
Ali Tureyen
Department of Gastroenterology, Ministry of Health Eskisehir City Hospital, Eskisehir, Turkey
Sevilay Gunay
Department of Molecular Biology and Genetics, Faculty of Science, Bartin University, Bartin, Turkey
Yavuz Erden
Department of Molecular Biology and Genetics, Faculty of Science, Bartin University, Bartin, Turkey
Sinan Ince
Department of Pharmacology and Toxicology, Faculty of Veterinary Medicine, Afyon Kocatepe University, Afyonkarahisar, Turkey
LiveDNA: 90.3724
International Journal of Pharmacology
Year: 2021 | Volume: 17 | Issue: 1 | Page No.: 57-64







ABSTRACT
Background and Objective: Colorectal cancer is the second most common type of cancer in the world. Boric acid has been reported for its efficacy to inhibit various cancer cells. The study aimed to investigate the antiproliferative and apoptotic effects of boric acid in a colorectal adenocarcinoma cell line (HT-29) and its role on chemotherapeutic effects of 5-fluorouracil (5-FU). Materials and Methods: Boric acid and 5-FU were treated to HT-29 cell line for 24 h and effects of boric acid and 5-FU on cell viability, DNA damage and apoptosis were investigated in HT-29 cells. Results: The results of the study revealed that boric acid and 5-FU significantly reduced cell viability at high doses. Low dose administration, where boric acid did not affect cell viability, did not cause genotoxicity and did not significantly change the expression of proteins involved in the apoptotic process. Moreover, boric acid administered with 5-FU partially prevented the chemotherapeutic efficacy of 5-FU by reducing its genotoxic effect. Conclusion: Boric acid administered in high doses causes an antiproliferative effect on HT-29 cells, whereas the use of boric acid (especially in low doses) together with 5-FU causes the chemotherapeutic effect of the drug to decrease.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ali Tureyen, Sevilay Gunay, Yavuz Erden and Sinan Ince, 2021. Effect of Boric Acid in Colorectal Adenocarcinoma Cell Line and Role on Chemotherapeutic Effect of 5-Fluorouracil. International Journal of Pharmacology, 17: 57-64.
DOI: 10.3923/ijp.2021.57.64
URL: https://scialert.net/abstract/?doi=ijp.2021.57.64
DOI: 10.3923/ijp.2021.57.64
URL: https://scialert.net/abstract/?doi=ijp.2021.57.64
INTRODUCTION
Colorectal cancer is the second most common type of cancer in the world1. Colorectal adenocarcinoma cells are highly resistant to immunological treatments with chemotherapeutic agent or radiation2,3. Chemotherapeutic agents have limited efficacy in the treatment of colon cancer, especially due to internal or acquired drug resistance in cancer cells4. For this reason, to increase the effectiveness of the treatment against the development of resistance in the treatment of colon cancer, there is the use of combined or efficacy enhancing applications of these drugs. Many studies have reported that natural (plant or fungal origin) or chemical (natural or synthesis) compounds affect the effectiveness of chemotherapeutic agents5-7. In a study investigating the efficacy of quercetin, which is a flavonoid, in colorectal cancer cells (HT-29), it was reported that quercetin alone exhibited significant cytotoxic effects on HT-29 cells and this increased the cytotoxic effect of the drug in combination with doxorubicin. However, in some studies, boron compounds such as boric acid, borax and boron esters are stated to be effective in preventive or treatment in cases such as cervical, prostate, chest and lung cancer7-9. However, to date, the mechanism of anticarcinogenic activities of boron compounds has not been fully explained. Barranco and Eckhert7 demonstrated in their study that boric acid administration to DU-145 prostate cancer cells transforms cells into granular, low-volume and flattened cells, however, there is upward β-galactosidase regulation with a decrease in the expression of proliferation-related proteins. Korkmaz et al.10 suggested that boron is effective in cervical cancer cells caused by the human papillomavirus by inhibiting the transformation of the virus. Besides, calcium fructoborate, which is a boron compound, has been reported to be protective against breast cancer cases due to its increased calcitriol levels in the blood11.
This study, it was aimed to determine the effects of boric acid, which is a boron compound on HT-29, having human colon adenocarcinoma cells, alone and in combination with 5-fluorouracil (5-FU) on cytotoxic, genotoxic and cell apoptosis.
MATERIALS AND METHODS
Study area: The study was carried out at the Laboratory of Molecular Biology and Genetics, Faculty of Science, Bartin University, Turkey, from January-March, 2021.
Cell line culture: HT-29 cells (ATCC, USA) were used in the present study. Cells were cultured in 75 cm2 culture flasks using RPMI-1640 medium (10% Fetal bovine serum, 1% penicillin-streptomycin and 1% non-essential amino acid addition). Cells were incubated at 5% CO2 density and 37°C during the whole experimental period (Thermo Forma II CO2 Incubator, USA). The confluent cells were removed with a trypsin-EDTA solution and viability levels were determined using 0.4% trypan blue.
