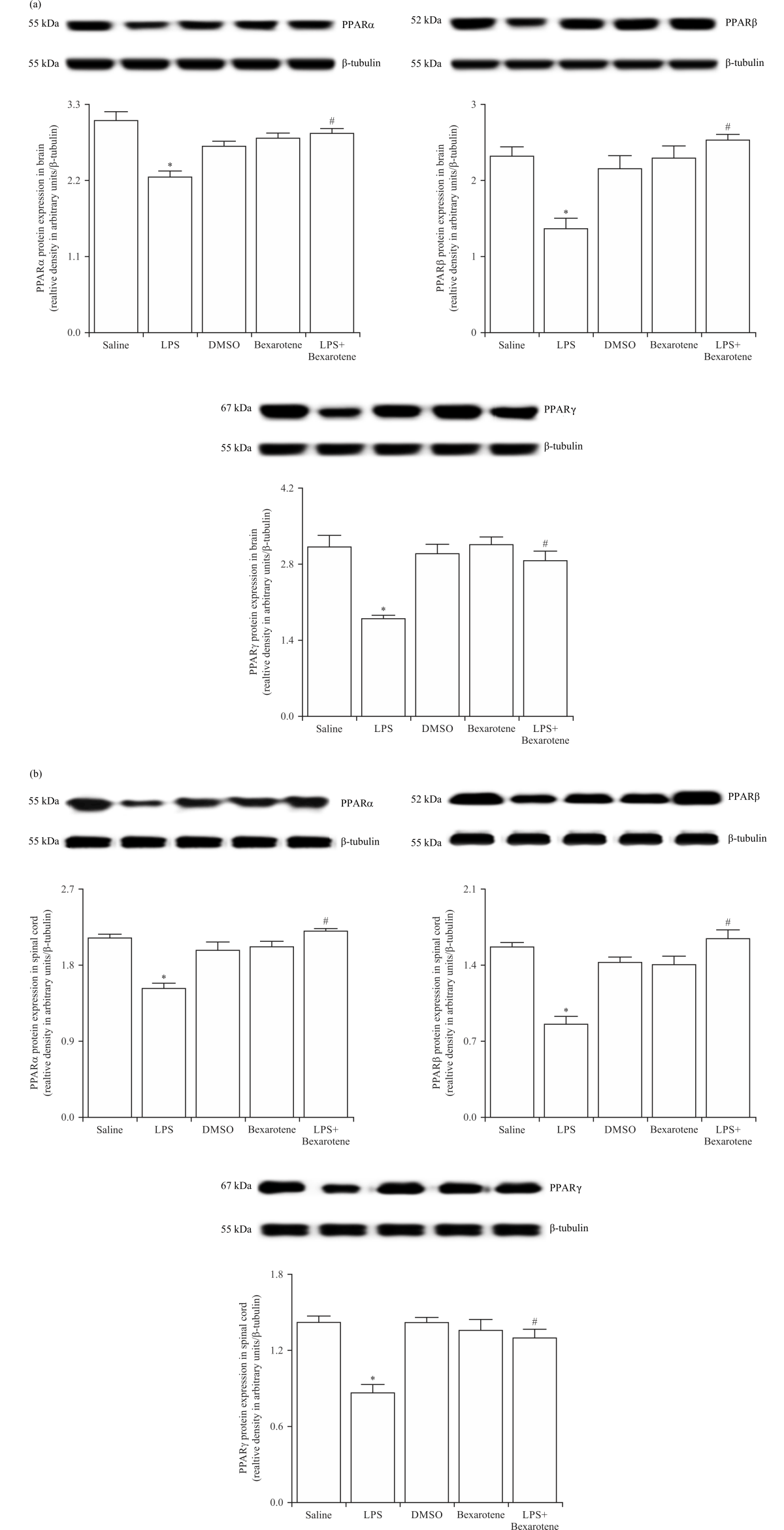 |
| Fig. 5(a-b): | Bexarotene ameliorated the LPS-induced decrease in PPARα, PPARβ and PPARγ protein expression in the (a) Brain and (b) Spinal cord tissues of control and endotoxemic mice Expression of PPARα, PPARβ and PPARγ proteins in the tissues was measured by immunoblotting, data are expressed as the Mean±SEM of 4 animals, *p<0.05 vs. saline-injected group, and #p<0.05 vs. LPS-injected group |
An important point to emphasize is that the results obtained from in vitro and in vivo studies using LPS are inconsistent. For instance, LPS has been reported to be a potent stimulus of TLR4-mediated inflammation in the range of 1-300 ng mL–1 in vitro25. Moreover, according to the results obtained from studies conducted under in vitro conditions, reduced expression of PPARγ26, Bcl-227 and CREB phosphorylation26,28 was associated with enhanced activity of MEK1/229,30, ERK1/231-33 and AP-131,32,34,35 in the brain and/or spinal cord tissues of mice as a result of LPS administration.
