
Benjaporn Buranrat
Faculty of Medicine, Mahasarakham University, Talad, Mueang, Maha Sarakham 44000, Thailand
LiveDNA: 66.35857
International Journal of Pharmacology
Year: 2021 | Volume: 17 | Issue: 5 | Page No.: 339-349







ABSTRACT
Background and Objective: Careya arborea (CA) is a plant or vegetable found in Thailand and consumed as a fresh vegetable that reported anticancer effect. The action of CA on growth, apoptosis and migration of cervical HeLa cancer cells and underlying mechanism has not been explored. So, current article studied the human HeLa cervical cancer cell to examine the anticancer action of DW and ethanolic CA extract. Materials and Methods: HeLa cell was treated with the different doses of CA extract (DW and 95% ethanol) and the viability was determined by SRB and cell cycle distribution method. Apoptosis was determined by AO/EB staining, Annexin V-FITC and PI staining, ROS production. Cells migration was examined by wound healing method. Results: Treatment with both of CA extracts promoted HeLa cell death correlating with suppression the colony formation by dose-dependent manner. The ethanolic extract had more potency than DW extract with lower IC50 values of SRB and colony formation assay. CA extracts significantly inhibited the cancer cell distribution at G0/G1 phase and stimulated apoptosis after detecting with AO/EB staining and confirming with flow cytometry. Increased late apoptotic cell death was indicated after incubating with CA extract and the percentage were 66.2% for DW extract and 35% for ethanolic extract. The mechanism of CA extract on cell death and apoptosis were detected about reduction of mitochondrial function and induction of ROS formation. Lastly, CA suppressed the cancer cells migration in a concentration-dependent manner, especially high CA dose. Conclusion: Treatment with CA extract tends to increase death, stimulate of apoptosis and reduce migration capability of cervical cancer cells through increasing ROS formation and attenuating mitochondrial function.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Benjaporn Buranrat, 2021. Growth Inhibition, Apoptosis Induction and Migratory Suppression by Careya arborea Leaf Extract in HeLa Cervical Cells. International Journal of Pharmacology, 17: 339-349.
DOI: 10.3923/ijp.2021.339.349
URL: https://scialert.net/abstract/?doi=ijp.2021.339.349
DOI: 10.3923/ijp.2021.339.349
URL: https://scialert.net/abstract/?doi=ijp.2021.339.349
INTRODUCTION
Cervical cancer is one of the frequently most common types of gynecological cancers and it is also the fourth cancer that leading female mortality1. The data has been reported that each year found cervical cancer approximately ~500,000 of new cases and the statistical analysis was found more than 250,000 related deaths worldwide2. The incidence rate decreased in developed countries; however still high in developing countries. Early diagnosis, early surgical procedure and early started with chemotherapeutic agents enable patients to receive effective treatments, nevertheless, prognosis of cervical cancer is still poor3. Adjuvant chemotherapeutic drugs following resection of cervical cancer goal to suppress the cancer cells proliferation, but the limitation of treatment is that normal cells are toxic, killed and serious complications are occurred. Cervical cancer patient’s death may from cancer aggression, progression, angiogenesis, metastasis and resistance4. Hence, we urgently need to find new agents that have low toxicity and high efficacy for cervical cancer treatment and prevention.
Over the last few years, several research data has been performed to explore the capability anticancer properties of natural products or plants or vegetables, which are considered as low toxicity, fewer side effects and high efficacy as well5. Our previous study demonstrated that Careya arborea (CA) had more efficacies against human breast cancer cells by attenuating cancer cell proliferation, stimulating apoptosis and inhibiting migration6. The CA efficacies on several cancer cell types are more interested including cervical cancer. CA is belonging to Barringtonieacea family and frequently found in the North-east part of Thailand. Young leaves and flowers of CA usually cooked as a soup and consumed daily as a fresh vegetable. It has a bitter taste and slightly sour. This plant has been considerably examined and active constituents from several parts were reported to include tannins, coumarin, triterpenoids, flavonoid, sterols, saponins, betulinic acid, betulin and lupeol7.