MTT cell viability assay: Following the boric acid (Sigma-Aldrich, B0394, USA) treatments, cell viabilities of HT-29 were determined by 3-(4,5-Dimethylthiazol-2-yl)- 2,5-Diphenyltetrazolium Bromide (MTT) assay. Initially, HT-29 cells were cultured in 96-well plates with a density of 15×103 cells per well. After incubation (24 h), different concentrations of boric acid (1-100 mM) or 5-FU (1-100 μM, reference anticancer drug) were exposed to cells. After this time, HT-29 cells were incubated within 0.5 mg mL–1 MTT (Sigma-Aldrich M2128, USA) solution and the optical densities were determined with a microplate reader (Thermo Multiskan Go, USA) at a wavelength of 570 nm12. The absorbance values obtained from the control groups were averaged and this value was considered as 100% cell viability.
DNA damage analysis: Alkali Comet analysis was performed to DNA damage analysis13. Initially, HT-29 cells were inoculated in 6-well plates and boric acid at a dose of 10 mM and 5-FU at a dose of 100 μM were treated to cells for 24 h. HT-29 cells washed twice with PBS and they were collected and counted. Approximately 10.000 cells/10 μL suspensions were mixed with 80 μL 1% Low Melting Agarose (LMA). The mixture was transferred to slides (1% normal melting agarose-coated) and prepared by closing with a coverslip. Lysis solution (pH: 10) which is including 2.5M NaCl, 1% Triton-X, 10mM Tris, 100mM EDTA and 10% DMSO at 4°C were applied to slides for 1 h. Afterwards, electrophoresis at 25V (max. 300 mA) for 20 min in a horizontal electrophoresis tank was performed on slides. Eventually, slides were washed with neutralization solution (0.4M Tris, pH: 7.5) for 3×5 min and DNA damage into the cell was monitored with a fluorescence microscope (Zeiss Axioscope, Germany) by applying ethidium bromide. Tail DNA (%), tail intensity, tail length (px) and tail moment parameters of 250 cells randomly selected from each group were evaluated and analyzed in TriTek Comet Score.
Western blot analysis: Boric acid was applied into the HT-29 cells grown in 100 mm culture disks and they were incubated 24 h. Total 500 mL RIPA buffer (10 mM Tris-HCl, pH 7.4, 1% Triton X-100, 150 mM NaCl, 0.2 mM Sodium orthovanadate, 1mM EDTA, 0.2 mM PMSF and protease inhibitor cocktail) was added on the HT-29 cells and they were washed twice with PBS and the scraped cells were transferred into tubes on ice. HT-29 cells were lysed with vortex every 5 min for 30 min and then the sonication process was applied for 15 sec. Tubes were centrifuged at 15.000 rpm for 30 min (4°C) and the supernatants were allocated. Protein concentrations of the samples were measured according to the Lowry method14, followed by subjecting to 12% SDS gel electrophoresis. The proteins in the gel were then transferred to the PVDF membrane using the BioRad Trans-Blot Turbo transfer system (BioRad, USA). After blocking the membranes with 5% milk powder, they were treated with primary antibodies overnight at 4°C. After the membranes were washed with TBS-T and they were incubated with secondary antibody (conjugated with HRP) for 1 h at room temperature. Ultimately, membranes treated with electrochemiluminescence solution (ECL Clarity, BioRad, USA) were visualized in the Fusion FX-7 (Vilber, Germany) system. Analysis of the blots was performed in the Image J program. Human anti-β-actin antibody was used as a loading control.
In the study, primary antibodies which are Beclin-1 (1:500, sc-48341), Caspase-3 (1:1000, sc-7272), p53 (1: 1000, sc-126), Bax (1: 1000, sc-20067), Bcl-2 (1: 1000, sc-7382) and β-actin (1: 1000, sc-81178) and anti-mouse secondary antibody (1: 3000, sc-516102) were used and they were obtained from Santa Cruz Biotechnology, Inc.
Statistical analysis: Sigma Plot 12 for the Windows package program was used in the analyses. Quantitative data were summarized as Mean±Standard Deviation (SD). Before making intergroup comparisons of the variables examined, the control for normal distribution and homogeneity of variances were performed. In the case of normal distribution, the difference between group means for the relevant variables was evaluated by one-way analysis of variance (One-Way ANOVA). In cases where normality assumptions were not provided, analyses were performed using the Kruskal-Wallis H test. Values of p<0.05 were considered statistically significant.
RESULTS
Cell viability: The level of viability change (%) in HT-29 cells 24 h after boric acid (1-100 mM) administration is shown in Fig. 1a.