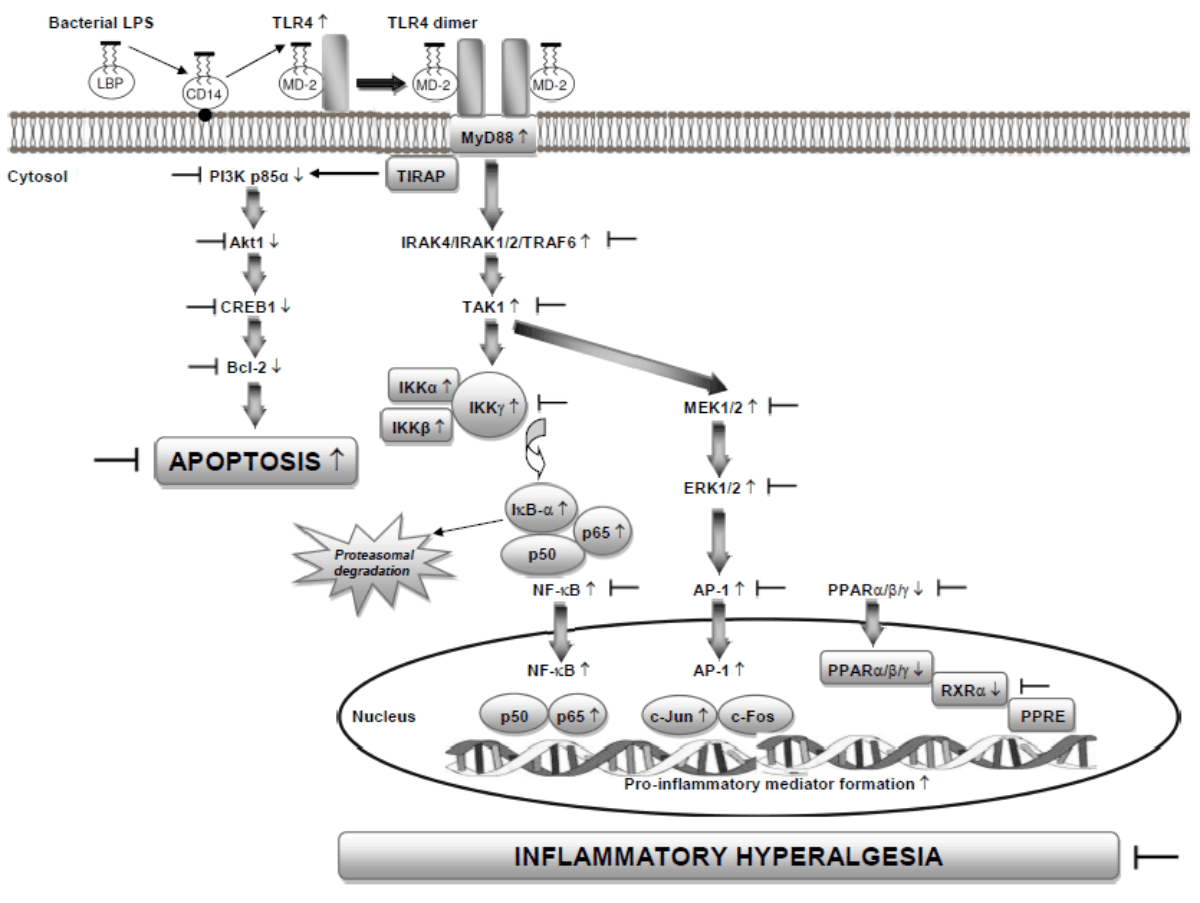 |
| Fig. 6: | Effects of bexarotene on the changes induced by LPS in the pro-inflammatory TLR4/MyD88-dependent TRAF6/IKKα/β/γ/IκB-α and TRAF6/MEK1/2/ERK1/2/AP-1 signaling pathways as well as increase in the anti-apoptotic PI3K p85α/Akt1/CREB1/Bcl-2 signaling pathway in addition to anti-inflammatory PPARα/β/γ proteins in the CNS during inflammatory hyperalgesia in mice based on the findings of the present study in addition to our previous findings8-11 ↑: Increased and ↓: decreased |
There were also studies showing that LPS not only enhances activities of PI3K26 and Akt26,31-33, but also diminishes their activities27,36. A very small amount of LPS has been shown to cause a low degree of inflammation by activating the TLR4 complex I/MyD88/glycogen synthase kinase 3/c-Jun N-terminal kinase (JNK) pathway, while it has been reported to have an anti-inflammatory effect by suppressing the TLR4 complex I/MyD88/TRIF/Akt/ERK pathway as a compensator to prevent further progression of the inflammatory response37. So, a low amount of LPS can suppress instead of activating the compensatory and anti-inflammatory PI3K and negative regulators of inflammatory genes such as CREB and interleukin (IL)-10. In addition, a low or high amount of LPS activates IKKs, NF-κB, p38 MAPK and JNK via TLR4 complex I/MyD88/toll-IL-1 receptor domain-containing adapter-inducing interferon-β (TRIF)/Akt/ERK via TLR4 complex II/TRIF/MyD88/toll/IL-1 receptor-associated kinases resulting in an acute, strong and transient inflammation. On the other hand, it has been reported that LPS at 0.2 mg kg–1 (i.p.) dose (in the low dose range 0.1-1 mg kg–1) has a neuroprotective effect, while, at 5 mg kg–1 (i.p.) dose in a high dose, it causes persistent inflammation and a progressive neurotoxic effect for up to 10 months38. According to the results obtained from in vivo studies, it has been reported that (1) Intracerebroventricular (i.c.v.) injection of LPS diminishes PI3K/Akt/Bcl-2 activity and PPARγ expression at a 1 ml/area dose39 as well as Bcl-2 levels at a 250 mL kg–1 dose40 in the brains of mice. The LPS injection at a dose of 1 mg kg–1 (i.p.) for 7 days is reported to cause an enhancement in the TLR4, NF-κB