
Shanshan Zhang
School of Medicine of Pingdingshan University, Pingdingshan, Henan 467000, China
LiveDNA: 86.37247
Fengjuan Yuan
School of Medicine of Pingdingshan University, Pingdingshan, Henan 467000, China
Xiao Liu
School of Medicine of Pingdingshan University, Pingdingshan, Henan 467000, China
Yali Liu
School of Medicine of Pingdingshan University, Pingdingshan, Henan 467000, China
International Journal of Pharmacology
Year: 2022 | Volume: 18 | Issue: 6 | Page No.: 1161-1170







ABSTRACT
Background and Objective: As known, diabetes mellitus (DM) is a metabolic disorder that is highly correlated with the dysfunction of pancreatic β cells. Herein, the study aimed to investigate the potential role of miR-33-5p as a regulator of pancreatic β cells and the underlying mechanism in DM. Materials and Methods: The RNA and protein expression were analyzed by Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR) and western blot, respectively. The target gene of miR-33-5p was identified and verified by TargetScan, dual-luciferase reporter and RNA immunoprecipitation (RIP) assays. The MTT, TUNEL, as well as ELISA analyses, were performed after streptozotocin (STZ)-induced INS-1 cells were transfected with miR-33-5p mimics/inhibitors and si-RND2 alone and in combination. Results: The miR-33-5p was lowly expressed in islet and serum of DM patients, STZ-induced mice and INS-1 cells. In vitro showed that the suppression of cell proliferation and insulin secretion and the increase of apoptosis induced by STZ was reversed by miR-33-5p overexpression in INS-1 cells, accompanied by the activation ofPI3K/AKT signalling pathway. Mechanically, RND2 was identified as the target of miR-33-5p in DM. Furthermore, the effects of miR-33-5p depletion on the cell proliferation, apoptosis and insulin secretion of STZ-induced INS-1 cells were restrained by RND2 knockdown. Conclusion: Overall, current findings proposed that miR-33-5p/RND2 were promising targets that regulated the balance of insulin secretion.
PDF Abstract XML References Citation
Copyright: © 2022. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Shanshan Zhang, Fengjuan Yuan, Xiao Liu and Yali Liu, 2022. miR-33-5p Ameliorates β Cell Dysfunction and PI3K/AKT Signaling-Mediated Insulin Secretion in Diabetes via Targeting RND2. International Journal of Pharmacology, 18: 1161-1170.
DOI: 10.3923/ijp.2022.1161.1170
URL: https://scialert.net/abstract/?doi=ijp.2022.1161.1170
DOI: 10.3923/ijp.2022.1161.1170
URL: https://scialert.net/abstract/?doi=ijp.2022.1161.1170
INTRODUCTION
Diabetes mellitus (DM) is a metabolic disease caused by insufficient insulin secretion and is mainly classified into type 1 diabetes mellitus (T1DM) and type 2 diabetes mellitus (T2DM)1,2. The occurrence and development of diabetes are primarily related to environmental and genetic factors as well as autoimmune system disorders3,4. Clinically, diabetes is generally accompanied by some complications, such as blindness, renal failure, cardiovascular disease and even amputation5,6. Currently, increasing evidence has demonstrated that pancreatic β cells are related to balanced insulin secretion7 and plasma glucose levels8 and it has been proved that the dysfunction or apoptosis of β cells is highly associated with the onset and development of diabetes9. Hence, it is necessary to investigate the apoptosis and insulin secretion of pancreatic β cells and propose a novel strategy for the treatment of diabetes.
MicroRNAs (miRNAs) have been clarified to modulate messenger RNAs via binding to the region of 3'-UTR10. Recently, the functional role of miRNAs in regulating DM has been validated. For example, Le May C and colleagues have determined the metabolic function of miR-574-5p and miR-3135b in blood glucose levels for gestational diabetes mellitus11 revealed that miR-212-5p carrying exosomes derived from M1 macrophage restrain Akt/GSK-3β/β-catenin signalling pathway via binding to the 3'-UTR of SIRT2, blocking the secretion of β-cell insulin12. In addition, Cheng et al.12. found that miR-296-3p overexpression repaired pancreatic β cell dysfunction induced by UA in Type 2 Diabetes. It was uncovered that microRNA-33-5p (miR-33-5p) was downregulated in DM patients13, while its functional role in the apoptosis and insulin secretion of pancreatic β cells has not been disclosed.
The Rho GTPase 2 (RND2) is a branch of the Rho GTPase family14, which is mainly related to the regulation of actin cytoskeleton organization15 and cell cycle16. In addition, Xu et al.17. verified that RND2 weakened apoptosis and autophagy via the p38 MAPK signalling pathway in glioblastoma. Enormous investigations have validated the effect of RND2 on neurons. For example, Heng et al.18 found that RND2, which was negatively mediated by RP58, regulated migration of neurons in the development of the cerebral cortex and RND2 expression, which was suppressed by COUP-TFI, was negatively associated with the radial migration and the normal morphology of callosal projection neurons19. However, whether RND2 plays a crucial role in pancreatic β cell apoptosis remains elusive.
It is hypothesized that miR-33-5p may target RND2. In this study, expression levels of miR-33-5p and RND2 in diabetes
patients and pancreatic β cells were determined and their correlation was analyzed. Moreover, the role of miR-33-5p and RND2 in β cell apoptosis and insulin secretion was also explored, which may provide novel spots for diabetes treatment.
MATERIALS AND METHODS
Study area: All of the research was performed between March, 2018 and November, 2020 in the School of Medicine of Pingdingshan University.
Serum and tissue collection: Islet tissues and serum samples were collected from the School of Medicine of Pingdingshan University, the Philippine Women’s University. The experiments followed the Declaration of Helsinki and the protocol was approved by the ethics committee of the School of Medicine of Pingdingshan University, the Philippine Women’s University.