Amusingly, different each parts of CA are demonstrated to be useful defeat different diseases and had several pharmacological effects. It is conventionally used in anticancer, antiinflammation, antianthelmintic, bronchitis, skin disease, diarrhea, dyspepsia and ulcer7. However, CA actions on anticancer activities are less information in all types of cancer. Our previous study found that edible leaf of CA defeated the breast cancer MCF-7 cells by attenuating cells proliferation, stimulating apoptosis and also suppressing migration. Growth inhibition was carried out through induction of ROS generation, activation of caspase 3 activity and migratory inhibition was downregulating the expression of MMP 2 and MMP 96. In vivo study demonstrated that CA reduced the volume of solid tumor that was injected by lymphoma cells8.
Nevertheless, so far less anticancer activity has been testified from CA. Current article studied the human HeLa cervical cancer cell to examine the anticancer action of DW and ethanolic CA extract.
MATERIALS AND METHODS
This study was conducted at Mahasarakham University, Thailand in 2019-2020.
Plant extraction: Young CA leaves were obtained from Udon Thani Province, Thailand in 2019, further identified and collected at the Pharmaceutical Laboratory Service Center at Prince of Songkla University (Herbarium No. SKP022031901). The extraction procedure, edible young leaves (0.5 kg) were dried at 60°C for 24 hrs and mashed to small pieces. Then, leaves were divided to two groups, first group was boiled in DW (0.25 kg) and second group was macerated (0.25 kg) for 7 days in 95% ethanol (1.5 L), filtered, evaporated, lyophilized and stored in a freezer (-20°C). The yield was 15.12 and 8.23% for DW and ethanolic extract, respectively.
Cell culture and cytotoxic method: HeLa cervical cancer cells was cultured in 10% FBS of DMEM medium with 1% antibiotic (Penicillin and streptomycin). All chemicals were obtained from Gibco BRL Life Technologies (Grand Island, NY, USA).
DW and ethanolic extract of CA were dissolved in 100% DMSO for primary stock. The action of CA extract on HeLa cells growth was examined by the sulforhodamine B (SRB). Briefly, HeLa cells (1×104 cells/well) were plated overnight in 96 well-cultured plate and then incubated with different concentration of CA (0-250 μg mL–1) for 24, 48, 72 hrs. End of incubation periods, the cancer cells were exposed to 10% trichloroacetic acid (TCA), washed with DW for three times, added with 0.4% sulforhodamine B (SRB) for 30 min at room temperature and solubilized with 200 μL 10 mM Tris base. The optical density value was read at 540 nm by a microplate reader.
Colony formation method: The action of CA extract on HeLa cells replication was examined by colony formation.
For colony formation method, 500 cells were cultured in triplicate into 6-well cultured plate for 24 hrs. Next day, the different doses of CA extracts (0-250 μg mL–1) were added into each well for 24 hrs and washed the cells with PBS buffer for two times. HeLa cells were cultured for another ten days in CO2 incubator, during which period the medium was changed every two-three days. The colonies of cancer cells were then added to 100% methanol for 15 min at -20°C, stained with 0.5% crystal violet, dried and counted the colonies.
Cell cycle distribution method: The action of CA extract on HeLa cells cycle distribution was examined by the flow cytometry method.
For cell cycle distribution analysis, 2.5×105 cells/mL were plated onto 6-well cultured plate for 24 hrs and then various doses of CA extracts (0-250 μg mL–1) were added in each well for 24 hrs incubation period. After cells were trypsinized, washed with PBS buffer, then fixed with ice-cold 70% ethanol at -20°C. Next, the cancer cells were exposed to PI solution (Cat No. 550825, BD Biosciences, CA, USA) for 30 min at 4°C and analyzed the phases of cell cycle by flow cytometer (BD Biosciences, CA, USA) using BD Accuri C6 Plus software and fluorescent signals were displayed as histograms. Gated cells were manually categorized into cell-cycle stages.