p65 and ERK1/2 activity, while it decreases Bcl-2 levels and did not change Akt activity41. When administered first intragastric at a dose of 0.5 mg kg–1 and then at a dose of 1.2 mg kg–1 (i.p.), LPS also diminished in the CREB activity28. Moreover, injection of LPS at 3, 5, or 8 mg kg–1 (i.p.) doses is reported to reduce42 or enhance the activity of ERK1/2 and CREB43 and decrease CREB activity44, respectively. In addition, LPS injection to mice (1) At a dose of 5 mg kg–1 (intrathecal) reduces PI3K/Akt activity in the lung45, (2) At a dose of 5 mg kg–1 (i.p.) enhances PI3K/Akt/CREB activity in the liver46, (3) At a dose of 8 mg kg–1 (i.p.) increases PI3K/Akt/Bcl-2 activity in the lung47. In addition, it was found that LPS injection to rats resulted in enhanced Akt/Bcl-2 activity at a dose of 10 μg per rat (i.c.v.) in the brain48 as well as suppressed PI3K/Akt activity at a dose of 5 mg kg–1 (i.p.)46 and Akt activity at a dose of 14 mg kg–1 (intravenous)49 in the lung. On the other hand, LPS can also induce a peripheral inflammatory stimulus on microglia and the brain at doses of 0.33-200 mg kg–1 (i.p.)50. It has been suggested that these conflicting results may be due to the strain, amount or dose of LPS, route and duration of administration and the test method used, as well as the type of experimental animal used in the model51,52. In the current study, we found that systemic administration of LPS (10 mg kg–1, i.p.) to mice resulted in hyperalgesia associated with (1) enhanced expression of TRAF6, p-IKKα/β/γ, p-IkB-α, p-MEK1/2, p-ERK1/2, c-Jun and p-c-Jun proteins and (2) Diminished expression of IkB-α, p-PI3K p85α, p-Akt1, p-CREB1, Bcl-2, PPARα/β/γ proteins in the brain and spinal cord tissues. Hence, in line with the previous findings8-11 and the above-mentioned studies, increased formation of pro-inflammatory mediators as a consequence of the enhanced activity of TLR4/MyD88-dependent TRAF6/IKKα/β/γ/IκB-α and TRAF6/MEK1/2/ERK1/2/AP-1 signaling pathways associated with down-regulation of anti-apoptotic PI3K p85α/Akt1/CREB1/Bcl-2 signaling pathway in addition to reduced expression of anti-inflammatory PPARα/β/γ proteins in the CNS of mice seems to be involved in the LPS-induced inflammatory hyperalgesia.