Construction of diabetic mice models: For the establishment of diabetic mice, 6-8 weeks aged male C57BL/6 mice (20±2 g) were provided by the School of Medicine of Pingdingshan University, the Philippine Women’s University and were maintained in pathogen-free conditions. 60 mg kg–1 STZ (Sigma, United States) was consecutively injected intraperitoneally into mice for 5 days, while mice in the control group were injected with an equal volume of vehicle. Seven days after injection, the concentration of blood glucose was tested using a glucose meter (Sinocare, China) and successful construction of diabetic mice models was indicated by consecutive concentrations >16.7 mmol L–1 for 3 days. Then the serum was collected for further research.
Cell culture and treatment: The INS-1 cells were purchased from the American Type Culture Collection (ATCC, USA) and were cultured at 37°C in the indicated medium. Then INS-1 cells were treated by 3 mmol L–1 STZ for 12 and 24 hrs, respectively. The miR-33-5p-mimics, miR-33-5p-inhibitors, si-RND2-1 and si-RND2-2 were provided by Gene Chem (Shanghai, China) in Supplementary Table 1.
MTT: Cell proliferation was evaluated using an MTT assay kit (Cinnagen, Iran). In brief, INS-1 cells were maintained in 96-well plates at a density of 2,000 cells/well for 24, 48 and 72 hrs, respectively. After the previous medium was removed, MTT solution (Aladdin, United States) was supplemented. The absorbance was determined at 570 nm using a microplate reader.
TUNEL: To assess apoptosis of INS-1 cells, TUNEL staining was performed. In brief, INS-1 cells were treated and fixed with 4% paraformaldehyde and TUNEL reagent (Merck KGaA, Darmstadt, Germany) was used for staining. The TUNEL-positive cells were tested by optical microscopy (Olympus).
ELISA: For the detection of insulin secretion, an ELISA kit (Beyotime Institute of Biotechnology, Beijing, China) was used as per manufacturer instructions.
Luciferase reporter assay: Luciferase reporter assay was employed to determine the correlation between miR-33-5p and RND2. miR-33-5p mimics with corresponding NC-mimics were co-transfected with RND2-MUT or RND2-WT into INS-1 cells. Using the Dual-Luciferase Reporter Assay System (Promega, Madison, United States), RND2 luciferase activity was evaluated.
RNA immunoprecipitation assay (RIP): For verification of the intersection between miR-33-5p and RND2, RNA immunoprecipitation (RIP) Kit was used as per the manufacturer's instructions. In brief, after lysing INS-1 cells in RIP buffer, the collected lysates were cultured with magnetic beads conjugated with antibodies of Ago2 or IgG (Abcam, UK). The abundance of RND2 was estimated using qRT-PCR.
Western blotting: To determine the protein levels in INS-1 cells, the total proteins were first collected utilizing RIPA lysis buffer. After centrifugation, protein samples from each group were subjected to separation by SDS-PAGE and subsequently transferred onto PVDF membranes, followed by blocking with 5% skimmed milk. Then, the membranes were subjected to overnight incubation with primary antibodies at 4°C before the mixing with chemiluminescent HRP substrate. In this experiment, all primary antibodies were supplied by Abcam (UK), including IRS2 (ab134101,1/5,000), Bcl-2(ab182858,1/2,000), Bax (ab3191,0.417 μg mL–1), Cleaved caspase 3(ab214430,1/5,000), Total caspase 3 (ab13847,1/500), Cleaved caspase 9 (ab2324,1 μg mL–1), Total caspase 9 (ab52298,1/800), p-PI3K (ab278545,0.5 μg mL–1), PI3K (ab32089, 1/1,000), p-AKT(ab38449, 1/700), AKT(ab8805, 1/500) and RND2 (ab200713, 1/1,000). ECL (Amersham, Cytiva) was used to visualize protein bands.
qRT-PCR: For the determination of RNA levels of miR-33-5p, total RNA was collected by 1 mL TRIzol following the guidelines of the RNeasy Mini Kit (ThermoFisher Scientific, USA). Omniscript RT Kit (Qiagen) was used to synthesize cDNAs and SYBR® Premix Ex Taq TM (Takara) Kit was also used. Real-time PCR was performed. GAPDH was considered as the internal parameter. All primer sequences were listed in Supplementary Table 2.
Statistical analysis: GraphPad Prism and SPSS 22.0 software were employed for statistical analysis. Data were displayed as Mean±Standard Deviation (SD). The differences between groups were determined using a t-test or analysis of variance (ANOVA). The p-value<0.05 indicated a significant difference.
RESULTS
miR-33-5p was lowly expressed in STZ-treated pancreatic β cells: To disclose whether miR-33-5p is involved in the regulation of pancreatic β cell function, the expression levels of miR-33-5p in the serum from DM patients (Normal = 25, DM = 20) and STZ-treated diabetic mice were determined and it was found that miR-33-5p was lowly expressed in diabetic serum samples in Fig. 1a, the relative expression of miR-33-5p: 1.057±0.391 vs. 0.499±0.105, p<0.005 in Fig. 1b, 1.057±0.400 vs. 0.538±0.086, p< 0.005. In addition, islet tissues derived from DM patients displayed a higher level of miR-33-5p, in comparison with normal islet tissues in Fig. 1c, the relative expression of miR-33-5p: 1.039±0.306 vs. 0.569±0.124, p<0.005. For further investigation, the expression of miR-33-5p in INS-1 cells were analyzed after treating with STZ (3 mmol L–1) for 12 and 24 hrs, respectively. The qPCR assay indicated that the expression of miR-33-5p in INS-1 cells (1.007±0.142) was significantly restrained after STZ treatment for 12 hrs (0.507±0.231, p<0.05) and 24 hrs (0.312±0.049, p<0.005) in Fig. 1d. Collectively, these findings implied that miR-33-5p was downregulated in STZ-treated pancreatic β cells and may affect the biological function of β cells.
miR-33-5p overexpression induced pancreatic β cell proliferation and insulin secretion: To investigate whether miR-33-5p plays a role in pancreatic β cells, miR-33-5p-overexpressed INS-1 cells were constructed. The result showed that the transfection of miR-33-5p led to a significant increase of miR-33-5p expression the miR-33-5p expression fold changed from 1.006±0.135 to 9.043±2.039, p<0.005 in INS-1 cells in Fig. 2a. The MTT assay elaborated that the cell viability of β cells (100±6.00%) was strikingly decreased to 68.85±9.12% after STZ treatment, which could be efficiently retained by miR-33-5p overexpression (112.33±7.56%) in Fig. 2b.