Acridine orange/Ethidium bromide (AO/EB) staining method: The action of CA treatment on apoptosis was determined by the AO/EB staining. The morphological changes of cancer cells apoptosis were detected by AO/EB fluorescent dye. The HeLa cancer cells (1×104 cells/well) were seeded into 96-well cultured plate for 24 hrs and exposed to various doses of CA extract (0-250 μg mL–1) for 24 hrs. The HeLa cells were added 10 μL of the dye mixture containing AO (1 μg mL–1) and EB (1 μg mL–1) in 1:1 ratio and incubated 15 min at RT in dark. The cultured plate was determined immediately under the florescence microscope (20x magnification) and images were captured.
Apoptosis by flow cytometric method: The action of CA treatment on apoptosis was determined by PI and Annexin V-FITC double staining method. For apoptosis, HeLa cancer cells (2.5×105 cells/well) were plated into 6-well cultured plate for 24 hrs and exposed to various doses of CA extract (0-250 μg mL–1) for 24 hrs. HeLa cells were harvested by trypsin, washed by PBS buffer and resuspended with 1× apoptosis binding assay buffer. Apoptotic cells were identified by annexin V-FITC and PI solution (Cat No. 558547, BD Biosciences, CA, USA) and then incubated at room temperature for 15 min. Apoptosis was detected by flow cytometer (BD Biosciences, CA, USA) within 60 min using BD Accuri C6 Plus software.
Mitochondrial membrane potential by flow cytometric method: The action of CA treatment on mitochondrial function was determined by JC-1 staining method. For mitochondrial function, HeLa cancer cells (2.5×105 cells/well) were seeded into 6-well cultured plate for 24 hrs and treated with various doses of CA extract (0-250 μg mL–1) for 24 hrs. The cancer cells were harvested, washed and collected the pellet cancer cells. The cells were added in 100 mL medium containing 5 mL JC-1 assay reagents for 30 min at 37°C in the dark and finally added 400 mL of DMEM medium. Mitochondrial membrane potential was determined by flow cytometric analysis (BD Biosciences, CA, USA) using BD Accuri C6 Plus software.
Reactive oxygen species (ROS) by flow cytometric method: The action of CA treatment on ROS formation was determined by DCF-DA staining method. For ROS formation, HeLa cancer cells (2.5×105 cells/well) were plated into 6-well cultured plate for 24 hrs and exposed to various doses of CA extract (0-250 μg mL–1) for 24 hrs. Then pellet cells were collected and added the new complete DMEM medium with 25 mM 2,7-Dichlorofluorescin diacetate, DCF-DA (Cat. No. D6883, Sigma Merck KGaA, Darmstadt, Germany) for 30 min at 37°C in the dark. ROS generation was determined by flow cytometry (BD Biosciences, CA, USA) using BD Accuri C6 Plus software and fluorescent signals were displayed as histograms.
Cell migration method: The action of CA treatment on migratory ability was determined by the wound healing method. For cell migration, HeLa cancer cells (2.5×105 cells/well) were plated into 24-well cultured plate for 24 hrs, scratched cells by 0.2 mL pipette tips and added the new medium with various doses of CA extract (0-250 μg mL–1) for 48 hrs. Wound closure was measured by the width length of the wounds using Image J software (NIH, Bethesda, MD, USA) at 0 and 48 hrs.
Statistical analysis: All data was received from at three independent experiments (n = 3) and represented as the mean±SE. Using Student's t-test to compare between treatment and control groups and set the data statistically significant difference less than 0.05.
RESULTS
CA extracts on cell growth and cell replication: Cytotoxic action of CA extract on HeLa cells was assessed by examining its action on cell proliferation using the SRB and colony formation method. The data demonstrated that both of CA extract (DW and ethanol) exhibited a cytotoxicity effect against HeLa cells in a concentration- and time-dependent manner with the IC50 values of 69.87±13.38 and 37.33±6.53 μg mL–1 for DW and ethanolic extract at 72 hrs, respectively (Fig. 1a-b, Table 1). Interestingly, ethanolic extract had more potency than DW extract on HeLa cell death.