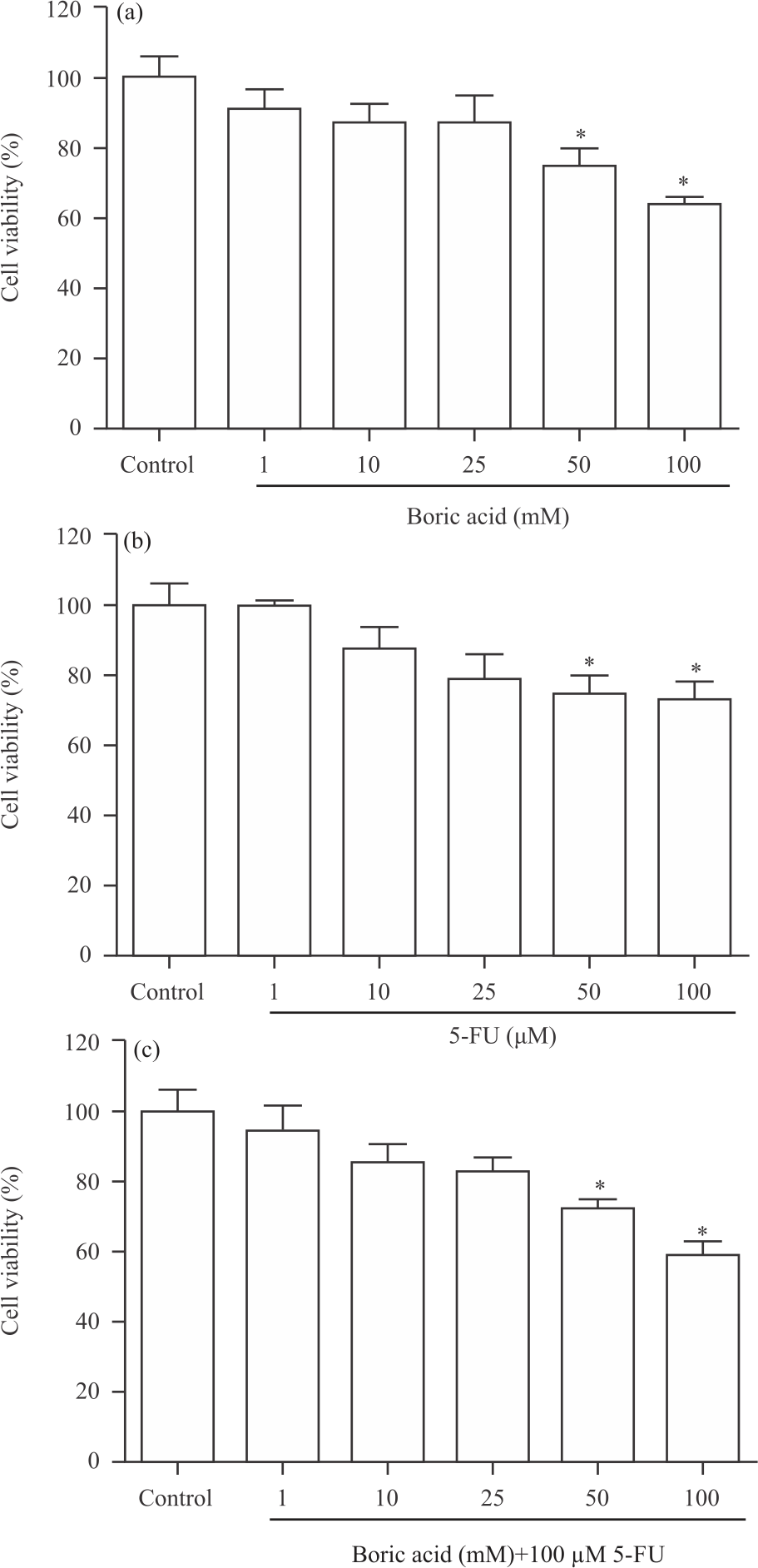 |
| Fig. 1(a-c): | Percentage alteration in HT-29 cell viability after (a) Application of boric acid and (b) 5-fluorouracil (5-FU) It was determined that the treatment of boric acid at doses of 50 and 100 mM reduced cell viability after 24 h in HT-29 cell. Also, 50 and 100 μM of 5-FU treatment were effective in HT-29. Besides, boric acid (50-100 mM)+100 μM 5-FU treatment was decreased cell viability in HT-29 cell (c). *p<0.05 vs. control group |
It was observed that boric acid doses of 1, 10 and 25 mM did not affect cell viability, whereas doses of 50 and 100 mM significantly reduced cell viability (p<0.05). Viability in HT-29 cells decreased by approximately 25 and 36% at 50 and 100 mM doses of boric acid, respectively. Besides, the change in viability in HT-29 cells 24 h after administration of 5-FU, an anticancer drug, at doses of 1-100 μM, is shown in Fig. 1b.