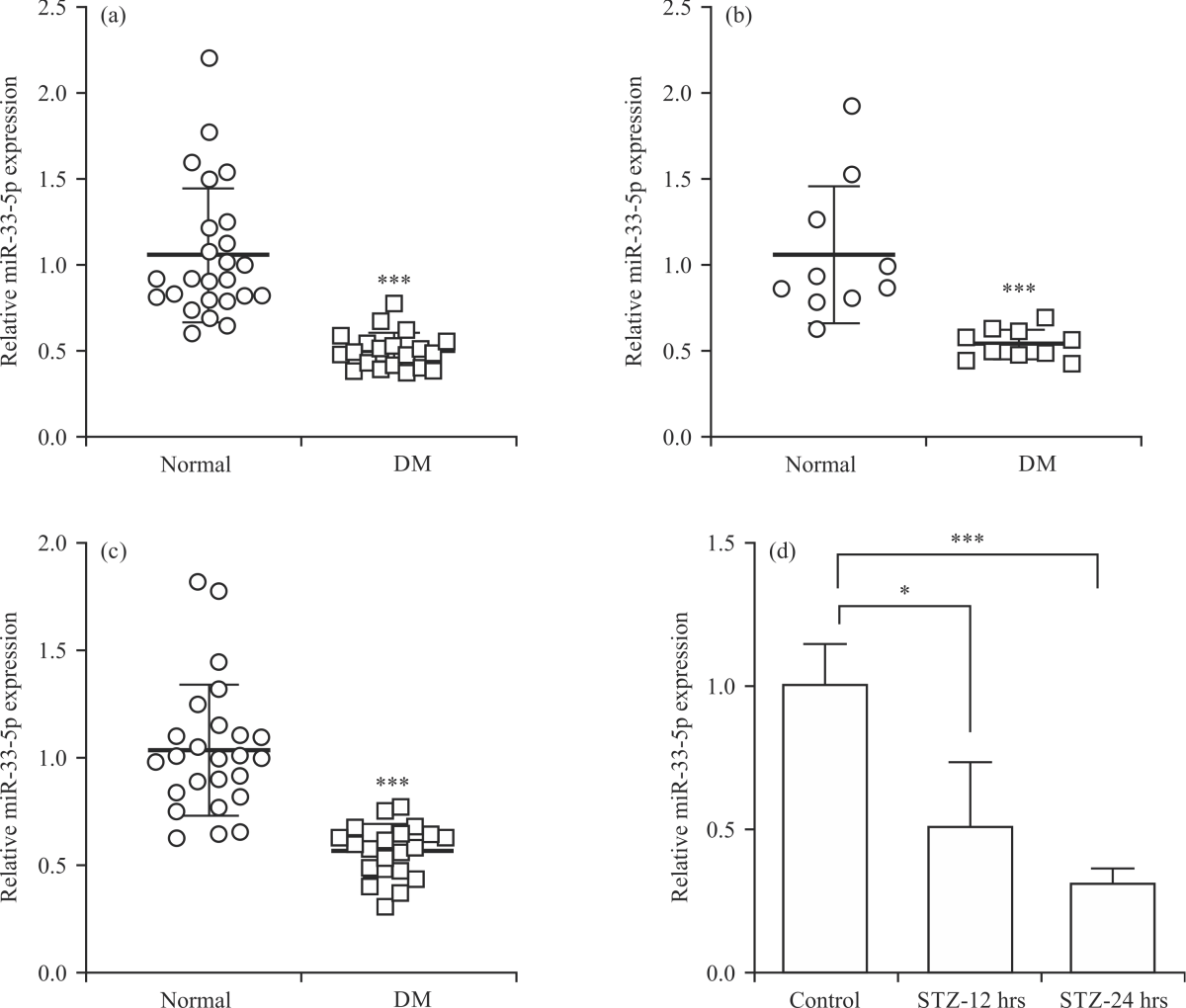 |
| Fig. 1(a-d): | miR-33-5p was lowly expressed in STZ-treated pancreatic β cells, (a) DM patients (Normal = 25, DM = 20), (b) STZ-treated diabetic mice, (c) Islet tissues derived from DM patients (Normal = 25, DM = 20) and (d) STZ-treated INS-1 cells *Statistically significant at p<0.05 and ***p<0.005. The q-PCR assay was utilized to detect the expression levels of miR-33-5p in the serum |
In addition, after STZ treatment, the TUNEL assay showed that the percentage of TUNEL-positive β cells rose from 20.53±1.87-29.68±3.34% (p<0.01), indicating that STZ treatment induced cell apoptosis in Fig. 2c. The enhanced cell apoptosis induced by STZ was blocked by overexpressing miR-33-5p (16.78±1.63%, p< 0.01). Moreover, it was observed that STZ treatment resulted in an obvious drop from 37.78±6.16-25.29±4.53 μU mL–1, p<0.01) in insulin secretion derived from β cells, which was rescued by miR-33-5p overexpression (56.74±6.44 μU mL–1, p<0.01) in Fig. 2d. Mechanistically, the decreased Bcl-2 expression induced by STZ was rescued by miR-33-5p overexpression, while expressions of pro-apoptotic proteins (Bax, Cleaved caspase 3 and Cleaved caspase 9) were inhibited. In addition, miR-33-5p overexpression upregulated insulin receptor substrate-2 (IRS2) and activated PI3K/AKT signalling pathway, indicating that miR-33-5p was involved in insulin secretion in Fig. 2e. Taken together, findings demonstrated that overexpressed miR-33-5p facilitated pancreatic β cell proliferation and induced insulin secretion.