Bexarotene, an RXR agonist, has been shown to reduce COX-2 expression and/or activity in various malignant cells and is currently used at high doses in the treatment of cutaneous T cell lymphoma with its approved indication. In recent years, it has been reported that bexarotene, which has also been shown to be effective in many acute and chronic CNS diseases associated with inflammation, may be useful in the treatment of neurodegeneration-related disorders, such as traumatic brain injury, in which inflammation plays an important role when used at doses 100 times lower than rodent cancer models4-6. In a study of intestinal neoplasia, bexarotene was shown to diminish COX-2 messenger ribonucleic acid expression and the expression of COX-2-induced pro-inflammatory cytokines such as IL-1β, IL-2, IL-6 and TNF-α53. In an in vitro study, it has been shown that bexarotene suppresses the activity of NF-kB at the transcriptional level by reducing the transactivation potential of the binding site of p65 without affecting the translocation of NF-κB, a transcription factor that causes of enhanced expression of pro-inflammatory cytokines in thyroid cancer cells and the binding ability of one of its subunits, p6554. In another study, it has been suggested that bexarotene inhibits phosphorylation of IKK/I κB-α in Zucker diabetic rat hearts and mediates NF-κB to remain bound to IκB-α in the cytosol, thus preventing its transcriptional activity from being transferred to the nucleus55. In addition, there are studies showing that enhanced NF-κB transcriptional activity in various pathologies is similarly reduced with different RXR ligands56,57. Yuan et al.58 also showed that bexarotene ameliorates depression-like behavior in mice by protecting against neuroinflammation and synaptic injury mediated by the LPS- or corticosterone-induced CREB/brain-derived neurotrophic factor/ERK pathway. Moreover, the results of our study, in which we performed a septic shock model with LPS in rats, show that bexarotene has an inhibitory effect on hypotension accompanied by inflammation and tissue damage during endotoxemia, as well as enhanced expression and/or activity of CYP4F6, which is responsible for leukotriene B4 synthesis, with heterodimer formation of PPARα/β/γ in addition to the reduction in iNOS expression/levels14. There are also studies showing that bexarotene can prevent brain injury and neuroinflammation by increasing the efficiency of the PPARγ/sirtuin 6/forkhead box O3a pathway in subarachnoid hemorrhage models in mice and rats59,60. In the present study, treatment with bexarotene exhibited a marked improvement in the latency compared with endotoxemic mice. Furthermore, bexarotene prevented the LPS-induced (1) Increase in the expression of TRAF6 and c-Jun proteins in addition to phosphorylation of IKKα/β/γ, IkB-α, MEK1/2, ERK1/2 and c-Jun proteins and (2) Decrease in the expression of IkB-α, Bcl-2, PPARα/β/γ proteins as well as phosphorylation of PI3K p85α, Akt1 and CREB1 in the tissues. Based on the results of our previous findings on the inflammatory hyperalgesia model induced by LPS injection8-11 and the above-mentioned studies in the literature, reduced pro-inflammatory mediator formation as a result of suppression of TLR4/MyD88-dependent TRAF6/IKKα/β/γ/IκB-α and TRAF6/MEK1/2/ERK1/2/AP-1 signaling pathways associated with up-regulation of anti-apoptotic PI3K p85α/Akt1/CREB1/Bcl-2 signaling pathway as well as anti-inflammatory PPARα/β/γ proteins at transcriptional and/or post-transcriptional level in the CNS of mice seems to be involved in the analgesic and anti-inflammatory effects of bexarotene.
Although the molecular mechanisms of its beneficial effects on inflammatory hyperalgesia and apoptosis could not be investigated in detail within the scope of this study, it has been suggested that bexarotene exerts its anti-inflammatory and anti-apoptotic effects either directly, through activation of endogenous PPARα/β/γ-RXR-mediated pathways or indirectly, as a result of increasing the expression and/or activity of PI3K p85α/Akt1/CREB1/Bcl-2 signaling pathway and/or suppression of the expression/activity of various factors/enzymes involved in the TRAF6/IKKα/β/γ/IκB-α and TRAF6/MEK1/2/ERK1/2/AP-1 signaling pathways. In addition, bexarotene may also exert its beneficial effects due to its synergistic effect on the signaling pathways that play a role in the pathogenesis of inflammatory hyperalgesia caused by LPS. However, additional studies should be done to prove the validity of these hypotheses.
CONCLUSION
This study provides evidence for the first time that RXR agonist bexarotene can prevent inflammatory hyperalgesia and increase in the activity of the LPS-induced pro-inflammatory TLR4/MyD88-dependent TRAF6/IKKα/ β/γ/IκB-α and TRAF6/MEK1/2/ERK1/2/AP-1 signaling pathways associated with the decrease in the activity of anti-apoptotic PI3K p85α/Akt1/CREB1/Bcl-2 signaling pathway in addition to anti-inflammatory PPARα/β/γ proteins in the CNS of mice. Current findings showed that RXR agonists such as bexarotene, which can also enter the CNS when administered systemically, can be useful as anti-inflammatory/analgesic drugs in the prevention and treatment of acute neuroinflammatory diseases accompanied by pain, as well as chronic painful conditions in which inflammation plays a role in the pathophysiology that may develop due to bacterial infections.
SIGNIFICANCE STATEMENT
S
This study explored the ameliorating effect of bexarotene on the TLR4/MyD88-dependent pro-inflammatory, anti-apoptotic and anti-inflammatory signaling pathways that can be beneficial for treatment of inflammatory hyperalgesia. This study will help the researchers to uncover the critical area of controlling of inflammatory pain during endotoxemia that many researchers were not able to explore. It is expected that the results obtained from this study will contribute to the pre-clinical and clinical studies to be conducted to develop RXR agonists as drugs for the treatment of inflammatory neurodegenerative diseases accompanied by hyperalgesia.
