
E. Baris
Graduate School of Health Sciences, Dokuz Eylul University, Turkey
O. Simsek
Graduate School of Health Sciences, Dokuz Eylul University, Turkey
H. Efe
Department of Medical Biology, Faculty of Medicine, Dokuz Eylul University, Turkey
S. Oncu
Department of Pharmacology, Faculty of Medicine, Dokuz Eylul University, Turkey
A. Gelal
Department of Pharmacology, Faculty of Medicine, Dokuz Eylul University, Turkey
E. Hamurtekin
Department of Pharmacology, Faculty of Medicine, Eastern Mediterranean University, Turkey
M. Tosun
Department of Pharmacology, Faculty of Medicine, Izmir University of Economics, Turkey
S. Ozbal
Department of Histology and Embryology, Faculty of Medicine, Medical Sciences, Dokuz Eylul University, Turkey
Z. Yuce
Department of Medical Biology, Faculty of Medicine, Dokuz Eylul University, Turkey
M.A. Arici
Department of Pharmacology, Faculty of Medicine, Dokuz Eylul University, Turkey
LiveDNA: 90.28676
International Journal of Pharmacology
Year: 2021 | Volume: 17 | Issue: 2 | Page No.: 84-96







ABSTRACT
Background and Objective: Cytidine-5-diphosphate-choline (CDP-choline) and choline activate the cholinergic anti-inflammatory pathway in case of inflammation. This study investigated the role of CDP-choline and choline along with the contribution of the cyclooxygenase (COX) pathway on the lipopolysaccharide (LPS)-induced endotoxemia model in rats. Materials and Methods: Endotoxemia model was induced by LPS administration. CDP-choline or choline 5 min before and 6 hrs after LPS injection. The sepsis severity, body weight changes, survival rate were evaluated. Serum prostaglandins, Tumour Necrosis Factor (TNF)-α, total choline levels were measured. COX-2 mRNA expression and protein levels were analyzed. Spleen tissues were evaluated histomorphological. One-way analysis of variance analysis (ANOVA) or Kruskal Wallis tests was used for statistical analysis. Results: COX-2 expressions in liver and brain tissues, serum prostaglandin E2, 6-keto prostaglandin F1α, Thromboxane A2 and TNFα levels were increased 24 hrs after LPS administration. Administrations of CDP-choline or choline were decreased COX-2 expression in the liver. Serum prostaglandin levels were decreased in the CDP-choline-treated group, whereas, only prostaglandin E2 level was decreased in the choline-treated group. Total choline levels in serum and brain were increased after CDP-choline or choline administration. Accordingly, serum TNFα levels and TNFα expression in the liver were decreased in CDP-choline and choline-treated groups. TNFα expression in the brain was decreased in the choline-treated group, whereas, increased in the CDP-choline-treated group. Conclusion: CDP-choline and choline decreased LPS-induced COX-2 enzyme expression and prostaglandin levels in the periphery by increasing serum and brain total choline levels in the LPS-induced endotoxemia model in rat.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
E. Baris, O. Simsek, H. Efe, S. Oncu, A. Gelal, E. Hamurtekin, M. Tosun, S. Ozbal, Z. Yuce and M.A. Arici, 2021. Effects of CDP-Choline and Choline on COX Pathway in LPS-Induced Inflammatory Response in Rats. International Journal of Pharmacology, 17: 84-96.
DOI: 10.3923/ijp.2021.84.96
URL: https://scialert.net/abstract/?doi=ijp.2021.84.96
DOI: 10.3923/ijp.2021.84.96
URL: https://scialert.net/abstract/?doi=ijp.2021.84.96
INTRODUCTION
Sepsis, a systemic inflammatory response to infection, triggered by some antigenic components of microorganisms, is characterized by widespread inflammation leading to multiple organ failure1. The rat endotoxemia model is commonly used as it resembles the sepsis-associated acute inflammatory response. Lipopolysaccharide (LPS), an endotoxin of gram-negative bacteria, is a well-known inducer of pathogen-associated molecular pattern (PAMP) recognition receptors (toll-like receptors, TLRs) is used in endotoxemia models2,3. Exposure to LPS leads to endotoxemia which activates mononuclear phagocytic cells (MPCs) through TLRs to initiate systemic inflammation2. Activated MPCs release various pro-inflammatory cytokines such as tumour necrosis factor-α (TNFα), interleukin 1β (IL-1β) and IL-6, causing hemodynamic changes that eventually lead to multiple organ failure4. Furthermore, direct action of endotoxin and stimulation of cytokines simultaneously causes the release of prostaglandins produced via an inducible form of cyclooxygenase (COX-2), an enzyme that contributes to further release of inflammatory mediators involved in cardinal signs of inflammation5-8.
The cholinergic anti-inflammatory pathway (CAP) is a part of the endogenous response to systemic inflammation throughout the body and defined as a neural mechanism that inhibits pro-inflammatory cytokine release resulted from the activation of the vagus nerve via cholinergic receptors9. Alpha7 nicotinic acetylcholine receptors (α7nAChR), expressed in numerous kinds of MPCs are required for the interaction between cholinergic nerves and the immune system. Activation of α7nAChRs on macrophages by acetylcholine (ACh) inhibits pro-inflammatory cytokine release including TNFα and interleukins in different experimental inflammation models10. Therefore, α7nAChR-activating agents appear to be valuable therapeutics for the modulation of systemic inflammatory response.
Cytidine-5-diphosphate-choline (CDP-choline) is rapidly cleaved into choline and cytidine by phosphodiesterases (PDEs) in the cell membrane which in turn increases choline levels in blood circulation and brain11. CDP-choline is an important component of phosphatidylcholine (PC) synthesis, the main phospholipid in the brain, by the kennedy pathway. Additionally, PC is the source of bioactive lipids including arachidonic acids12. PC can be hydrolysed by phospholipase A2 (PLA2) which contributes to the production of arachidonic acid from membrane phospholipids, prostaglandins and leukotrienes13 and it has been demonstrated that PLA2 activity significantly increased in systemic inflammatory conditions14. Moreover, central purinergic signalling has been shown to contribute to the immune response and exacerbation of inflammatory state in sepsis15. Owing to its cytidine moiety, CDP-choline might also exert some anti-inflammatory effectiveness through purinergic receptor activation16.
Choline, a precursor of neurotransmitter ACh, interacts directly with α7nAChRs and induces cholinergic neurotransmission17. Several studies performed on animal models of sepsis/endotoxemia have demonstrated that stimulation of the CAP via CDP-choline and choline enhancing ACh release, moderate pro-inflammatory cytokine levels and improve survival rates by preventing multiple organ failure due to inflammation4,17-21.
Therefore, this study investigated the effects of CDP-choline and choline as well as the role of the COX-2 pathway in the LPS-induced endotoxemia model in rats.
MATERIAL AND METHODS
Study area: This research project was carried out at the Department of Pharmacology, Dokuz Eylul University, Izmir, Turkey from June, 2018-February, 2019.
Materials: Lipopolysaccharide (LPS) from Escherichia coli (O55:B5), Choline chloride, CDP-choline (Cytidine 5-diphosphocholine sodium salt dihydrate) were obtained from Sigma-Aldrich (St. Louis, Mo., USA). All drugs were dissolved in saline (0.9% sodium chloride).
Animals: The experimental protocol of this study was approved by the Dokuz Eylul University, Turkey Multidisciplinary Laboratory Animal Studies Local Ethics Committee (No: 51/2017). Thirty-eight male rats (Wistar albino, 180-200 g) used in the study were individually housed during the experiment under standard controlled conditions (12 hrs dark/light cycle, at 22±2°C). Rats freely accessed food and water during housing. All procedures were performed according to the ‘Principles of Laboratory Animal Care’ by the ‘National Institute of Health’ publication.
Experimental protocol: An experimental endotoxemia model was performed with a sublethal dose of LPS administration (10 mg kg–1)22. Animals with no significant change in their basal body weights were randomly separated into four experimental groups, 1. Control (n = 6), 2. LPS+saline (n = 12), 3. LPS+375 mg kg–1 CDP-choline (n = 10) and 4. LPS+90 mg kg–1 choline (n = 10). Saline or LPS were administered 5 min after the drug injections. Saline, CDP-choline and choline injections were repeated at the 6th hrs of the experimental protocol. All injections (in 1 mL) were performed intraperitoneally (i.p.). The CDP-choline dose was determined as the equimolar dose of choline as reported earlier21.