miR-33-5p targeted RND2 in β cells: To unravel the underlying mechanism of miR-33-5p in β cells, the Target Scan Database was used to predict the target genes of miR-33-5p, which were intersected with differentially upregulated genes from the GSE25724 dataset and four intersected genes, including CTBP2, CEND1, RND2 and ERC1, were identified in Fig. 3a. After miR-33-5p was upregulated, Western blotting indicated that only RND2 was significantly downregulated by miR-33-5p overexpression in Fig. 3b. Therefore, RND2 was identified as the targeted gene in further exploration. In addition, qPCR assay demonstrated that RND2 was highly expressed in serum samples (1.028±0.235) derived from diabetic patients, compared with that in normal serum samples (2.213±0.407, p<0.005) in Fig. 3c. Besides, the expression of RND2 (1.003±0.096) was increased by both 12 hrs (1.586±0.232, p<0.05) and 24 hrs (2.888±1.032, p<0.05) STZ treatments in Fig. 3d.
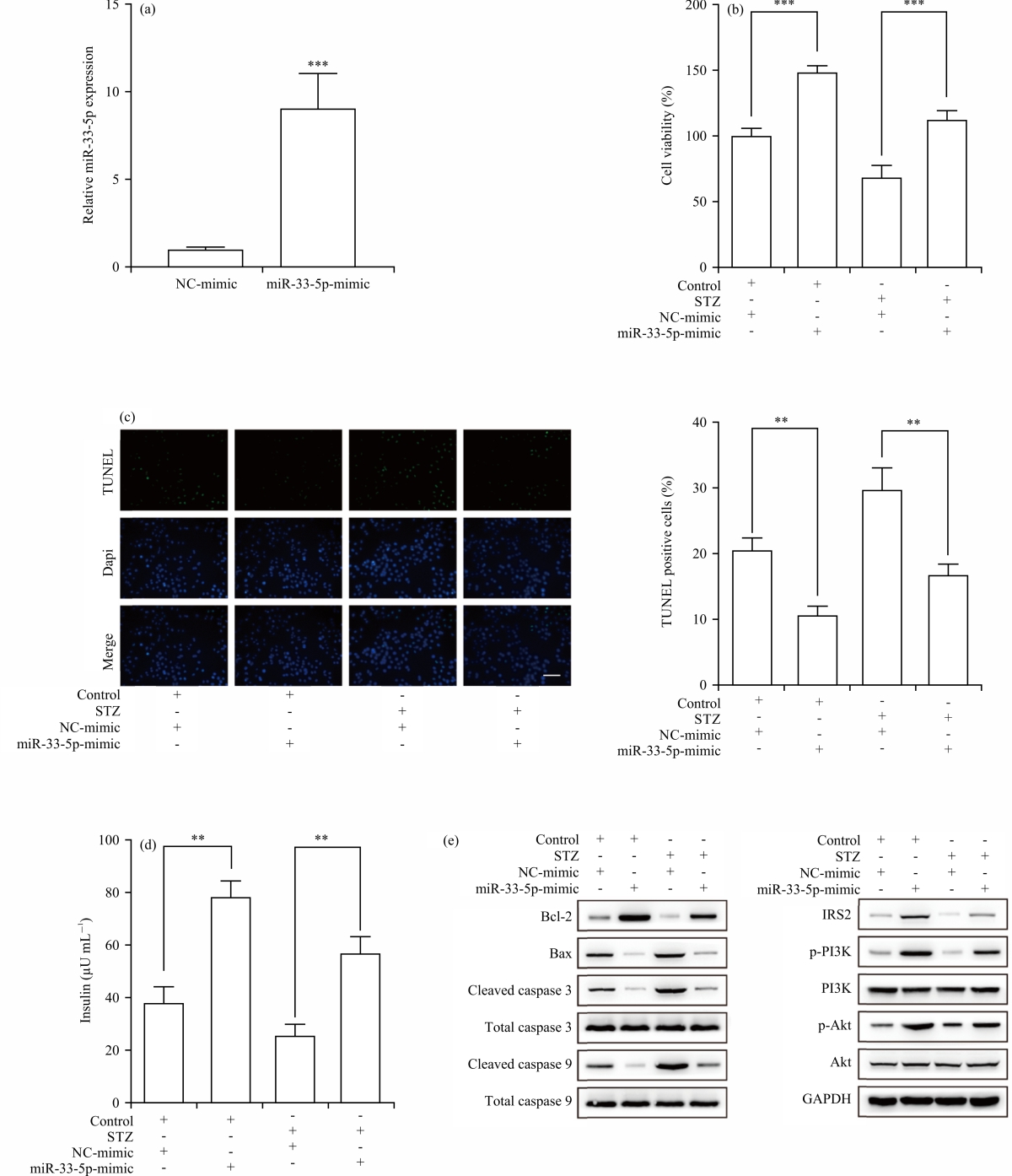 |
| Fig. 2(a-e): | miR-33-5p overexpression induced pancreatic β cell proliferation and insulin secretion, (a) q-PCR, (b) MTT, (c) TUNEL, (d) Insulin secretion and (e) Western blotting **Statistically significant at p<0.01 and ***p<0.005. Pancreatic β cells were transfected with miR-33-5p mimics and the overexpressed efficiency was examined ELISA was utilized to assess apoptosis-related proteins and IRS2/PI3K/AKT signalling pathway |
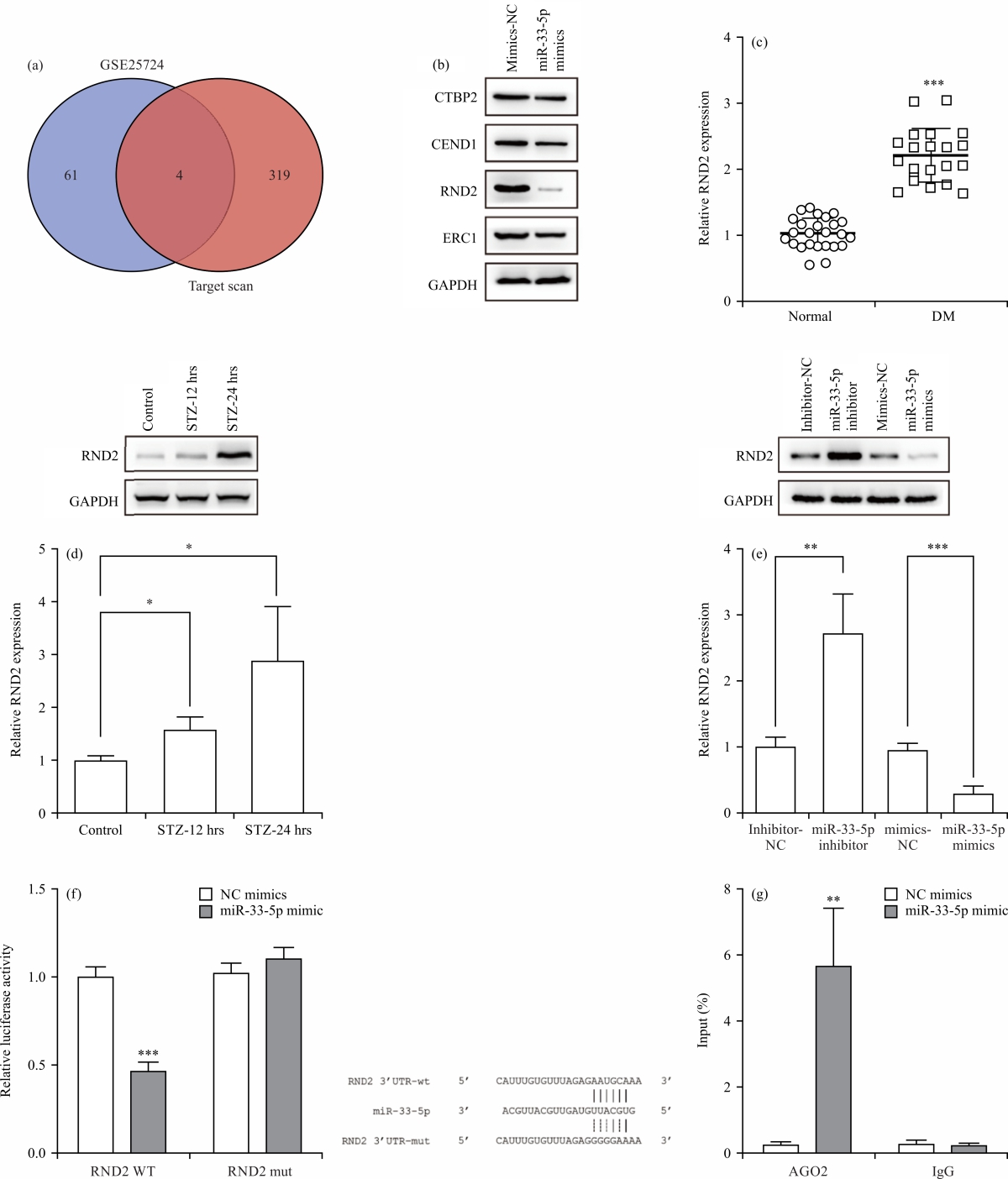 |
| Fig. 3(a-g): | miR-33-5p targeted RND2 in β cells, (a) Target Scan database and GSE25724 dataset, (b) Western blotting, (c) q-PCR and Western blotting in the serum of DM patients (Normal = 25, DM = 20), (d) STZ-treated INS-1 cells, (e) q-PCR and Western blotting in miR-33-5p-downregulated or miR-33-5p-overexpressed INS-1 cells, (f) Dual-luciferase and (g) RIP assays in the correlation of miR-33-5p and RND2 *Statistically significant at p<0.05, **p<0.01 and ***p<0.005 |