ACKNOWLEDGMENT
This study was supported by the Research Fund of Mersin University in Turkey with Project Number 2021-1-TP2-4299. The findings of this work were included in the Master’s Thesis of Pharm. M.S. Omer Bahceli.
REFERENCES
- de Almeida, N.R., M. Conda-Sheridan, 2019. A review of the molecular design and biological activities of RXR agonists. Med. Res. Rev., 39: 1372-1397.
CrossRefDirect Link - Brtko, J. and Z. Dvorak, 2020. Natural and synthetic retinoid X receptor ligands and their role in selected nuclear receptor action. Biochimie, 179: 157-168.
CrossRefDirect Link - Leal, A.S., L.A. Reich, J.A. Moerland, D. Zhang and K.T. Liby, 2021. Potential Therapeutic Uses of Rexinoids. In: Advances in Pharmacology, Leal, A.S., L.A. Reich, J.A. Moerland, D. Zhang and K.T. Liby (Eds.), Elsevier, Netherlands, pp: 141-183.
CrossRefDirect Link - Gui, Y., S. Duan, L. Xiao, J. Tang and A. Li, 2020. Bexarotent attenuated chronic constriction injury-induced spinal neuroinflammation and neuropathic pain by targeting mitogen-activated protein kinase phosphatase-1. J. Pain., 21: 1149-1159.
CrossRefDirect Link - McFarland, K., T.A. Spalding, D. Hubbard, J.N. Ma, R. Olsson and E.S. Burstein, 2013. Low dose bexarotene treatment rescues dopamine neurons and restores behavioral function in models of Parkinson's disease. ACS Chem. Neurosci., 4: 1430-1438.
CrossRefDirect Link - Zhong, J., C. Cheng, H. Liu, Z. Huang and Y. Wu et al., 2017. Bexarotene protects against traumatic brain injury in mice partially through apolipoprotein E. Neuroscience, 343: 434-448.
CrossRefDirect Link - Deuis, J.R., L.S. Dvorakova and I. Vetter, 2017. Methods used to evaluate pain behaviors in rodents. Front. Mol. Neurosci., Vol. 10.
CrossRefDirect Link - Tunctan, B., E. Ozveren, B. Korkmaz, C.K. Buharalioglu, L. Tamer, U. Degirmenci and U. Atik, 2006. Nitric oxide reverses endotoxin-induced inflammatory hyperalgesia via inhibition of prostacyclin production in mice. Pharmacol. Res., 53: 177-192.
CrossRefDirect Link - Buharalioglu, K., O. Ozbasoglu, B. Korkmaz, T. Cuez, S. Sahan-Firat, A. Yalçın and B. Tunctan, 2009. Thalidomide potentiates analgesic effect of COX inhibitors on endotoxin-induced hyperalgesia by modulating TNF-{alpha}, PGE and NO synthesis in mice. FASEB J., 23: 742.4-742.4.
CrossRefDirect Link - Dolunay, A., S.P. Senol, M. Temiz-Resitoglu, D.S. Guden, A.N. Sari, S. Sahan-Firat and B. Tunctan, 2017. Inhibition of NLRP3 inflammasome prevents LPS-induced inflammatory hyperalgesia in mice: Contribution of NF-κB, caspase-1/11, ASC, NOX, and NOS isoforms. Inflammation, 40: 366-386.
Direct Link - Senol, S.P., M. Temiz-Resitoglu, D.S. Guden, A.N. Sari, S. Sahan-Firat and B. Tunctan, 2021. Suppression of TLR4/MyD88/TAK1/NF-κB/COX-2 signaling pathway in the central nervous system by bexarotene, a selective RXR agonist, prevents hyperalgesia in the lipopolysaccharide-induced pain mouse model. Neurochem. Res., 46: 624-637.
CrossRefDirect Link - Cagli, A., S.P. Senol, M. Temiz-Resitoglu, D.S. Guden, A.N. Sari, S. Sahan-Firat and B. Tunctan, 2021. Soluble epoxide hydrolase inhibitor trifluoromethoxyphenyl-3-(1-propionylpiperidin-4-yl)urea prevents hyperalgesia through regulating NLRC4 inflammasome-related pro-inflammatory and anti-inflammatory signaling pathways in the lipopolysaccharide-induced pain mouse model. Drug Dev. Res., 82: 815-825.
CrossRefDirect Link - Tunctan, B., B. Korkmaz, A.N. Sari, M. Kacan and D. Unsal et al., 2013. 5, 14-HEDGE, a 20-HETE mimetic, reverses hypotension and improves survival in a rodent model of septic shock: Contribution of soluble epoxide hydrolase, CYP2C23, MEK1/ERK1/2/IKKβ/IκB-α/NF-κB pathway and proinflammatory cytokine formation. Prostaglandins Other Lipid Mediat., 102-103: 31-41.