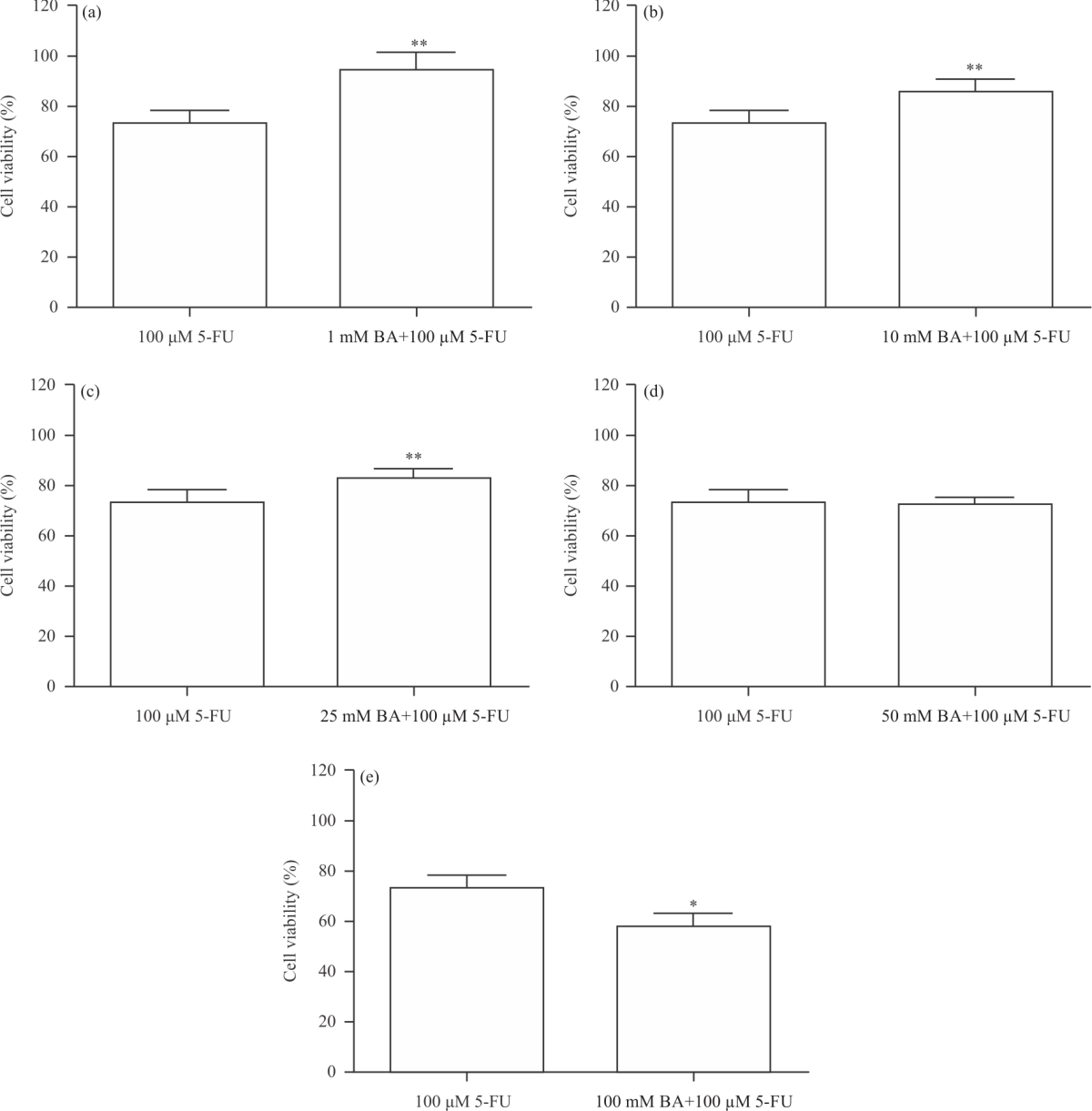 |
| Fig. 2(a-e): | Percentage alteration in HT-29 cell viability after the application of 100 μM 5-fluorouracil (5-FU) and boric acid (BA) (1-100 mM) plus 100 μM 5-FU It was determined that HT-29 cell viability was found to be high-level in BA treatment at doses of 1, 10, and 25 mM compared to 5-FU (p<0.01). On the other hand, 100 mM BA plus 100 μM of 5-FU treatment was reduced cell viability compared to 100 μM of 5-FU treatment in HT-29 cell (p<0.05) |
According to MTT results, 50 and 100 μM administrations of 5-FU resulted in a significant decrease in cell viability (p<0.05). On the other hand, in this study, the anticancer effect of boric acid (1-100 mM) on HT-29 cells was evaluated by combining it with 5-FU (100 μM). It was observed that HT-29 cell viability decreased significantly in 50 and 100 mM+100 μM 5-FU applications of boric acid (Fig. 1c) (p<0.05). Besides as a result of boric acid+5-FU administrations compared to 100 μM 5-FU administrations, it was determined that boric acid 1-25 mM administrations reduced the effects of 5-FU on cancer cell (Fig. 2a-d), p<0.01). However, administration of high dose boric acid (100 mM) with 5-FU showed stronger cytotoxicity than administration of 5-FU alone (Fig. 2e, p<0.05). As a result of cytotoxicity tests, we showed that boric acid administered in low doses did not exhibit a cytotoxic effect and the administration of these doses in combination with 5-FU caused a decrease in the cytotoxicity of 5-FU.