The severity of endotoxemia was evaluated at the beginning of the experiment and the 6th and 24th hrs during the protocol by using a sepsis scoring system with the highest total score of 2823. Sepsis scoring performed blindly by 3 researchers was averaged and a sum of all individual parameters presented as total sepsis score. Rats were weighed at the beginning and 24th hrs. Survival rates during the experiments were monitored at the 6th and 24th hrs following the LPS exposure. Rats were sacrificed by decapitation at the 24th hrs under mild ether anaesthesia and their trunk blood was collected for prostaglandin and choline measurements. Total RNA and protein samples isolated from liver and brain tissues were used to determine COX-2 mRNA expression and protein levels via real-time reverse transcription-polymerase chain reaction(rtRT-PCR) and Western Blot analyses, respectively. Brain samples were excised with a razor blade by cutting from the edges of hypothalamic tissue22,24. Spleen samples were dissected for histomorphological evaluations. All specimens were transferred into cryotubes and stored at -80°C until the analysis.
Histomorphological analysis: For histomorphological examination, spleen tissue samples were fixed with 10% formalin then embedded in paraffin blocks. Sections of 5 μm thickness were stained with hematoxylin-eosin (H-E) and examined for hemorrhagic and inflammatory changes. A grading system was used to score the tissue injury semi-quantitatively as (0) absent, (1) slight, (2) moderate and (3) severe. Each section was evaluated blindly and the average score determined25.
Image analysis methods: All sections were digitally photographed and analysed by using a computer-assisted image analyser system coupled to a microscope (Olympus BX-51, Japan) with a high-resolution video camera (Olympus DP-1, Japan). For morphometric evaluation, a computerized image analysis system (UTHSC Image Tool software version 3.0, University of Texas Health Science Center, San Antonio, TX) was used.
Reverse Transcription-Polymerase Chain Reaction(RT-PCR) analysis: Liver and brain tissues were rapidly removed and homogenized following the sacrification. Total RNA was isolated by an extraction kit (GeneJET RNA Purification Kit, ThermoScientific) according to the protocol provided by the manufacturer. The quality of RNA was calculated by OD260/OD280 nm absorption ratio. RNA from each sample was converted to complementary DNA using a cDNA synthesis kit (OneScript Plus G236, Applied Biological Materials, Richmond, BC, Canada) according to the manufacturer’s instructions. The cDNA samples were amplified with RT-PCR (Biorad CFXconnect) by using forward and reverse primers of COX-2 and TNFα gene. Cycling conditions were 95°C for 10 min for polymerase activation/denaturation and 40 cycles (95°C for 15 sec and 55°C for 60 sec for COX- 2/56°C for 60 sec for TNFα ) for amplification followed by a dissociation stage (65°C for 5 sec then 5 sec each at 0.5°C for increments between 65-95°C). House-keeping gene β-actin was used as an internal positive control for normalization (Table 1). ΔΔCt method was employed for the relative quantification of mRNA expression22.
Western blot analysis: To evaluate the effects of CDP-choline and choline on the COX-2 pathway, liver and brain tissue homogenates were prepared with a lysis buffer and run on Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE). Proteins separated in the gel were transferred onto polyvinylidene fluoride (PVDF) membranes and immunoblotted with primary antibodies specific to COX-2 and β-actin followed by incubation with goat anti-rabbit IgG antibody-HRP conjugate as a secondary antibody. After blotting, protein bands were visualised by using Enhanced Chemiluminescence (ECL) reagent (ThermoScientific 32132)26.
Measurement of serum prostaglandin, TNFα and choline levels: Serum levels of the inactive non-enzymatic hydrolysis product of PGI2, 6-keto-PGF1α, (Abnova, KA0295), PGE2 (Cusabio, CSB-E07967r), TxA2 (Abnova, KA0295), TNFα (Invitrogen, BMS622) and total choline level in serum and brain (Biovision, K615-100) were measured via ELISA according to the manufacturer’s instructions.
| Table 1: Primer sequences used in real-time quantitative RT-PCR | ||
| Target gene | Forward sequence | Reverse sequence |
| COX-2 | TGTACAAGCAGTGGCAAAGG | TAGCATCTGGACGAGGCTTT |
| TNF α | AAATGGGCTCCCTCTCATCAGTTC | TCTGCTTGGTGGTTTGCTACGAC |
| β-actin | TGTCACCAACTGGGACGATA | GGGGTGTTGAAGGTCTCAAA |
Statistical analysis: One-way analysis of variance analysis (ANOVA) with post hoc Tukey-Kramer multiple comparisons for parametric results, Kruskal Wallis for non-parametric results and Student’s t-test or Mann-Whitney U tests for comparison between two groups were employed where appropriate. The survival rate was analyzed by log-rank (Mantel-Cox) test. Statistical analysis were performed by using GraphPad 5(La Jolla, USA). Data were shown as Mean±Standard Error of the mean (SEM) and p<0.05 was accepted as significant.
RESULTS
Survival percentage: LPS+Saline group (n = 12) has 41.6 and 33.3% survival at the 6th and 24th hrs, respectively. The survival ratio decreased in the LPS+Saline group compared to that of the control group (n = 6) at the 6th and 24th hrs (p<0.05 and p<0.01). The percentage survival in LPS+CDP-choline and LPS+choline groups (n = 10) compared to LPS+Saline group was improved to 80.0% (p = 0.07) at the 6th hrs, whereas that of LPS+CDP-choline group and LPS+choline group at 24th hrs was 50.0 (p = 0.22) and 60.0% (p = 0.44), respectively (Table 2).
Sepsis severity: Total sepsis score, defined as zero in all groups at the beginning of the experiment was increased by LPS+Saline treatment at 6th and 24th hrs (p<0.01), decreased by CDP-choline or choline treatments at 24th hrs (p<0.05) (Table 3). The activity was improved significantly in the LPS+CDP-choline group compared to that of LPS+Saline at the 6th hrs (1.0±0.3 vs. 2.0±0, p<0.05). The levels of consciousness, active respiration rate (0.2±0.2 vs. 2.3±0.7 and 0.5±0.2 vs. 1.5±0.3 respectively, p<0.05) and response to stimulus (0.0±0.0 vs. 1.7±0.5, p<0.01) scores in the 24th hrs were improved significantly by LPS+choline treatment compared to that of LPS+Saline. The appearance, respiration rate and quality were improved by LPS+CDP-choline treatment compared to that of LPS+Saline at the 24th hrs (1.0±0 vs. 2.7±0.7, 0.0±0.0 vs. 1.5±0.3 and 0.0±0.0 vs. 0.7±0.2 respectively, p<0.05).
Body weight loss: Decrease in body weights at the end of the experimental protocol were -27.4±2.1, -21.7±5.2 and -13.6±6.7g in LPS+Saline, LPS+CDP-choline and LPS+choline groups, respectively. In LPS+Saline and LPS+CDP-choline groups, body weight loss was significantly higher than that of the control (p<0.0001 and p<0.005). However, there was no significant difference between the LPS+choline and control groups (p = 0.11) as well as LPS+CDP-choline and LPS+choline groups compared to the LPS+Saline group (p = 0.38 and p = 0.14, respectively) (Fig. 1a).