CrossRefPubMedDirect Link - Tunctan, B., S.P. Kucukkavruk, M. Temiz-Resitoglu, D.S. Guden, A.N. Sari and S. Sahan-Firat , 2018. Bexarotene, a selective RXRα agonist, aeverses hypotension associated with inflammation and tissue injury in a rat model of septic shock. Inflammation, 41: 337-355.
CrossRefDirect Link - Festing, M.F.W., 2018. On determining sample size in experiments involving laboratory animals. Lab. Anim., Vol. 52.
CrossRefDirect Link - Ciesielska, A., M. Matyjek and K. Kwiatkowska, 2021. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci., 78: 1233-1261.
CrossRefDirect Link - Shi, J.H. and S.C. Sun, 2018. Tumor necrosis factor receptor-associated factor regulation of nuclear factor κB and mitogen-activated protein kinase pathways. Front. Immunol., Vol. 9.
CrossRefDirect Link - Cianciulli, A., C. Porro, R. Calvello, T. Trotta, D.D. Lofrumento and M.A. Panaro, 2020. Microglia mediated neuroinflammation: Focus on PI3K modulation. Biomolecules, Vol. 10.
CrossRefDirect Link - Yang, Y., J. Lv, S. Jiang, Z. Ma and D. Wang et al., 2016. The emerging role of toll-like receptor 4 in myocardial inflammation. Cell Death Dis., Vol. 7.
CrossRefDirect Link - Strosznajder, A.K., S. Wójtowicz, M.J. Jeżyna, G.Y. Sun and J.B. Strosznajder, 2021. Recent insights on the role of PPAR-β/δ in neuroinflammation and neurodegeneration, and its potential target for therapy. Neuromol. Med. 23: 86-98.
CrossRefDirect Link - Wójtowicz, S., A.K. Strosznajder, M. Jeżyna and J.B. Strosznajder, 2020. The novel role of PPAR alpha in the brain: Promising target in therapy of Alzheimer's disease and other neurodegenerative disorders. Neurochem. Res., 45: 972-988.
CrossRefDirect Link - Calil, I.L., A.C. Zarpelon, A.T.G. Guerrero, J.C.A. Filho and S.H. Ferreira et al., 2014. Lipopolysaccharide induces inflammatory hyperalgesia triggering a TLR4/MyD88-dependent cytokine cascade in the mice paw. PLoS One, Vol. 9.
CrossRefDirect Link - Qi, S., F. Zhao, Z. Li, F. Liang and S. Yu, 2020. Silencing of PTX3 alleviates LPS-induced inflammatory pain by regulating TLR4/NF-κB signaling pathway in mice. Biosci. Rep., Vol. 40.
CrossRefDirect Link - Zucoloto, A.Z., M.F. Manchope, L. Staurengo-Ferrari, F.A. Pinho-Ribeiro and A.C. Zarpelon et al., 2017. Probucol attenuates lipopolysaccharide-induced leukocyte recruitment and inflammatory hyperalgesia: Effect on NF-кB activation and cytokine production. Eur. J. Pharmacol., 809: 52-63.
CrossRefDirect Link - Chae, B.S., 2018. Pretreatment of low-dose and super-low-dose LPS on the production of in vitro LPS-induced inflammatory mediators. Toxicol. Res., 34: 65-73.
CrossRefDirect Link - Choi, M.J., E.J. Lee, J.S. Park, S.N. Kim, E.M. Park and H.S. Kim, 2017. Anti-inflammatory mechanism of galangin in lipopolysaccharide-stimulated microglia: Critical role of PPAR-γ signaling pathway. Biochem. Pharmacol., 144: 120-131.
CrossRefDirect Link - Khan, M.S., T. Ali, M.W. Kim, M.H. Jo, J.I. Chung and M.O. Kim, 2019. Anthocyanins improve hippocampus-dependent memory function and prevent neurodegeneration via JNK/Akt/GSK3β signaling in LPS-treated adult mice. Mol. Neurobiol., 56: 671-687.
CrossRefDirect Link - Zou, Z.Q., J.J. Chen, H.F. Feng, Y.F. Cheng and H.T. Wang et al., 2017. Novel phosphodiesterase 4 inhibitor FCPR03 alleviates lipopolysaccharide-induced neuroinflammation by regulation of the cAMP/PKA/CREB signaling pathway and NF-κB inhibition. J. Pharmacol. Exp. Ther., 362: 67-77.
CrossRefDirect Link - Lee, Y.H., C.H. Lin, P.C. Hsu, Y.Y. Sun and Y.J. Huang et al., 2015. Aryl hydrocarbon receptor mediates both proinflammatory and anti-inflammatory effects in lipopolysaccharide-activated microglia. Glia, 63: 1138-1154.