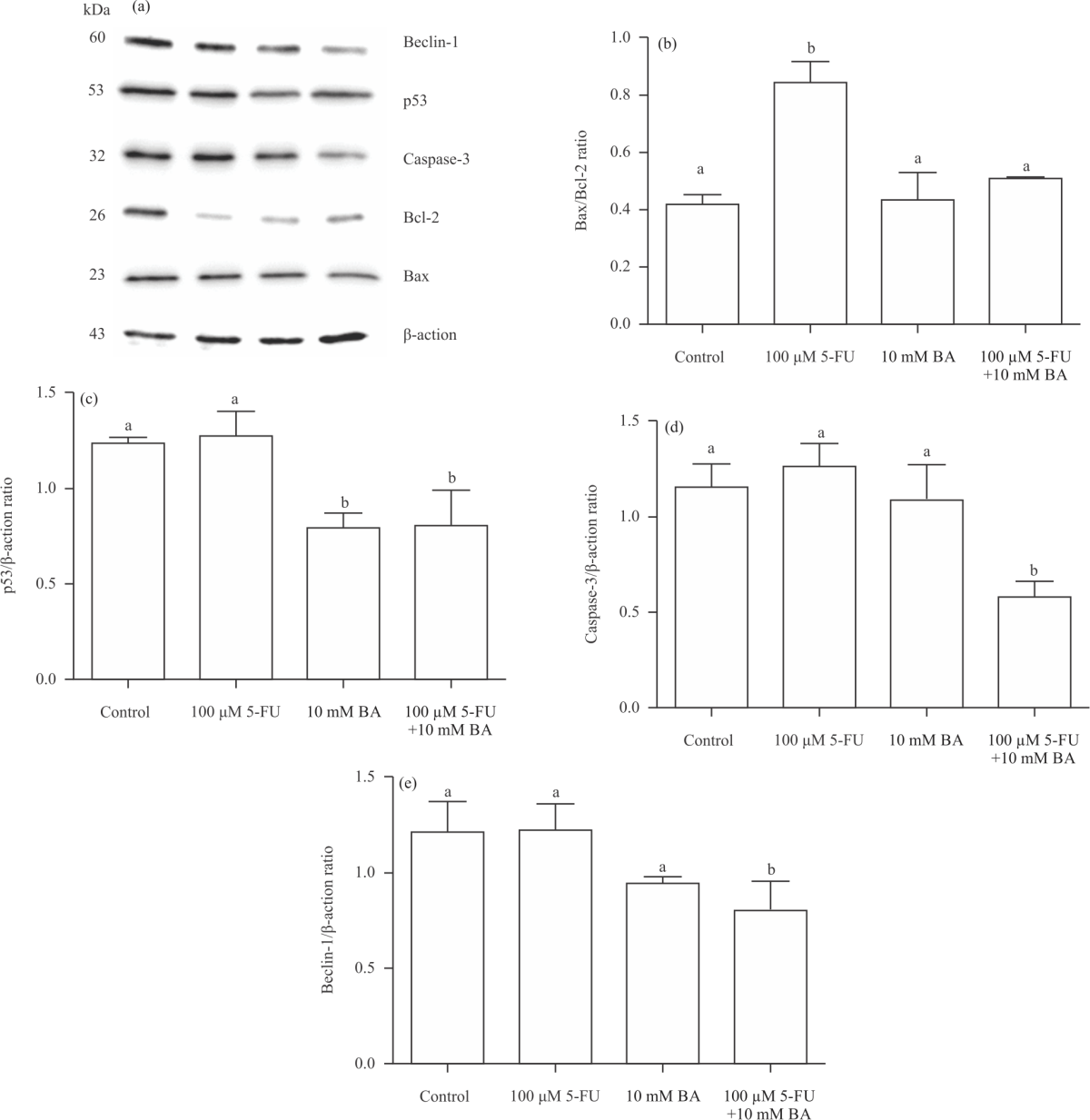 |
| Fig. 3(a-e): | Western blot results of boric acid (BA; 10 mM), 5-fluorouracil (5-FU; 100 μM) and BA (10 mM) plus 5-FU (100 μM) in HT-29 cell (a) Band images obtained from analyzes, (b) The changes in Bax/Bcl-2, (c) p53, (d) Caspase-3 and (e) Beclin-1 expressions after 5-FU, BA and 5-FU+BA applications were shown. It was determined that expressions of these genes did not affect the treatment of BA (10 mM) and BA also decreased the chemotherapeutic effect of 5-FU. a,b: Statistically differences compared to control group (p<0.05) |
To better clarify this situation, the effects of 5-FU applications combined with low dose boric acid in genotoxicity analyses and western blot analyses were investigated. Thus, an answer was sought to the question of how or in what ways boric acid inhibits 5-FU-induced cell cytotoxicity.