Histomorphological evaluation in spleen tissue: Histomorphological injury score was significantly increased in spleen tissue in the LPS+Saline group (2.2±0.2) compared to the control group (0.4±0.1) (p<0.001). In the LPS+CDP-choline (1.2±0.0) and LPS+choline (1.5±0.0) groups the histomorphological damage score decreased compared to LPS+Saline group (p<0.05) (Fig. 1b).
| Table 2: Percentage survival in the experimental groups | ||||
| Percentage survival in experimental groups | ||||
| Time (hrs) | Control (n = 6) | LPS+Saline (n = 12) | LPS+CDP-choline (n = 10) | LPS+Choline (n = 10) |
| 6th | 100.0% | 41.6%† | 80.0% | 80.0% |
| 24th | 100.0% | 33.3%†† | 50.0% | 60.0% |
Survival percentage was analyzed by log-rank (Mantel-Cox) test, (n = 6-12 per group), (†), (††); p<0.05, p<0.01 vs. control group | ||||
| Table 3: Evaluation of the sepsis severity in experimental groups | ||||||
| Score (6th hrs) | Score (24th hrs) | |||||
| Variables | LPS+Saline | LPS+Choline | LPS+CDP-choline | LPS+Saline | LPS+Choline | LPS+CDP-choline |
| Appearance | 2.0±0.0** | 2.0±0.0** | 1.6±0.2 | 2.7±0.7** | 1.3±0.3 | 1.0±0.0† |
| Level of consciousness | 2.5±0.3** | 1.2±0.5 | 2.2±0.6 | 2.3±0.7* | 0.2±0.2† | 0.8±0.4 |
| Activity | 2.0±0.0** | 0.8±0.4 | 1.0±0.3† | 1.7±0.2** | 0.3±0.2† | 0.8±0.4 |
| Response to stimulus | 2.3±0.5** | 0.6±0.4 | 1.6±0.2 | 1.7±0.5†† | 0.0±0.0†† | 0.4±0.4 |
| Eyes | 2.0±0.4** | 1.4±0.6 | 1.0±0.6 | 2.0±0.8* | 0.3±0.3 | 0.4±0.2 |
| Respiration rate | 1.5±0.5* | 1.2±0.4 | 1.2±0.2 | 1.5±0.3** | 0.5±0.2† | 0.0±0.0† |
| Respiration quality | 0.7±0.2* | 0.5± 0.2 | 0.8±0.4 | 0.7±0.2* | 0.3±0.2 | 0.0±0.0† |
| Total score | 14.0±1.1** | 8.3±2.5 | 8.6±2.4 | 14.0±2.2** | 3.0±0.8† | 3.0±1.1† |
Total sepsis score was 0 at the 6th and 24th hrs in the control group. Kruskal-Wallis and Mann-Whitney U tests were used for statistical analysis. Data were shown as Mean±SEM (n = 4-6 per group). (*), (**); p<0.05, p<0.01, p<0.001 vs. control group, (†), (††); p<0.05, p<0.01 vs. LPS+Saline group | ||||||
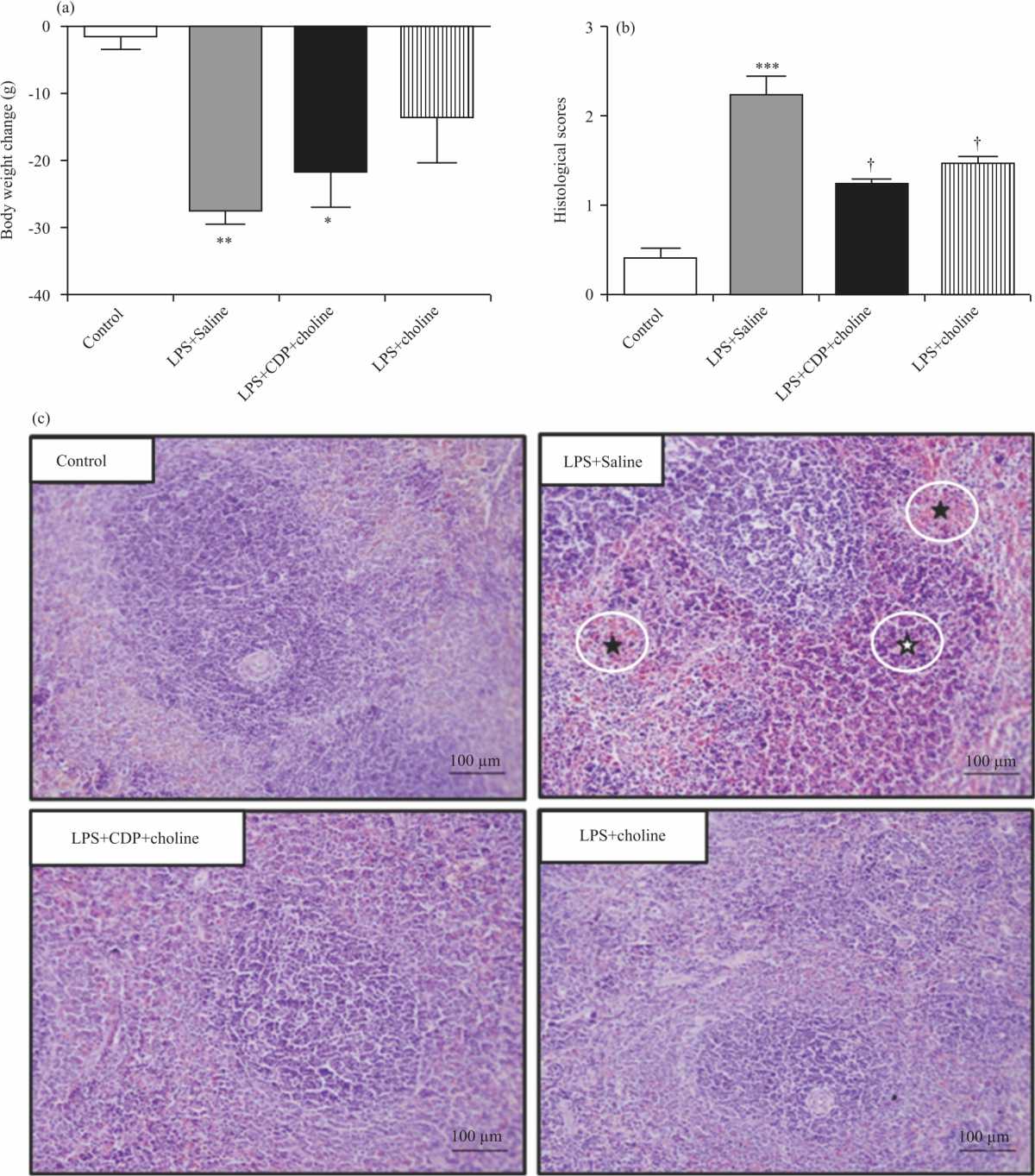 |
| Fig. 1(a-c): | Effects of CDP-choline (375 mg kg–1) and choline (90 mg kg–1) on sepsis severity Shown are body weight loss (A), histomorphological scores (B) and Representative light-microscopic images of H-E staining in spleen tissue (C) of experimental groups; ( ✫) indicates mononuclear cell infiltration and (★) indicates haemorrhage in spleen tissue. One-way analysis of variance analysis (ANOVA) with post hoc Tukey-Kramer multiple comparisons, Kruskal Wallis with Mann Whitney U and Student’s t-tests were used for statistical analysis. Data were shown as Mean±S.E.M. (n = 4-6 per group). (*), (**), (***); p<0.05, <0.01, p<0.001 vs. control group. (†), p<0.05 vs LPS+Saline group |
H-E staining revealed a normal histological structure of spleen tissue in the control group. Spleen tissues showed diffuse and severe haemorrhage and mild inflammation with Mononuclear Phagocytic Cell (MPC) infiltration in LPS+Saline group, reduced spleen injury with little haemorrhage and inflammation in LPS+CDP-choline and LPS+choline groups (Fig. 1c).
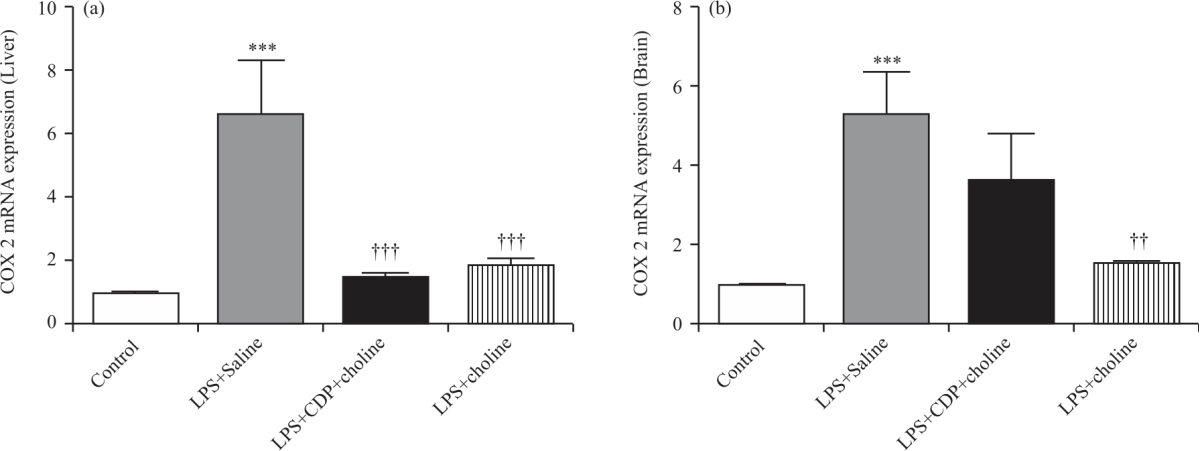 |
| Fig. 2(a-b): | COX-2 enzyme mRNA expressions in the liver (A) and brain (B) tissues in experimental groups ΔΔCt method was employed for the relative quantification of mRNA expression. Kruskal Wallis and Mann Whitney U tests were used for statistical analysis. Data were normalized to beta-actin and shown as Mean±S.E.M. (n = 4-6 per group). (**), (***); p<0.01, p<0.001 vs. control group. (††),(†††); p<0.01,p<0.001 vs. LPS+Saline group |
COX-2 mRNA and protein expression: COX-2 mRNA expression increased significantly in liver and brain tissues by LPS+Saline treatment compared to that of control (p<0.001, Fig. 2a, b). COX-2 mRNA expressions in liver samples from CDP-choline- and choline-treated groups decreased significantly compared to the LPS+Saline group (p<0.001, Fig. 2a). Although not statistically significant, COX-2 mRNA expression in brain samples increased in CDP-choline- treated group compared to that of the control. However, COX-2 mRNA expression in the brain significantly decreased in a choline-treated group compared to the LPS+Saline group (p<0.01). In the LPS+saline group, COX-2 protein expressions in liver and brain samples increased compared to the control group (p<0.001 and p<0.05, respectively). COX-2 protein expressions in liver samples from CDP-choline- and choline-treated groups decreased significantly compared to the LPS+Saline group (p<0.01 and p<0.05 respectively, Fig. 3a) without any significant change in the brain (Fig. 3b).