CrossRefDirect Link - Zhang, L., Z. Zhang, Y. Fu, P. Yang, Z. Qin, Y. Chen and Y. Xu, 2016. Trans-cinnamaldehyde improves memory impairment by blocking microglial activation through the destabilization of iNOS mRNA in mice challenged with lipopolysaccharide. Neuropharmacology, 110: 503-518.
CrossRefDirect Link - Jung, J.S., M.J. Choi, Y.Y. Lee, B.I. Moon, J.S. Park and H.S. Kim, 2017. Suppression of lipopolysaccharide-induced neuroinflammation by Morin via MAPK, PI3K/Akt, and PKA/HO-1 signaling pathway modulation. J. Agric. Food Chem., 65: 373-382.
CrossRefDirect Link - Lee, E.J., H.M. Ko, Y.H. Jeong, E.M. Park and H.S. Kim, 2015. β-Lapachone suppresses neuroinflammation by modulating the expression of cytokines and matrix metalloproteinases in activated microglia. J. Neuroinflammation, Vol. 12.
CrossRefDirect Link - Qi, G., Y. Mi, R. Fan, R. Li, Z. Liu and X. Liu, 2019. Nobiletin protects against systemic inflammation-stimulated memory impairment via MAPK and NF-κB signaling pathways. J. Agric. Food Chem., 67: 5122-5134.
CrossRefDirect Link - Hui, B., L. Zhang, Q. Zhou and L. Hui, 2018. Pristimerin inhibits LPS-triggered neurotoxicity in BV-2 microglia cells through modulating IRAK1/TRAF6/TAK1-mediated NF-κB and AP-1 signaling pathways in vitro. Neurotox. Res., 33: 268-283.
CrossRefDirect Link - Jung, Y.S., J.H. Park, H. Kim, S.Y. Kim and J.Y. Hwang et al., 2016. Probucol inhibits LPS-induced microglia activation and ameliorates brain ischemic injury in normal and hyperlipidemic mice. Acta Pharmacol. Sin., 37: 1031-1044.
CrossRefDirect Link - Yan, J., J. Bai, C. Gao, Y. Liang, B. Zhao and Y. Bian, 2017. Chronic unpredictable stress abrogates the endotoxin tolerance induced by repeated peripheral LPS challenge via the TLR4 signaling pathway. Neurosci. Lett., 645: 7-13.
CrossRefDirect Link - Maitra, U., H. Deng, T. Glaros, B. Baker, D.G.S. Capelluto, Z. Li and L. Li, 2012. Molecular mechanisms responsible for the selective and low-grade induction of proinflammatory mediators in murine macrophages by lipopolysaccharide. J. Immunol., 189: 1014-1023.
CrossRefDirect Link - Qin, L., X. Wu, M.L. Block, Y. Liu and G.R. Breese et al., 2007. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia, 55: 453-462.
CrossRefPubMedDirect Link - Xu, J.L.B., Z. Chen, C. Zhou, L. Liao and Y. Qin et al., 2018. PI3K/AKT/JNK/p38 signalling pathway-mediated neural apoptosis in the prefrontal cortex of mice is involved in the antidepressant-like effect of pioglitazone. Clin. Exp. Pharmacol. Physiol., 45: 525-535.
CrossRefDirect Link - Badshah, H., M. Ikram, W. Ali, S. Ahmad, J.R. Hahm and M.O. Kim, 2019. Caffeine may abrogate LPS-induced oxidative stress and neuroinflammation by regulating Nrf2/TLR4 in adult mouse brains. Biomolecules, Vol. 9.
CrossRefDirect Link - Yang, L., R. Zhou, Y. Tong, P. Chen, Y. Shen, S. Miao and X. Liu, 2020. Neuroprotection by dihydrotestosterone in LPS-induced neuroinflammation. Neurobiol. Dis., Vol. 140.
CrossRefDirect Link - Zong, M.M., H.M. Yuan, X. He, Z.Q. Zhou, X.D. Qiu, J.J. Yang and M.H. Ji, 2019. Disruption of striatal-enriched protein tyrosine phosphatase signaling might contribute to memory impairment in a mouse model of sepsis-associated encephalopathy. Neurochem. Res., 44: 2832-2842.
CrossRefDirect Link - Reinert, K.R.S., C.D. Umphlet, A. Quattlebaum and H.A. Boger, 2014. Short-term effects of an endotoxin on substantia nigra dopamine neurons. Brain Res., 1557: 164-170.
CrossRefDirect Link - Zhang, S., X. Wang, S. Ai, W. Ouyang, Y. Le and J. Tong, 2017. Sepsis-induced selective loss of NMDA receptors modulates hippocampal neuropathology in surviving septic mice. PLoS ONE, Vol. 12.
CrossRefDirect Link - Sui, H., M. Luo, Y. Miao, W. Cheng and S. Wen et al., 2019. Cystic fibrosis transmembrane conductance regulator ameliorates lipopolysaccharide-induced acute lung injury by inhibiting autophagy through PI3K/AKT/mTOR pathway in mice. Respir. Physiol. Neurobiol., Vol. 273.