DNA damage in HT-29 cells: The level of damage to HT-29 cell DNA 24 h after boric acid and 5-FU administration is shown in Table 1. Tail DNA (%) and tail intensity levels increased significantly 24 h after 5-FU administration compared to the control group (p<0.05).
| Table 1: Comet assay results in HT-29 after the application of 10 mM Boric Acid (BA), 100 μM 5-fluorouracil (5-FU) and BA (10 mM) plus 5-FU (100 μM) | ||||
Tail DNA (%) | Tail length (pixel) | Tail moment | Tail intensity | |
| Control | 31.31 (15.85-51.51) | 22.50 (12.00-44.25) | 16.10 (1.89-35.72) | 82278.00 (35600.25-145270.25) |
| 100 μM 5-FU | 40.87 (30.71-65.95)* | 37.00 (20.50-53.50) | 18.74 (7.25-41.89)¥ | 116751.00 (60879.00-198937.00)* |
| 10 mM BA | 29.83 (17.52-43.02)# | 17.00 (1.00-39.00)# | 5.25 (0.34-18.07) | 52380.00 (33686.50-107952.00)# |
| 100 μM 5-FU+10 mM BA | 35.31 (25.77-67.13)¥ | 39.00 (20.00-46.00)¥ | 23.72 (9.96-39.62)¥ | 75022.00 (40734.00-141679.50)# |
| Values are expressed as median (25-75%), *p<0.05 vs. control, #p<0.05 vs. 5-FU, ¥p<0.05 vs. BA | ||||
In boric acid applied groups, tail DNA (%), tail length and tail intensity levels showed a significant decrease compared to the 5-FU group (p<0.05). In the groups where 5-FU and boric acid were applied together, tail DNA (%), tail length and tail moment levels were quite high compared to the group where boric acid was applied only (p<0.05). These results showed that 5-FU exhibited genotoxic effects on HT-29 cells as expected. However, only low dose (10 mM) boric acid administration did not have a genotoxic effect on DNA compared to the control group. Moreover, boric acid caused a significant decrease in tail intensity level when administered with 5-FU compared to 5-FU (p<0.05). These results show that boric acid alone does not exhibit a genotoxic effect on DNA, whereas it can partially prevent DNA damage after 5-FU administration.
Apoptosis in HT-29 cell: Western blot analyses were performed to understand the possible molecular basis of the cell death mechanism in the study. Studies targeting apoptosis and autophagy in cancer cells are important in increasing the effectiveness of treatment. Changes in apoptosis and autophagy markers in HT-29 cells after boric acid administration are shown in Fig. 3a.
Analyses revealed that the Bax/Bcl-2 protein ratio, which is an important marker of the intrinsic apoptotic pathway after boric acid and 5-FU administration, showed a significant increase in the 5-FU group (Fig. 3b, p<0.05). On the other hand, this level was found to be similar to the control group in the boric acid and 5-FU+boric acid groups. 5-FU administration did not cause a significant change in p53 and caspase-3 protein expressions. While p53 expression decreased in the boric acid applied group compared to the control group (Fig. 3c, p<0.05), no significant change was found in the caspase-3 protein level. After 5-FU+boric acid administration, a significant decrease was observed in p53, caspase-3 and Beclin-1 expressions (Fig. 3c-e, p<0.05). Moreover, boric acid applied was found to reduce the p53 level in HT-29 cells compared to the control group. These results show that boric acid does not induce apoptosis in HT-29 cells and also reduces the chemotherapeutic efficacy of 5-FU.
DISCUSSION
In the study, boric acid and 5-FU significantly reduced HT-29 cell viability at high doses. The low dose of boric acid treatment did not affect cell viability, genotoxicity and apoptotic process. Also, boric acid administered with 5-FU partially prevented the chemotherapeutic efficacy of 5-FU by reducing its genotoxic effect. It has been stated in vitro studies that different cancer cell lines are used in which the boron has shown an inhibitory effect on cancer cells in recent years. Boric acid (0-12.5 mM) has been reported to prevent cell proliferation by acting as high doses of oxidants on DU-145 human prostate cancer cells15. Again, boric acid has been reported to cause a gradual decrease in the growth and survival of human hepatocarcinoma (HepG2) cells depending on the dose (5 mM and above)16. Besides, it has been reported that boric acid over 5 mM on Hek and HeLa cell lines cause a significant decrease in cell proliferation17. On the other hand, in a study showing that boric acid reduces cytotoxicity caused by toxic substances on normal cell lines, boric acid (2.5, 5 and 10 mM) has been reported to reduce cytotoxicity decreasing with formaldehyde on epithelial lung cell lines (A549) at increased doses18. Similar to the cytotoxic effect seen in other cancer cell lines, the administration of 50 and 100 mM concentrations of boric acid decreased the viability of HT-29 cells in this study, which showed that high doses of boric acid had an antiproliferative effect. Besides, it was shown that high dose boric acid affected increasing the cytotoxicity of 5-FU when boric acid was administered in combination with 5-FU, whose efficacy was known in cancer cells.
In vitro and in vivo studies have reported that boron compounds prevent genotoxic effects caused by chemical agents19-21. Boric acid (2.5 or 5 mg L–1) has been reported to have a preventive effect against sibling chromatid exchange and genotoxicity in micronuclei in paclitaxel (10 or 20 μg L–1) human blood cultures, which is used in the treatment of cancer. Besides, in the study, it was stated that boric acid did not cause a genotoxic effect when administered alone at the specified doses20. Similarly, it was emphasized that boric acid administration at low levels (2.5 and 10 μM) against cadmium
chloride and lead chloride-induced genotoxicity in V79 hamster lung fibroblast cell cultures has a protective effect21. In another in vitro study, Tombuloglu et al.16 found that boric acid administered at 24 mM dose with genotoxicity tests and microarray hybridizations caused serious DNA damage in HepG2 cells and there was a significant change in the transcriptomic profile. In this study, we revealed that 10 mM boric acid application did not have a genotoxic effect on HT-29 cell lines. Besides, our study results showed that boric acid administered at a concentration of 10 mM on HT-29 cells can partially prevent DNA damage after 5-FU administration. The results show that boric acid can play different cellular roles depending on the dose, maintains cell viability at low doses and exhibits cytotoxic effect at high doses as stated in the current literature.