Prostaglandin levels: Serum levels of PGE2, 6-keto-PGF1α and TXA2 increased (in pg mL–1: 13.3±3.1-39.0±6.0, 1387.0±296.3-2975.0±869.9 and 539.1±74.4-1227.0±121.2, p<0.05, p = 0.06 and p<0.05, respectively) in the LPS+Saline group compared to that of control, whereas decreased significantly in LPS+CDP-choline group compared to that of LPS+Saline (in pg mL–1: 13.7±4.1, 779.7±143.5, 520.0±105.6, p<0.05). Only serum PGE2 levels were decreased significantly (15.6±5.4 pg mL–1, p<0.05) by LPS+choline comparable to that of LPS+Saline (Fig. 4a). Although the serum 6-keto-PGF1α and TXA2 levels decreased in the LPS+choline group, the data did not reach any statistical significance (Fig. 4b-c).
Serum and brain total choline levels: Total choline levels in serum and brain were not altered significantly in the LPS+Saline group compared to that of the control group (in μM: 301.7±15.3 vs. 298.8±6.0 p = 0.84 and -181.4±30.9 vs. 124.7±3.8 p = 0.053, respectively). Serum and brain total choline levels increased significantly in LPS+CDP-choline (in μM: 396.0±19.44 and 155.5±14.19, p<0.05, p<0.05) and LPS+choline (in μM: 387.0±31.6 and 185.8±7. 6, p<0.05, p<0.05) groups comparable to control (Fig. 5).
TNFα mRNA expression and protein level: TNFα mRNA expression was increased significantly in liver and brain tissues by LPS+Saline treatment compared to that of control (p<0.001, Fig. 6a-b). TNFα mRNA expressions in liver samples from CDP-choline- and choline-treated groups decreased significantly compared to the LPS+Saline group (p<0.001). TNFα mRNA expressions in brain samples significantly increased in CDP-choline- treated group (p<0.001) while significantly decreased in the choline-treated group (p<0.01) compared to the LPS+Saline group (Fig. 6a-b).
Serum TNFα levels increased (in pg mL–1: 48.5±3.9, p<0.01) in the LPS+Saline group compared to that of control (in pg mL–1: 18.4±3.4), whereas decreased significantly in LPS+CDP-choline group (in pg mL–1: 16.1±5.2, p<0.01) and LPS+Choline group (in pg mL–1: 15.1±4.9, p<0.01) compared to that of LPS+Saline (Fig. 6c).
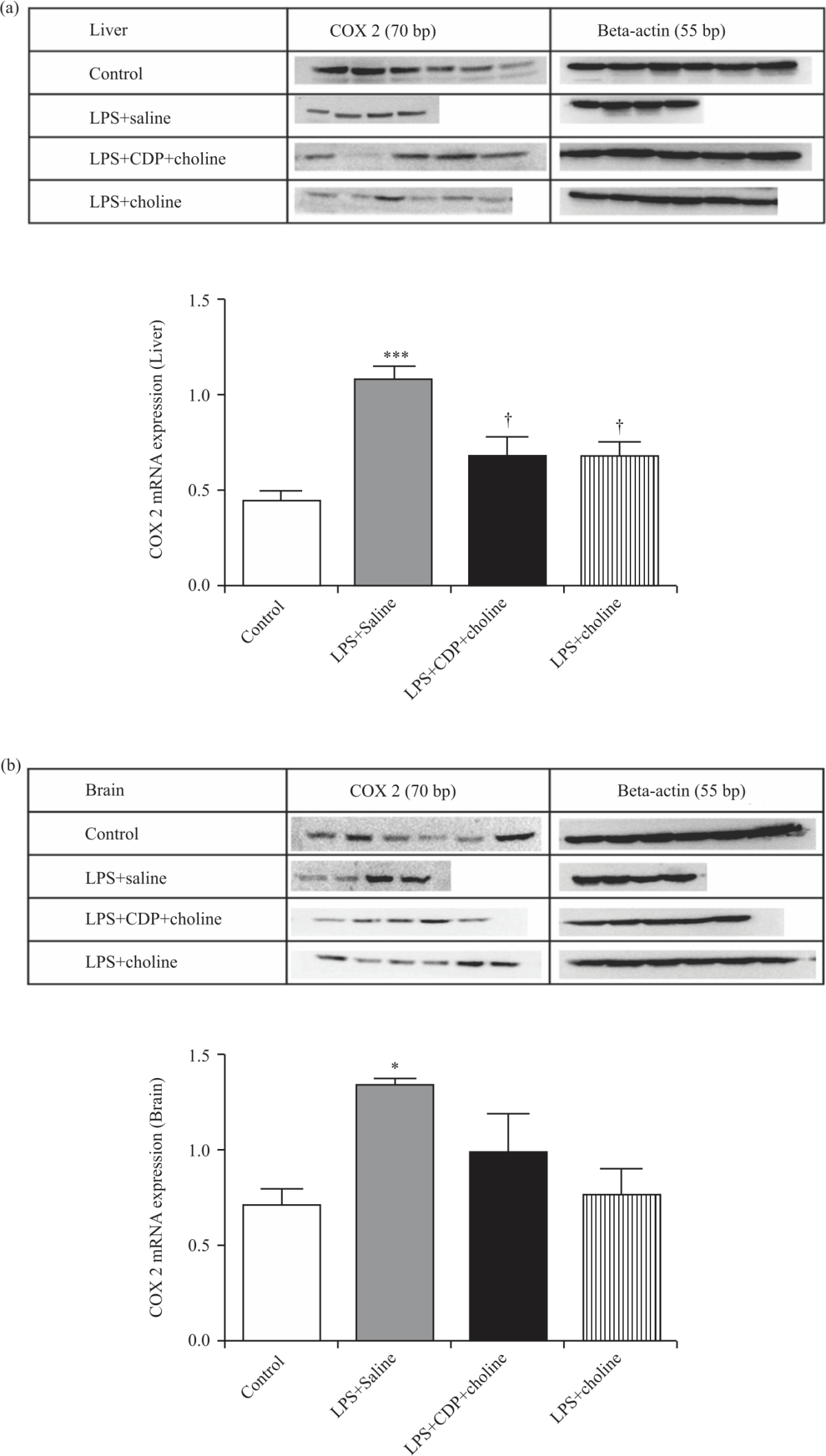 |
| Fig. 3(a-b): | COX-2 protein expressions in the liver (A) and brain (B) tissues from experimental groups One-way analysis of variance analysis (ANOVA) with post hoc Tukey-Kramer multiple comparison tests were used for statistical analyses. Data were normalized to beta-actin and shown as mean±S.E.M. (n = 4-6 per group). (*), (***); p<0.05, p<0.001 vs. control group. (†); p<0.05 vs. LPS+Saline group |
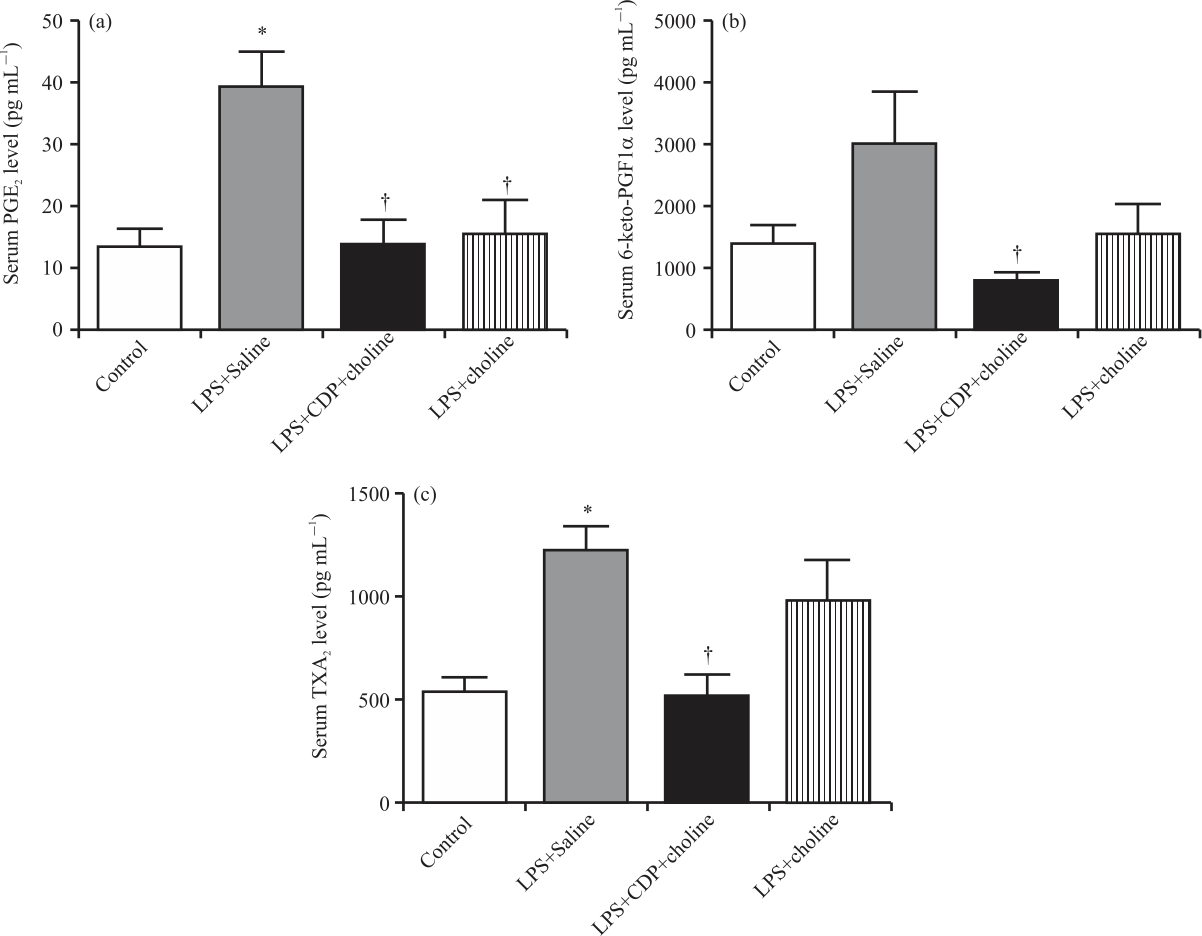 |
| Fig. 4(a-c): | Serum prostaglandin levels in experimental groups Shown are PGE2 (A); 6-keto-PGF1a (B), TxA2 (C) (PGE2: Prostaglandin E2, 6-keto-PGF1a: 6-keto Prostaglandin F1a, TxA2: Thromboxane A2. One-way analysis of variance analysis (ANOVA) with post hoc Tukey-Kramer multiple comparison tests were used for statistical analyses. Data were shown as mean±S.E.M. (n = 4-6 per group). (*); p<0.05 vs. control group. (†); p<0.05 vs. LPS+Saline group |
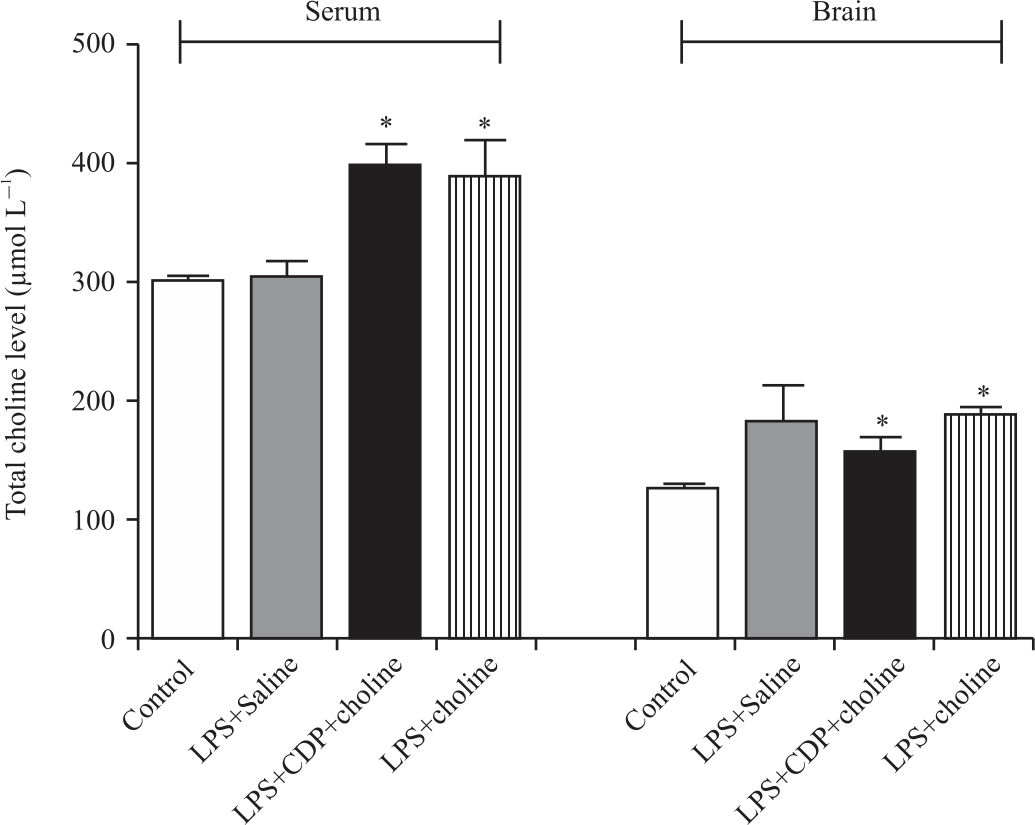 |
| Fig. 5: | Serum and brain total choline levels in experimental groups One-way analysis of variance analysis (ANOVA) with post hoc Tukey-Kramer multiple comparison tests and Student’s t-tests were used for statistical analyses. Data were shown as mean±S.E.M. (n = 4-6 per group). (*), p<0.05 vs control group |
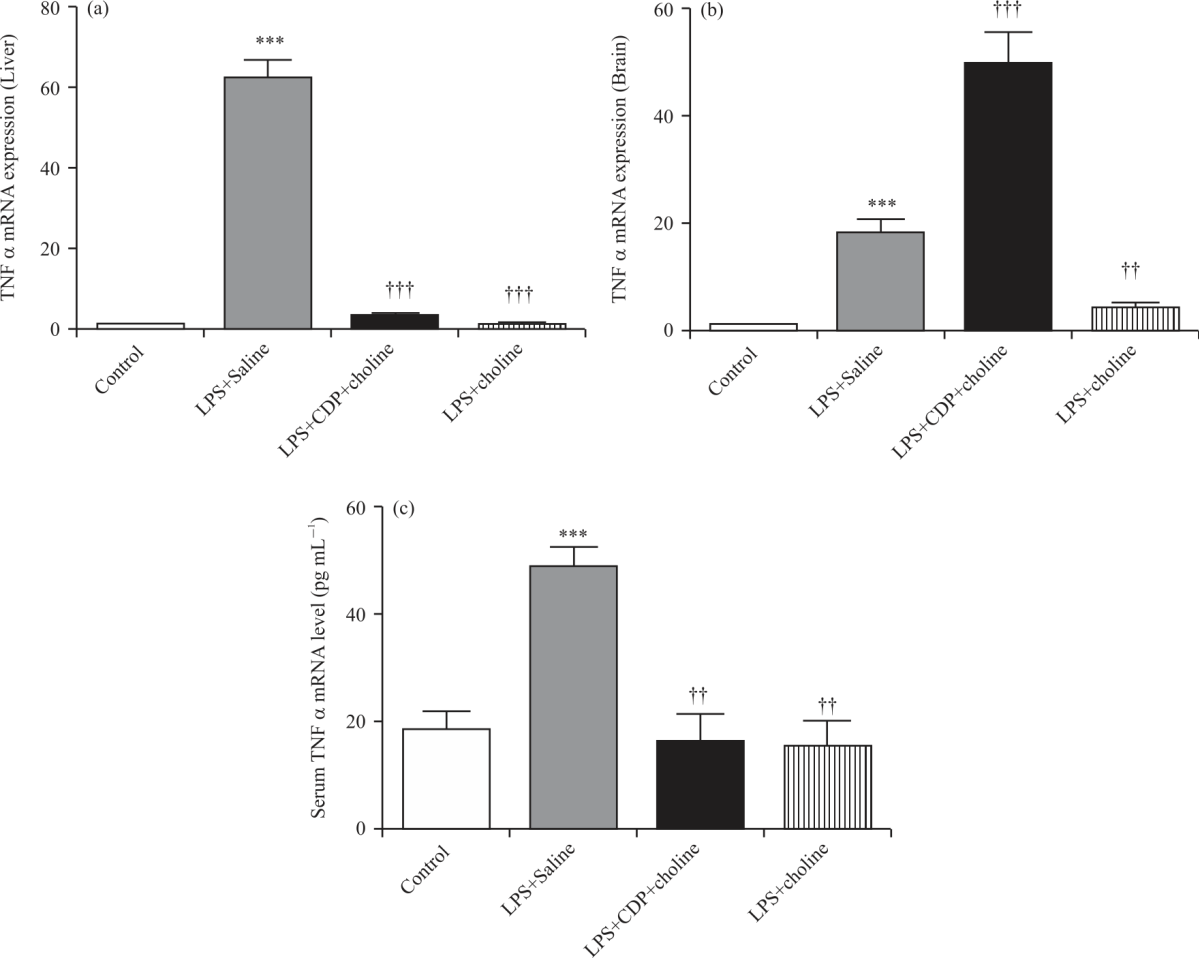 |