CrossRefDirect Link - Zhong, W., K. Qian, J. Xiong, K. Ma, A. Wang and Y. Zou, 2016. Curcumin alleviates lipopolysaccharide induced sepsis and liver failure by suppression of oxidative stress-related inflammation via PI3K/AKT and NF-κB related signaling. Biomed. Pharmacother., 83: 302-313.
CrossRefDirect Link - Wu, X.T., A.R. Ansari, X.X. Pang, H.Z. Li and Z.W. Zhang et al., 2019. Visfatin plays a significant role in alleviating lipopolysaccharide-induced apoptosis and autophagy through PI3K/AKT signaling pathway during acute lung injury in mice. Arch. Immunol. Ther. Exp., 67: 249-261.
CrossRefDirect Link - Wu, X., W. Xu, G. Cui, Y. Yan and X. Wu et al., 2013. The expression pattern of nischarin after lipopolysaccharides (LPS)-induced neuroinflammation in rats brain cortex. Inflamm. Res., 62: 929-940.
CrossRefDirect Link - Zhuo, X.J., Y. Hao, F. Cao, S.F. Yan and H. Li et al., 2018. Protectin DX increases alveolar fluid clearance in rats with lipopolysaccharide-induced acute lung injury. Exp. Mol. Med., 50: 1-13.
CrossRefDirect Link - Hoogland, I.C.M., C. Houbolt, D.J. van Westerloo, W.A. van Gool and D. van de Beek, 2015. Systemic inflammation and microglial activation: Systematic review of animal experiments. J Neuroinflammation, Vol. 12.
CrossRefDirect Link - Batista, C.R.A., G.F. Gomes, E.C. Jalil, B.L. Fiebich and A.C.P. de Oliveira, 2019. Lipopolysaccharide-induced neuroinflammation as a bridge to understand neurodegeneration. Int. J. Mol. Sci., Vol. 20.
CrossRefDirect Link - Meneses, G., M. Rosetti, A. Espinosa, A. Florentino and M. Bautista et al., 2018. Recovery from an acute systemic and central LPS-inflammation challenge is affected by mouse sex and genetic background. PLoS ONE, Vol.13.
CrossRefDirect Link - Janakiram, N.B., A. Mohammed, L. Qian, C.I. Choi, V.E. Steele and C.V. Rao, 2012. Chemopreventive effects of RXR-selective rexinoid bexarotene on intestinal neoplasia of ApcMin/+ mice. Neoplasia, 14: 159-168.
CrossRefDirect Link - Cras, A., B. Politis, N. Balitrand, D.D. Bettinger and P.Y. Boelle et al., 2012. Bexarotene via CBP/p300 induces suppression of NF-κB-dependent cell growth and invasion in thyroid cancer. Clin. Cancer Res., 18: 442-453.
CrossRefDirect Link - Nizamutdinova, I.T., R.S. Guleria, A.B. Singh, J.A. Kendall Jr., K.M. Baker and J. Pan, 2013. Retinoic acid protects cardiomyocytes from high glucose-induced apoptosis through inhibition of NF-κB signaling pathway. J. Cell Physiol., 228: 380-392.
CrossRefDirect Link - Chen, H., F. Ma, X. Hu, T. Jin, C. Xiong and X. Teng, 2013. Elevated COX2 expression and PGE2 production by downregulation of RXRα in senescent macrophages. Biochem. Biophys. Res. Commun., 440: 157-162.
CrossRefDirect Link - Ning, R.B., J. Zhu, D.J. Chai, C.S. Xu and H. Xie et al., 2013. RXR agonists inhibit high glucose-induced upregulation of inflammation by suppressing activation of the NADPH oxidase-nuclear factor-κB pathway in human endothelial cells. Genet. Mol. Res., 12: 6692-6707.
CrossRefDirect Link - Yuan, C., C. Dai, Z. Li, L. Zheng, M. Zhao, and S. Dong, 2020. Bexarotene improve depression-like behaviour in mice by protecting against neuro-inflammation and synaptic damage. Neurochem. Res., 45: 1500-1509.
CrossRefDirect Link - Tu, L., X.l. Yang, Q. Zhang, Q. Wang and T. Tian et al., 2018. Bexarotene attenuates early brain injury via inhibiting micoglia activation through PPARγ after experimental subarachnoid hemorrhage. Neurol. Res., 40: 702-708.
CrossRefDirect Link - Zuo, Y., L. Huang, B. Enkhjargal, W. Xu and O. Umut et al., 2019. Activation of retinoid X receptor by bexarotene attenuates neuroinflammation via PPARγ/SIRT6/FoxO3a pathway after subarachnoid hemorrhage in rats. J. Neuroinflammation, Vol. 16.
CrossRefDirect Link