Apoptosis plays an important role in the normal functions of cells. In contrast, cancer cells can escape apoptosis, which is expected to occur under normal conditions, with some mechanisms they develop. This increases the retention of cancer cells in the environment and allows cancer to spread rapidly. Thus, it is important to determine the roles of the substances to be used against cancer in cell signalling pathways22,23. When studies showing that boric acid causes apoptosis are examined, it is stated that boric acid administration below 1 mM causes a decrease in mRNA and protein levels of CHOP, a proapoptotic gene24 and again 10 μM boric acid causes growth inhibition in the prostate cancer cell line without any visible sign of apoptosis25. However, caspase-3, cytochrome C somatic and Bax which are apoptotic markers in DU-145 cells given boric acid at concentrations of 6.15 mM and higher have been reported to be upregulated15. In a study, Wei et al.26 administered borax, which is a boron compound in the concentration range of 1, 2 and 4 mM, to HepG2 cell line for up to 24 h, reported an increase in tumour suppressor p53 and proapoptotic Bax mRNA expression, whereas a decrease in mRNA expression of apoptosis regulator Bcl-2 was observed and expression levels of these proteins were also observed with western blotting. Similarly, Alexandre et al.16 reported that 24 mM boric acid administration over 24 h shaped a high number of transcription downregulation in the DNA synthesis and cell cycle of HepG2 cells and relatively low transcription up-regulation in the p53 signal pathway. Contrary to these studies, in a study investigating the effect of boric acid on Ehrlich Ascites Carcinoma (EAC), it was reported to significantly increase the total number and viability of peritoneal EAC cells in mice given 62.5, 125 and 250 mg kg–1 boric acid for 5 days27. In this study, boric acid did not affect
Beclin-1, caspase-3, Bcl-2 and Bax protein expression in HT-29 cells and significantly reduced p53 expression and apoptotic protein. This shows that a 10 mM dose of boric acid does not cause cell death through apoptosis in HT-29 cell lines. It also shows that HT-29 cell death after 5-FU administration is mediated by increased proapoptotic Bax protein. On the other hand, the similarity of Bax/Bcl-2 expression to the control group in the 5-FU+boric acid group and the decrease determined in Beclin-1 expression reveals that boric acid can inhibit the effects of 5-FU and mediate cell survival.
CONCLUSION
In conclusion, our study results showed that boric acid can cause cellular survival or cytotoxicity depending on the dose. Moreover, we can say that boric acid administered with 5-FU in low doses reduces the cytotoxicity and genotoxicity caused by 5-FU. Furthermore, boric acid administered in lower doses partially prevented the efficacy displayed by 5-FU on cellular death processes.
SIGNIFICANCE STATEMENT
This study revealed that boric acid had a cytotoxic effect in high doses, whereas administration with 5-FU weakened its cytotoxic effect on HT-29 cells. In this study, it can be reached to the approach that concomitant administration of cytotoxic drugs used together with boric acid in colon cancer treatment may delay the desired outcome.
REFERENCES
- Jiang, R. and B. Lönnerdal, 2017. Bovine lactoferrin and lactoferricin exert antitumor activities on human colorectal cancer cells (HT-29) by activating various signaling pathways. Biochem. Cell Biol., 95: 99-109.
CrossRefDirect Link - De Maria, S., I. Scognamiglio, A. Lombardi, N. Amodio and M. Caraglia et al., 2013. Polydatin, a natural precursor of resveratrol, induces cell cycle arrest and differentiation of human colorectal Caco-2 cell. J. Transl. Med., 11: 1-11.
CrossRefDirect Link - Todaro, M., M.P. Alea, A.B. Di Stefano, P. Cammareri and L. Vermeulen et al., 2007. Colon cancer stem cells dictate tumor growth and resist cell death by production of interleukin‐4. Cell Stem Cell, 1: 389-402.
CrossRefDirect Link - Gorlick, R. and J.R. Bertino, 1999. Drug resistance in colon cancer. Semin. Oncol., 26: 606-611.
Direct Link - Atashpour, S., S. Fouladdel, T.K. Movahhed, E. Barzegar and M.H. Ghahremani, 2015. Quercetin induces cell cycle arrest and apoptosis in CD133+ cancer stem cells of human colorectal HT29 cancer cell line and enhances anticancer effects of doxorubicin. Iran J. Basic Med. Sci., 18: 635-643.
Direct Link - Redondo-Blanco, S., J. Fernandez, I. Gutierrez-del-Rio, C.J. Villar and F. Lombo, 2017. New insights toward colorectal cancer chemotherapy using natural bioactive compounds. Front. Pharmacol., Vol. 8.