| Fig. 6(a-c): | TNF a mRNA expressions in the liver (A) and brain (B) tissues and serum TNF a levels (C) in experimental groups ΔΔCt method was employed for the relative quantification of mRNA expression. One-way analysis of variance analysis (ANOVA) with post hoc Tukey-Kramer multiple comparison tests and Student’s t-tests were used for statistical analyses. Data were normalized to beta-actin and shown as Mean±SEM, (n = 4-6 per group). (**), (***); p<0.01, p<0.001 vs. control group. (††),(†††); p<0.01, p<0.001 vs. LPS+Saline group |
DISCUSSION
This study investigated the role of cytidine-5-diphosphate-choline (CDP-choline) and choline as well as the contribution of the COX-2 pathway on the inflammatory response induced by lipopolysaccharide (LPS) in endotoxemic rats.
In this study, endotoxemia was induced by 10 mg kg–1 LPS22 and the success of the model was determined by evaluating the rats' survival rates, sepsis scoring, body weight changes and histomorphological changes in the spleen. The dose was in the upper limit of the related reported studies and the rate of survival, increase in sepsis scoring and body weight decrease were compatible with the previous studies in LPS induced endotoxemia. Although not statistically significant, CDP-choline and choline have been detected beneficial effects on improving survival rates in early phases of endotoxin challenge. The survival rate (20% in 24 hrs) in the present study was similar to that published earlier. Shrum et al.23 demonstrated that survival rates diminish to 20.0% in a murine animal model with total sepsis scores over 15. Previous observations reported that CDP-choline and choline increased survival rates by diminishing LPS induced organ injury, preventing disseminated intravascular coagulation, changing serum lipid responses and decreasing systemic inflammatory parameters in endotoxemic animals21,27. Along with previous reports, CDP-choline and choline treatments appear to improve the survival rate via exerting protective effects in multiple organ systems in endotoxemic animals.
Sepsis is an acute inflammatory response and endotoxemia is a model of acute inflammation, therefore we used a novel scoring system composed of different variables demonstrated to evaluate sepsis severity in animal models which provides reliable and consistent results23,28. The evaluated parameters in sepsis scoring in rats were appearance, level of consciousness, activity, response to stimulus, eyes, respiratory rate and quality. The data showed that total sepsis scores as well as individual parameters, were significantly poor in the LPS+Saline group compared to the control group. Whereas CDP-choline and choline treatments significantly decreased total sepsis scores and individual parameters at late stages of sepsis challenge.
Response to stimulus in sepsis scoring was improved in the LPS+choline group compared to the LPS+Saline group. These novel findings might be related to the beneficial effects of choline on motor coordination. It was reported that the impaired motor coordination in endotoxemic mice29. Choline deficient diet is correlated with increased motor dysfunction in rats30,31. Respiratory parameters were improved in both treatment groups compared to the LPS+Saline group. CDP-choline and choline decreased the respiratory inflammation in the lung by decreasing inflammatory mediators released from mononuclear phagocytic cells (MPCs) were reported earlier32,33. Consistent with our findings, improvement in respiratory parameters might be related to the decreased pulmonary inflammation which should be confirmed by further studies.