To reveal the correlation between miR-33-5p and RND2, miR-33-5p-downregulated and miR-33-5p-overexpressed INS-1 cells were constructed and it was found that miR-33-5p inhibitor upregulated RND2, whereas miR-33-5p mimics produced a reverse effect in Fig. 3e. Furthermore, based on Dual-luciferase reporter assay results, miR-33-5p-mimics considerably weakened the luciferase activity (1.000±0.058-0.466±0.052, p<0.005) in INS-1 cells transfected with RND2 wide-type plasmids, but cells transfected with RND2-MUT plasmids were not affected (1.021±0.059-1.104±0.063, p = 0.219) in Fig. 3f. Compared with the anti-IgG group, the RIP assay validated that miR-33-5p overexpression induced a considerable increase of RND2 in anti-AGO2 complexes in Fig. 3g. In summary, these data showed that miR-33-5p could bind to the 3'-UTR of RND2 in INS-1 cells.
miR-33-5p enhanced β cell proliferation and insulin secretion via targeting RND2: To verify whether miR-33-5p acts as a driver in promoting β cell growth via targeting RND2, miR-33-5p inhibitors and RND2 siRNA were transfected alone or together into STZ-treated INS-1 cells. Western blotting confirmed the transfection of si-RND2was successfully down-regulated the expression of RND2 in Fig. 4a. The MTT assay showed that the cell viability of miR-33-5p silencing β cells (63.46±7.93%) was significantly lower than that of the control (β cells co-transfected with vector and NC inhibitor, 100±7.58%), which was rescued by RND2 depletion (104.91±7.13%, p<0.01) in Fig. 4b. The TUNEL assay clarified that down-regulated miR-33-5p caused an increase in the number of TUNEL-positive cells (from 20.79±2.73-27.86±1.26%), while it was rescued by RND2 knockdown (17.88±2.60, p<0.01) in Fig. 4c. In addition, RND2 knockdown also played a crucial role in recovering the decreased insulin secretion from β cells (from 33.07±3.00-55.25±4.52 μU mL–1, p<0.01) in Fig. 4d. Furthermore, RND2 depletion upregulated Bcl-2, IRS2, p-PI3K and p-AKT in INS-1 cells transfected with miR-33-5p inhibitors and downregulated Bax, Cleaved caspase 3 and Cleaved caspase 9 in Fig. 4e. Overall, these findings confirmed that miR-33-5p promoted β cell proliferation and insulin secretion in an RND2-dependent manner.