CrossRefDirect Link - Barranco, W.T. and C.D. Eckhert, 2006. Cellular changes in boric acid-treated DU-145 prostate cancer cells. Br. J. Cancer, 94: 884-890.
CrossRefDirect Link - Scorei, R., R. Ciubar, C.M. Ciofrangeanu, V. Mitran, A. Cimpean and D. Iordachescu, 2008. Comparative effects of boric acid and calcium fructoborate on breast cancer cells. Biol. Trace Elem. Res., 122: 197-205.
CrossRefDirect Link - Simsek, F., S. Inan and M. Korkmaz, 2019. An in vitro study in which new boron derivatives maybe an option for breast cancer treatment. Eurasian J. Med. Oncol., 3: 22-27.
CrossRefDirect Link - Korkmaz, M., E. Uzgoren, S. Bakirdere, F. Aydin and O.Y. Ataman, 2007. Effects of dietary boron on cervical cytopathology and on micronucleus frequency in exfoliated buccal cells. Environ. Toxicol., 22: 17-25.
Direct Link - Scorei, R., 2012. Is boron a prebiotic element? A mini-review of the essentiality of boron for the appearance of life on earth. Orig. Life Evol. Biosph., 42: 3-17.
CrossRefDirect Link - Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods, 65: 55-63.
CrossRefPubMedDirect Link - Singh, N.P., M.T. McCoy, R.R. Tice and E.L. Schneider, 1988. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res., 175: 184-191.
CrossRefPubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Hacioglu, C., F. Kar, S. Kacar, V. Sahinturk and G. Kanbak, 2020. High concentrations of boric acid trigger concentration-dependent oxidative stress, apoptotic pathways and morphological alterations in DU-145 human prostate cancer cell line. Biol. Trace Elem. Res., 193: 400-409.
CrossRefDirect Link - Tombuloglu, A., H. Copoglu, Y. Aydin-Son and N.T. Guray, 2020. In vitro effects of boric acid on human liver hepatoma cell line (HepG2) at the half-maximal inhibitory concentration. J. Trace Elem. Med. Biol., Vol. 62.
CrossRefDirect Link - Park, M., Q. Li, N. Shcheynikov, W. Zeng and S. Muallem, 2004. NaBC1 is a ubiquitous electrogenic Na+-coupled borate transporter essential for cellular boron homeostasis and cell growth and proliferation. Mol. Cell, 16: 331-341.
CrossRefDirect Link - Arslan-Acaroz, D. and N. Bayşu-Sozbilir, 2020. Ameliorative effect of boric acid against formaldehyde-induced oxidative stress in A549 cell lines. Environ. Sci. Pollut. Res., 27: 4067-4074.
CrossRefDirect Link - Ince, S., I. Kucukkurt, H.H. Demirel, D.A. Acaroz, E. Akbel and I.H. Cigerci, 2014. Protective effects of boron on cyclophosphamide induced lipid peroxidation and genotoxicity in rats. Chemosphere, 108: 197-204.
CrossRefDirect Link - Turkez, H., A. Tatar, A. Hacimuftuoglu and E. Ozdemir, 2010. Boric acid as a protector against paclitaxel genotoxicity. Acta Biochim. Pol., 57: 95-97.
CrossRefPubMedDirect Link - Ustundag, A., C. Behm, W. Follmann, Y. Duydu and G.H. Degen, 2014. Protective effect of boric acid on lead- and cadmium-induced genotoxicity in V79 cells. Arch. Toxicol., 88: 1281-1289.
CrossRefDirect Link - Sharma, A., L.H. Boise and M. Shanmugam, 2019. Cancer metabolism and the evasion of apoptotic cell death. Cancers, Vol. 11.
CrossRefDirect Link - Erden, Y., 2020. Capsanthin stimulates the mitochondrial apoptosis-mediated cell death, following DNA damage in MCF-7 cells. Nutr. Cancer, 73: 662-670.
CrossRefDirect Link - Barranco, W.T. and C.D. Eckhert, 2004. Boric acid inhibits human prostate cancer cell proliferation. Cancer Lett., 216: 21-29.
CrossRefDirect Link - Kobylewski, S.E., K.A. Henderson, K.E. Yamada and C.D. Eckhert, 2017. Activation of the EIF2α/ATF4 and ATF6 pathways in DU-145 cells by boric acid at the concentration reported in men at the US mean boron intake. Biol. Trace Elem. Res., 176: 278-293.
CrossRefDirect Link - Wei, Y., F.J. Yuan, W.B. Zhou, L. Wu, L. Chen, J.J. Wang and Y.S. Zhang, 2016. Borax-induced apoptosis in HepG2 cells involves p53, Bcl-2, and Bax. Genet. Mol. Res., Vol. 15.
CrossRefDirect Link - Qureshi, S., O.A. Al-Shabanah, M.M. Al-Harbi, A.M. Al-Bekairi and M. Raza, 2001. Boric acid enhances in vivo Ehrlich ascites carcinoma cell proliferation in Swiss albino mice. Toxicology, 165: 1-11.
CrossRefDirect Link