LPS-induced endotoxemia/sepsis is known to be associated with sickness behaviour including appetite loss and body weight decreases caused by the central effect of cytokines34. LPS-induced body weight loss was partially recovered in treatment groups compared to the LPS+Saline group in this study. Although not statistically significant, CDP-choline and choline treatment decreased body weight loss. Accordingly, previously demonstrated that choline-supplemented diet recovered maternal, fetal and placental weight loss in LPS induced endotoxemia model of rats through the action on α7nAChRs and cholinergic anti-inflammatory pathway35. We consider that CAP activation by CDP-choline and choline might account for its protective effects on LPS induced body weight loss in endotoxemic rats. Taken together, the protective effects of CDP-choline and choline on LPS-induced body weight loss via α7nAChRs needs to be studied further.
The spleen is a major organ that contributes to cholinergic control of systemic inflammatory response36. Histomorphological examination of lung, liver and kidney tissues from rats with CLP-induced sepsis showed that inflammation, vascular degeneration, haemorrhage and necrosis parameters were improved by CDP-choline treatment without affecting the spleen37. Recent studies have been implicated that CDP-choline exerts tissue-protective effects by reducing oxidative stress in ischemic and inflammatory conditions37,38. COX-2 inhibitor celecoxib ameliorated tissue damage in spleen, liver and lungs in rats with CLP model of sepsis via decreasing COX-2 mediated oxidative damage39. We observed that severe haemorrhage and mild inflammation with MPC infiltration decreased in the spleen tissues from LPS-induced endotoxemic rats. Based on our observation, decreased circulating prostaglandin levels and improved sepsis severity suggest that inflammatory response might be suppressed with CDP-choline and choline treatments in the spleen. However, the exact role of CDP-choline or choline on LPS-induced tissue damage together with the involvement of COX-2 downregulation and antioxidant effect of the compounds needs to be clarified.
COX-2 expression and its products increased in the LPS+Saline group compared to that of control confirming the impact of COX-2 upregulation in LPS-induced endotoxemia40,41. COX-2 upregulation is also known to contribute to inflammation by releasing prostaglandins42. Furthermore, COX-2 upregulation together with increased prostaglandin levels under the inflammatory conditions is associated with increased vascular permeability, the unrestrained release of inflammatory mediators and reactive radicals, hemodynamic changes eventually yielding multiple organ failure and death5,6,7. Co-administration of choline and acetylsalicylic acid in LPS-induced endotoxemia in mice showed a synergistic anti-inflammatory interaction between the compounds via affecting the arachidonic acid pathway41. Accordingly, our data showed that COX-2 expressions in the liver decreased in LPS+CDP-choline and LPS+choline groups. Accordingly, PGE2, a stable PGI2 metabolite 6-keto-PGF1α and TXA2 levels were decreased in LPS+CDP-choline group compared to that of LPS+Saline. Consistent with our observations, several studies showed activation of the cholinergic anti-inflammatory pathway by CDP-choline and choline attenuates pro-inflammatory cytokine levels in animal sepsis models4,11,20.