DISCUSSION
Insufficient insulin secretion is involved in the onset and progression of DM and pancreatic β cells mainly mediate insulin secretion and balance the glucose levels20. Accumulating evidence has proved that the dysfunction or apoptosis of pancreatic β cells is highly associated with DM. For example, Takahashi et al.21 proved that there was a striking drop in Islet microangiopathy and β-cells in Japanese T2DM patients and Mezza et al.22 revealed the crucial roles of dysfunctional β cells in the diagnosis of diabetes in the pancreaticoduodenectomy model. However, biomarkers uncovered in regulating biological functions of pancreatic β cells remain inadequate.
In this study, miR-33-5p was identified as a driver for pancreatic β cell proliferation and insulin secretion and RND2 was determined as the targeting gene of miR-33-5p. According to previous reports, miRNAs have been proved as crucial mediators of the viability and insulin secretion of pancreatic β cells. Cheng et al.23. demonstrated that miR-181a-5p sponged by lncRNA PVT1 could recover the pancreatic β cell injury caused by streptozotocin and strengthen the capacity of insulin secretion. Ma et al.24. reported that circRNA LRP6 functionally acted as a sponger of miR-9-5p and upregulated PRMT1, thereby aggravating STZ-induced β-cell apoptosis and hindering insulin secretion. In addition, Sun et al.25 found that miR-409-3p acted as an inhibitor in β cell apoptosis and as an inducer in insulin secretion. In a previous report, a reduction of miR-33-5p levels has been observed in the urine of both DM patients and DB/DB mice14. This study examined the expression levels of miR-33-5p in islet and serum of diabetic patients, STZ-treated pancreatic β cells and diabetic mice and it was found that miR-33-5p was lowly expressed in all diabetic samples and inhibited after STZ treatment. In addition, cell experiments validated that miR-33-5p overexpression ameliorated STZ-induced pancreatic β cell dysfunction, which was specifically involved in the recovery of pancreatic β cell viability and promoted cell apoptosis and also enhanced insulin secretion. Noticeably, the fundamental mechanism involved in β cell apoptosis and insulin secretion was investigated and it was found that miR-33-5p overexpression decreased Bcl-2 levels, while enhanced expressions of pro-apoptotic proteins, including Bax, Cleaved caspase 3 and Cleaved caspase 9. Furthermore, miR-33-5p overexpression upregulated IRS2 and activated PI3K/AKT signalling pathway and IRS2/AKT signalling has been proved to play a critical role in mediating pancreatic β cell dysfunction and insulin secretion26.
It is widely believed that miRNAs play a functional role via binding to the 3'-UTR of messengers13. Herein, RND2 was considered as the downstream gene of miR-33-5p based on bioinformatics online analysis and it was observed that RND2 was highly expressed in STZ-treated pancreatic β cells in a time-dependent manner.
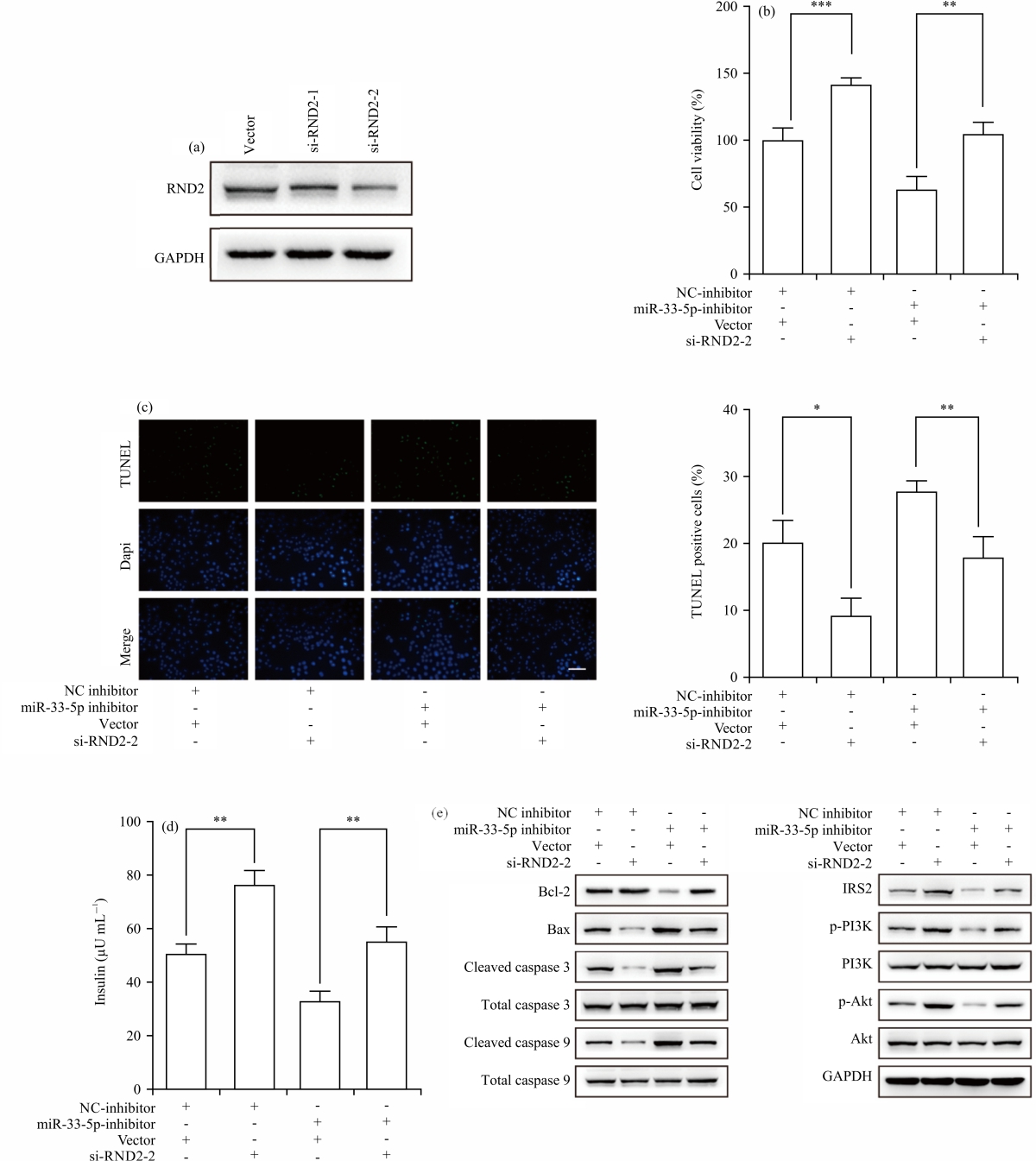 |
| Fig. 4(a-e): | miR-33-5p blocked β cell proliferation and insulin secretion via targeting RND2, (a) Western blotting in RND2 knockdown efficiency, (b) MTT, (c) TUNEL, (d) ELISA and (e) Western blotting in apoptosis-related proteins and IRS2/PI3K/AKT signalling pathway To verify whether miR-33-5p regulates β cell functions through targeting RND2, INS-1 cells treated by STZ for 24 hrs were co-transfected with miR-33-5p inhibitors and RND2 siRNA, *Statistically significant at p<0.05 and **p<0.01 |