There are some conflicting results about the action of CDP-choline and choline in brain tissues. In study investigating blood pressure and heart rate in healthy rats revealed that intracerebroventricular CDP-choline administration increased COX-2 reactivity in the hypothalamus43. Nicotine administration causes an increase in COX-2 expression and PGE2 level in microglial cells by activating the cholinergic anti-inflammatory pathway44. In this study, CDP-choline decreased the LPS induced COX-2 expression in the liver without affecting brain levels compared to the LPS+Saline group.
It is well known that endotoxemia leads to inflammation and an increase in TNFα levels in the brain45. Elevation in IL1β and TNFα levels increases PC hydrolysis and PLA2 activity. Evidence suggest that CDP-choline attenuated the loss of phospholipids and increase arachidonic acid in case of cerebral ischemia46. The study showed that i.p. CDP-choline significantly increased LPS induced TNFα expression without affecting COX-2 expression under the hypothesis suggest that beneficial effects of CDP-choline in the brain are opposed to TNFα via counteracting TNFα mediated events47. In our study downregulated COX-2 activity was also confirmed by decreased serum prostaglandin and TNFα levels.
Several studies demonstrated that exogenously administered CDP-choline and choline elevates total choline levels in the brain and blood circulation11,20,48. As shown earlier, the LPS challenge increases serum total choline concentrations by activating endogenous cholinergic anti-inflammatory pathway which is further enhanced by administration of choline and CDP-choline activating cholinergic receptors expressed on MPCs9,18. According to our findings, total choline levels did not increase in serum in response to LPS administration whereas it slightly increased in brain tissues although statistically not significant. However, i.p. administration of CDP-choline or choline elevated total choline concentrations in serum and brain tissues of endotoxemic animals. Briefly, besides peripheral anti-inflammatory effects, increasing total choline levels in the brain and central activation of the cholinergic anti-inflammatory pathway may account for protective effects of CDP-choline or choline in endotoxemic rats.
The underlying mechanism of CDP-choline and choline on the COX pathway in LPS-induced inflammatory response and its severity were not examined in the scope of the present experimental protocol. Moreover, it was not possible to monitor changes in COX-2 expression levels for a longer period due to poor survival rates in the LPS+Saline group. The sensitivity of the ELISA kit was not high enough to determine tissue prostaglandin levels. Although appears to be relevant at the pre-clinical level, the protective effects of CDP-choline and choline through the COX pathway and cholinergic receptor involvement in LPS-induced endotoxemia/sepsis needs to be confirmed unequivocally by further studies.
CONCLUSION
This study demonstrated that the anti-inflammatory effects of CDP-choline or choline appear to be associated with downregulation of COX-2 expression and proportional decreases in prostaglandin and TNFα levels in the periphery, however, the central action of CDP-choline counteracted this effect possibly via TNFα mediated mechanisms. CDP-choline or choline improved survival rate and decreased sepsis severity in endotoxemic rats.
SIGNIFICANCE STATEMENT
This study discovers the effects of CDP-choline and choline on cyclooxygenase (COX)-2 activity and prostaglandins in the central and periphery, which are known to contribute cardinal signs of inflammation. The results are providing supportive evidence on the anti-inflammatory role of CDP-choline and choline along with other pre-clinical studies allowing the development of candidate cholinomimetics in the treatment of sepsis and other inflammatory diseases.
ACKNOWLEDGMENTS
The authors acknowledge Prof. Dr. İsmail Hakkı Ulus (Acıbadem Univ., Istanbul) for his initial contributions of the study and Prof. Dr. Belgin Ünal (Dokuz Eylul Univ., Izmir) for statistical consulting. This research is fully sponsored by Dokuz Eylul University, Turkey with grant number {2018.KB.SAG.055}.
REFERENCES
- Stearns-Kurosawa, D.J., M.F. Osuchowski, C. Valentine, S. Kurosawa and D.G. Remick, 2011. The pathogenesis of sepsis. Annu. Rev. Pathol. Mech. Dis., 6: 19-48.
CrossRefDirect Link - Murdock, J.L. and G. Núñez, 2016. TLR4: The winding road to the discovery of the LPS receptor. J. Immunol., 197: 2561-2562.
CrossRefDirect Link - Dickson, K. and C. Lehmann, 2019. Inflammatory response to different toxins in experimental sepsis models. Int. J.Mol. Sci., Vol. 2019.
CrossRefDirect Link - Ilcol, Y.O., Z. Yilmaz, I.H. Ulus, 2005. Endotoxin alters serum-free choline and phospholipid-bound choline concentrations, and choline administration attenuates endotoxin-induced organ injury in dogs. Shock, 24: 288-293.
CrossRefDirect Link - Hata, A.N. and R.M. Breyer, 2004. Pharmacology and signaling of prostaglandin receptors: Multiple roles in inflammation and immune modulation. Pharmacol. Ther., 103: 147-166.
CrossRefPubMedDirect Link - Ricciotti, E. and G.A. FitzGerald, 2011. Prostaglandins and inflammation. Arteriosclerosis Thrombosis Vasc. Biol., 31: 986-1000.
CrossRefPubMedDirect Link - Hirata, T. and S. Narumiya, 2012. Prostanoids as regulators of innate and adaptive immunity. Adv. Immunol., 116: 143-174.
CrossRefDirect Link - Helliwell, R.J.A, L.F. Adams and M.D. Mitchell, 2004. Prostaglandin synthases: Recent developments and a novel hypothesis. Prostaglandins, Leukotrienes Essent. Fatty Acids, 70: 101-113.
CrossRefDirect Link - Pavlov, V.A., H. Wang, C.J. Czura, S.G. Friedman and K.J. Tracey, 2003. The cholinergic anti-inflammatory pathway: A missing link in neuroimmunomodulation. Mol. Med., 9: 125-134.
Direct Link - Wang, H., M. Yu, C.A. Amella, M. Ochani and M. Tanovic et al., 2003. Nicotinic acetylcholine receptor α7 subunit is an essential regulator of inflammation. Nature, 421: 384-388.
CrossRefPubMedDirect Link - Savci, V., G. Goktalay, M. Cansev, S. Cavun, M.S. Yilmaz and I.H. Ulus, 2003. Intravenously injected CDP-choline increases blood pressure and reverses hypotension in haemorrhagic shock: Effect is mediated by central cholinergic activation. Eur. J. Pharmacol., 468: 129-139.
CrossRefDirect Link - Cui, Z. and M. Houweling, 2002. Phosphatidylcholine and cell death. Biochim. Biophys. Acta. Mol. Cell Biol. Lip., 1585: 87-96.
CrossRefDirect Link - Six, D.A. and E.A. Dennis, 2000. The expanding superfamily of phospholipase A2 enzymes: Classification and characterization. Biochimica Biophysica Acta (BBA)-Mol. Cell Biol. Lipids, 1488: 1-19.
CrossRefDirect Link - Tan, T.L. and Y.Y. Goh, 2017. The role of group IIA secretory phospholipase A2 (sPLA2-IIA) as a biomarker for the diagnosis of sepsis and bacterial infection in adults-A systematic review. PLoS ONE, Vol. 12.
CrossRefDirect Link - Ledderose, C., Y. Bao, Y. Kondo, M. Fakhari, C. Slubowski, J. Zhang and W.G. Junger, 2016. Purinergic signaling and the immune response in sepsis: A review. Clin. Ther., 38: 1054-1065.
CrossRefDirect Link - Bagdas, D., F.A. Sonat, E. Hamurtekin, S. Sonal and M.S. Gurun, 2011. The antihyperalgesic effect of cytidine-5′-diphosphate-choline in neuropathic and inflammatory pain models. Behav. Pharmacol., 22: 589-598.
CrossRefDirect Link - Parrish, W.R., M. Rosas-Ballina, M. Gallowitsch-Puerta, M. Ochani and K. Ochani et al., 2008. Modulation of TNF release by choline requires alpha7 subunit nicotinic acetylcholine receptor-mediated signaling. Mol. Med., 14: 567-574.
CrossRefDirect Link - Pavlov, V.A., M. Ochani, L.H. Yang, M. Gallowitsch-Puerta and K. Ochani et al., 2007. Selective α7-nicotinic acetylcholine receptor agonist GTS-21 improves survival in murine endotoxemia and severe sepsis. Crit. Care Med., 35: 1139-1144.
CrossRefDirect Link - Yilmaz, Z., O.E. Inan, M. Kocaturk, A.T. Baykal and O. Hacariz et al., 2016. Changes in serum proteins after endotoxin administration in healthy and choline-treated calves. BMC Vet. Res., Vol. 12.
CrossRefDirect Link - Ilcol, Y.O., M. Cansev, M.S. Yilmaz, E. Hamurtekin and I.H. Ulus, 2007. Intraperitoneal administration of CDP-choline and its cholinergic and pyrimidinergic metabolites induce hyperglycemia in rats: Involvement of the sympathoadrenal system. Arch. Physiol. Biochem., 113: 186-201.
CrossRefDirect Link - Ilcol, Y.O., Z. Yilmaz, M. Cansev and I.H. Ulus, 2009. Choline or CDP-choline alters serum lipid responses to endotoxin in dogs and rats: Involvement of the peripheral nicotinic acetylcholine receptors. Shock, 32: 286-294.
CrossRefDirect Link - Tunctan, B., B. Korkmaz, A.N. Sari, M. Kacan and D. Unsal et al., 2013. Contribution of iNOS/sGC/PKG pathway, COX-2, CYP4A1 and gp91 phox to the protective effect of 5, 14-HEDGE, a 20-HETE mimetic, against vasodilation, hypotension, tachycardia and inflammation in a rat model of septic shock. Nitric Oxide, 33: 18-41.
CrossRefDirect Link - Shrum, B., R.V. Anantha, S.X. Xu, M. Donnelly, S.M. Haeryfar, J.K. McCormick and T. Mele, 2014. A robust scoring system to evaluate sepsis severity in an animal model. BMC Res. Notes, Vol. 7.
CrossRefDirect Link - Udvari, E.B., K. Völgyi, P. Gulyássy, D. Dimén and V. Kis et al., 2017. Synaptic proteome changes in the hypothalamus of mother rats. J. Proteomics, 159: 54-66.
CrossRefDirect Link - Ozer, E.K., M.T. Goktas, I. Kilinc, S. Pehlivan, H. Bariskaner, C. Ugurluoglu and A.B. Iskit, 2017. Coenzyme Q10 improves the survival, mesenteric perfusion, organs and vessel functions in septic rats. Biomed. Pharmacother., 91: 912-919.
CrossRefDirect Link - Erac, Y., C. Selli, B. Kosova, K.C. Akcali and M. Tosun, 2010. Expression levels of TRPC1 and TRPC6 ion channels are reciprocally altered in aging rat aorta: Implications for age-related vasospastic disorders. AGE, 32: 223-230.
CrossRefDirect Link - Yilmaz, Z., Y. Ozarda, M. Cansev, O. Eralp, M. Kocaturk and I.H. Ulus, 2010. Choline or CDP-choline attenuates coagulation abnormalities and prevents the development of acute disseminated intravascular coagulation in dogs during endotoxemia. Blood Coagul. Fibrinolysis. 21: 339-348.
CrossRefDirect Link - Huet, O., D. Ramsey, S. Miljavec, A. Jenney and C. Aubron et al., 2013. Ensuring animal welfare while meeting scientific aims using a murine pneumonia model of septic shock. Shock, 39: 488-494.
CrossRefDirect Link - Zhao, J., W. Bi, S. Xiao, X. Lan and X. Cheng et al., 2019. Neuroinflammation induced by lipopolysaccharide causes cognitive impairment in mice. Sci. Rep., Vol. 9.
CrossRefDirect Link - Idrus, N.M., K.R. Breit and J.D. Thomas, 2017. Dietary choline levels modify the effects of prenatal alcohol exposure in rats. Neurotoxicol. Teratol., 59: 43-52.
CrossRefDirect Link - Pacelli, C., A. Coluccia, I. Grattagliano, T. Cocco and G. Petrosillo et al., 2010. Dietary choline deprivation impairs rat brain mitochondrial function and behavioral phenotype. J. Nutr., 140: 1072-1079.
CrossRefDirect Link - Kistemaker, L.E.M., D.J. Slebos, H. Meurs, H.A.M. Kerstjens and R. Gosens, 2015. Anti-inflammatory effects of targeted lung denervation in patients with COPD. Eur. Respir. J., 46: 1489-1492.
CrossRefDirect Link - Yamada, M. and M. Ichinose, 2018. The cholinergic pathways in inflammation: A potential pharmacotherapeutic target for COPD. Front. Pharmacol., Vol. 9.
CrossRefDirect Link - Lawrence, C.B., D. Brough and E.M. Knight, 2012. Obese mice exhibit an altered behavioural and inflammatory response to lipopolysaccharide. DMM Dis. Model Mech., 5: 649-659.
CrossRefDirect Link - Zhang, M., X. Han, J. Bao, J. Yang, S.Q. Shi, R.E. Garfield and H. Liu, 2018. Choline supplementation during pregnancy protects against gestational lipopolysaccharide-induced inflammatory responses. Reprod. Sci., 25: 74-85.
CrossRefDirect Link - Martelli, D., M.J. McKinley and R.M. McAllen, 2014. The cholinergic anti-inflammatory pathway: A critical review. Auton. Neurosci., 182: 65-69.
CrossRefDirect Link - Sevim, Ç., B. Altinbas, M. Yalçin, S. Inan, M.Ö. Özyigit, I. Arican and D. Fazla , 2017. Protective effect of CDP-choline on hypotension and tissue injury in septic shock model. Ankara Üniv. Vet. Fak. Derg., 64: 103-110.
CrossRefDirect Link - Zazueta, C., M. Buelna-Chontal, A. Macías-López, N.G. Román-Anguiano and H. González-Pacheco et al., 2018. Cytidine-5'-diphosphocholine protects the liver from ischemia/reperfusion injury preserving mitochondrial function and reducing oxidative stress. Liver Transpl., 24: 1070-1083.
CrossRefDirect Link - Ozer, E.K., M.T. Goktas, I. Kilinc, H. Bariskaner, C. Ugurluoglu and A.B. Iskit, 2017. Celecoxib administration reduced mortality, mesenteric hypoperfusion, aortic dysfunction and multiple organ injury in septic rats. Biomed. Pharmacother., 86: 583-589.
CrossRefDirect Link - Smyth, E.M., T. Grosser, M. Wang, Y. Yu, G.A.F. Gerald, 2009. Prostanoids in health and disease. J. Lipid Res., 50: 423-428.
CrossRefDirect Link - Pan, Z.Y. and H. Wang, 2014. Synergistic interaction between choline and aspirin against acute inflammation induced by carrageenan and lipopolysaccharide. Int. Immunopharmacol., 20: 229-237.
CrossRefDirect Link - Adib-Conquy, M. and J.M. Cavaillon, 2007. Stress molecules in sepsis and systemic inflammatory response syndrome. FEBS Lett., 581: 3723-3733.
CrossRefDirect Link - Topuz, B.B., B. Altinbas, T. Ilhan, M.S. Yilmaz and H. Erdost et al., 2014. Centrally administered CDP-choline induced cardiovascular responses are mediated by activation of the central phospholipase-prostaglandin signaling cascade. Brain Res., 1563: 61-71.
CrossRefDirect Link - De Simone, R., M.A. Ajmone-Cat, D. Carnevale and L. Minghetti, 2005. Activation of α7 nicotinic acetylcholine receptor by nicotine selectively up-regulates cyclooxygenase-2 and prostaglandin E2 in rat microglial cultures. J. Neuroinflamm., Vol. 2.
CrossRefDirect Link - Alexander, J.J., A. Jacob, P. Cunningham, L. Hensley and R.J. Quigg, 2008. TNF is a key mediator of septic encephalopathy acting through its receptor, TNF receptor-1. Neurochem. Int., 52: 447-456.
CrossRefDirect Link - Rao, A.M., J.F. Hatcher and R.J. Dempsey, 2001. Does CDP-choline modulate phospholipase activities after transient forebrain ischemia? Brain Res., 893: 268-272.
CrossRefDirect Link - Adibhatla, R.M. and J.F. Hatcher, 2005. Cytidine 5'-diphosphocholine (CDP-Choline) in stroke and other CNS disorders. Neurochem. Res., 30: 15-23.
CrossRefDirect Link - Köppen, A., J. Klein, T. Holler and K. Löffelholz, 1993. Synergistic effect of nicotinamide and choline administration on extracellular choline levels in the brain. J. Pharmacol. Exp. Ther., 266: 720-725.
Direct Link