Moreover, it was revealed that RND2 depletion rescued the impairment of cell viability and the capacity of insulin secretion caused by miR-33-5p inhibition. However, it should be acknowledged that some limitations exist in this study. First, these findings regarding miR-33-5p/RND2 axis were merely explored by in vitro experiments, further in vivo confirmation was required. Secondly, the molecular mechanism by which the miR-33-5p/RND2 axis regulates PI3K/AKT signalling pathway in DM needs further work.
CONCLUSION
Collectively, it was disclosed that miR-33-5p was inhibited after pancreatic β cells were treated by STZ and miR-33-5p overexpression promoted recovery of STZ-induced pancreatic β cell dysfunction. In addition, as a target gene of miR-33-5p, RND2 depletion played a crucial role in rescuing the miR-33-5p inhibition-induced injury of β cell viability and insulin secretion. These findings demonstrated that targeting the miR-33-5p/RND2 axis is a promising therapeutic strategy for ameliorating DM.
SIGNIFICANCE STATEMENT
This study discovered that miR-33-5p restoration in pancreatic β cells was effective in reversing STZ-induced pancreatic β cell dysfunction and preliminarily uncovered the underlying mechanism, which can be beneficial for clinical research on diabetes mellitus. This study will help the researchers to uncover the critical areas of the regulatory mechanism behind miR-33-5p in diabetes mellitus that many researchers were not able to explore. Thus a new theory on the treatment of diabetes mellitus may be arrived at.
ACKNOWLEDGMENT
This work was supported by The Medical Education Research Project of Henan Province, based on the construction of the elderly nursing technology curriculum system based on the post competency training of elderly nursing talents, project number (No.wjlx2020293) and Pingdingshan University Health Assessment School-level Excellent Online Open Course (No. jpzx19247).
| Supplementary Table 1: | Sequences of siRNA against specific targets | |
| si-RND2-1 | 5'-3' | AGAGCAGACCGACAUAUAAUU |
| si-RND2-2 | 5'-3' | ACAUGUUCCUUCAGAGAUAAG |
| Supplementary Table 2: | Sequences of PCR primers used in this study | |
| miR-33-5p | Forward(5'-3') | ACACTCCAGCTGGGGTGCATTGTAGTT |
| Reverse(5'-3') | TGGTGTCGTGGAGTCG | |
| GAPDH | Forward(5'-3') | CCTCTGACTTCAACAGCGAC |
| Reverse(5'-3') | TCCTCTTGTGCTCTTGCTGG | |
| RND2 | Forward(5'-3') | ACACTGCGAGCTTTGAGATCG |
| Reverse(5'-3') | AGAGGCCGGACATTATCATAGT |
REFERENCES
- Silverii, G.A., C.D. Poggi, I. Dicembrini, M. Monami and E. Mannucci, 2021. Glucose control in diabetes during home confinement for the first pandemic wave of COVID-19: A meta-analysis of observational studies. Acta Diabetol., 58: 1603-1611.
CrossRefDirect Link - Gomes, J.M.G., J. de Assis Costa and R. de Cássia Gonçalves Alfenas, 2017. Metabolic endotoxemia and diabetes mellitus: A systematic review. Metabolism, 68: 133-144.
CrossRefDirect Link - Baynest, H.W., 2015. Classification, pathophysiology, diagnosis and management of diabetes mellitus. J. Diabetes Metab., Vol. 6.
CrossRefDirect Link - Atkinson, M.A., B.O. Roep, A. Posgai, D.C.S. Wheeler and M. Peakman, 2019. The challenge of modulating β-cell autoimmunity in type 1 diabetes. Lancet Diabetes Endocrinol., 7: 52-64.
CrossRefDirect Link - Hernandez, A.F., J.B. Green, S. Janmohamed, R.B. D'Agostino Sr and C.B. Granger et al., 2018. Albiglutide and cardiovascular outcomes in patients with type 2 diabetes and cardiovascular disease (harmony outcomes): A double-blind, randomised placebo-controlled trial. Lancet, 392: 1519-1529.
CrossRefDirect Link - Braunwald, E., 2019. Diabetes, heart failure, and renal dysfunction: The vicious circles. Prog. Cardiovasc. Dis., 62: 298-302.
CrossRefDirect Link - Itoh, Y., Y. Kawamata, M. Harada, M. Kobayashi and R. Fujii et al., 2003. Free fatty acids regulate insulin secretion from pancreatic β cells through GPR40. Nature, 422: 173-176.
CrossRefDirect Link - Schuit, F.C., P. Huypens, H. Heimberg and D.G. Pipeleers, 2001. Glucose sensing in pancreatic β-cells: A model for the study of other glucose-regulated cells in gut, pancreas, and hypothalamus. Diabetes, 50: 1-11.
CrossRefDirect Link - Zhang, Y., Z. Wang and R.A. Gemeinhart, 2013. Progress in microRNA delivery. J. Controlled Release, 172: 962-974.
CrossRefDirect Link - Wang, F., Z. Li, M. Zhao, W. Ye and H. Wu et al., 2021. Circulating miRNAs miR-574-5p and miR-3135b are potential metabolic regulators for serum lipids and blood glucose in gestational diabetes mellitus. Gynecological Endocrinol., 37: 665-671.
CrossRefDirect Link - Qian, B., Y. Yang, N. Tang, J. Wang and P. Sun et al., 2021. M1 macrophage-derived exosomes impair beta cell insulin secretion via miR-212-5p by targeting SIRT2 and inhibiting Akt/GSK-3β/β-catenin pathway in mice. Diabetologia, 64: 2037-2051.
CrossRefDirect Link - Cheng, M., Y. Guo, W. Zhong, X. Chen and G. Guo, 2022. Abnormal expression of MicroRNA-296-3p in type 2 diabetes patients and its role in pancreatic β-cells function by targeting tensin homolog deleted on chromosome ten. Biochem. Genet., 60: 39-53.
CrossRefDirect Link - Tsai, Y.C., P.L. Kuo, W.W. Hung, L.Y. Wu and P.H. Wu et al., 2018. Angpt2 induces mesangial cell apoptosis through the MicroRNA-33-5p-SOCS5 loop in diabetic nephropathy. Mol. Ther. Nucleic Acids, 13: 543-555.
CrossRefDirect Link - Miyamoto, Y., T. Torii, M. Terao, S. Takada, A. Tanoue, H. Katoh and J. Yamauchi, 2021. RND2 differentially regulates oligodendrocyte myelination at different developmental periods. Mol. Biol. Cell, 32: 769-787.
CrossRefDirect Link - Liaci, C., M. Camera, G. Caslini, S. Rando, S. Contino, V. Romano and G.R. Merlo, 2021. Neuronal cytoskeleton in intellectual disability: From systems biology and modeling to therapeutic opportunities. Int. J. Mol. Sci., Vol. 22.
CrossRefDirect Link - Riou, P., P. Villalonga and A.J. Ridley, 2010. Rnd proteins: Multifunctional regulators of the cytoskeleton and cell cycle progression. Bioessays, 32: 986-992.
CrossRefDirect Link - Xu, Y., Q. Sun, F. Yuan, H. Dong and H. Zhang et al., 2020. RND2 attenuates apoptosis and autophagy in glioblastoma cells by targeting the p38 MAPK signalling pathway. J. Exp. Clin. Cancer Res., Vol. 39.
CrossRefDirect Link - Heng, J.I.T., Z. Qu, C. Ohtaka-Maruyama, H. Okado and M. Kasai et al., 2015. The zinc finger transcription factor RP58 negatively regulates Rnd2 for the control of neuronal migration during cerebral cortical development. Cerebral Cortex, 25: 806-816.
CrossRefDirect Link - Alfano, C., L. Viola, J.I.T. Heng, M. Pirozzi and M. Clarkson, 2011. COUP-TFI promotes radial migration and proper morphology of callosal projection neurons by repressing Rnd2 expression. Development, 138: 4685-4697.
CrossRefDirect Link - Liu, Y., J. Han, Z. Zhou and D. Li, 2019. Paeoniflorin protects pancreatic β cells from STZ-induced damage through inhibition of the p38 MAPK and JNK signaling pathways. Eur. J. Pharmacol., 853: 18-24.
CrossRefDirect Link - Takahashi, K., H. Mizukami, S. Osonoi, Y. Takeuchi and K. Kudoh et al., 2021. Islet microangiopathy and augmented β‐cell loss in Japanese non‐obese type 2 diabetes patients who died of acute myocardial infarction. J. Diabetes Invest., 12: 2149-2161.
CrossRefDirect Link - Mezza, T., P.M. Ferraro, G. di Giuseppe, S. Moffa and C.M.A. Cefalo et al., 2021. Pancreaticoduodenectomy model demonstrates a fundamental role of dysfunctional β cells in predicting diabetes. J. Clin. Invest., Vol. 131.
CrossRefDirect Link - Cheng, Y., Q. Hu and J. Zhou, 2021. Silencing of lncRNA PVT1 ameliorates streptozotocin-induced pancreatic β cell injury and enhances insulin secretory capacity by regulating miR-181a-5p. Can. J. Physiol. Pharmacol., 99: 303-312.
CrossRefDirect Link - Ma, J., Y. Wu and Y. He, 2021. Silencing circRNA LRP6 down‐regulates PRMT1 to improve the streptozocin‐induced pancreatic β-cell injury and insulin secretion by sponging miR-9-5p. J. Bioenerg. Biomembr., 53: 333-342.
CrossRefDirect Link - Sun, R., W. Xue and J. Zhao, 2021. Hsa_circ_0054633 mediates apoptosis and insulin secretion in human pancreatic β cells through miR-409-3p/caspase-8 axis. Diabetes Res. Clin. Pract., Vol. 176.
CrossRefDirect Link - Hu, Y., H. Zhao, J. Lu, D. Xie and Q. Wang et al., 2021. High uric acid promotes dysfunction in pancreatic β cells by blocking IRS2/AKT signalling. Mol. Cell. Endocrinol., Vol. 520.
CrossRefDirect Link