To quantify cell replication upon CA treatment, colony formation method was carried out. The data indicated that, colonies formation of HeLa cells had undergone inhibition after exposing to both CA extract in a concentration-dependent manner (Fig. 1a-b, Table 2). In conclusion, CA extract strongly inhibits cell replication in HeLa cervical cancer cells.
CA extracts on cell cycle distribution: To explore the CA effects on cell proliferative attenuation and the cell cycle distribution was used by using flow cytometric analysis. The data demonstrated that the DW extract of CA increased the cell cycle at G0/G1 phase in a dose-dependent manner (Fig. 2a) with the highest potency at the dose of 250 μg mL–1 after treatment for 24 hrs. Correspondingly, the HeLa cells after exposing with ethanolic extract slightly accumulate at G0/G1 phase, mostly at 250 μg mL–1 of CA extract (Fig. 2b). Additionally, an increase in G0/G1 phase, which were predicted as apoptotic/necrotic fractions was occurred and the arrested was significantly detected in HeLa cells after treating with CA extract at the dose of 250 μg mL–1.
CA extracts on cell apoptosis: To measure apoptosis upon CA extract effect, AO/EB staining, PI and Annexin V-FITC double staining were carried out. AO/EB staining confirmed that, HeLa cells number had undergone reduction after exposing to DW and ethanol of CA extract in a concentration-dependent manner (Fig. 3a-b). For DW extract, the cell morphology did not alter; however, ethanolic had more change of the cell morphology and significant to stain the red color as necrosis detected.
For PI and Annexin V-FITC double staining demonstrated that, DW extract showed the late apoptosis induction by dose-dependent manner from 4.0, 8.0, 11.2 and 66.2% of 0, 50, 100 and 250 μg mL–1, respectively. Ethanolic extract treatment demonstrated that this extract CA extract can emits auto-fluorescent, however, we detected the trend of late apoptosis induction by dose-dependent manner from 11.1, 13.5, 21.6 and 35% (Fig. 3a-b). These results recommended that CA extract strongly inhibits cell growth and stimulates late apoptosis cell death pathway in cervical cancer cells.
Further, to determine the mechanism of action of CA extract on HeLa cells and Mitochondrial Membrane Potential (MMP) and ROS formation were carried out. MMP was used to detect the mitochondrial function and the population of cells moved to lower right quadrant it means the loss of mitochondrial function or dysfunction was found. From the results indicated that DW extract induced the loss of mitochondrial function from 0.6, 9.0, 9.5 and 17.0% from 0, 50, 100 and 250 μg mL–1, respectively. Similarly, ethanolic extract had higher effects than DW extract by induction of mitochondrial dysfunction by dose-dependent manner from 7.8, 8.4, 13.2 and 53.7%. CA induced HeLa cell apoptosis via mitochondrial dysfunction (Fig. 4a-b).
| Table 1: IC50 values and Emax of CA extract on HeLa cell viability | |||
| Groups of treatment | Incubation periods (hrs) | IC50 (μg mL–1) | Emax |
| DW extract | 24 | 269.43±52.82 | 50.64±7.78 |
48 | 197.17±1.81* | 52.09±4.22 | |
72 | 69.87±13.38* | 70.21±1.59* | |
| Ethanolic extract | 24 | 147.86±32.26 | 59.80±3.02 |
48 | 94.73±18.25* | 74.76±3.95* | |
72 | 37.33±6.53* | 85.47±4.17* | |
*p<0.05 when compared with each 24 hrs incubation time. Values were Mean±SE | |||
| Table 2: IC50 values and Emax of CA extract on HeLa cell replication | |||
| Assay | Groups of treatment | IC50 (μg mL–1) | Emax |
| Colony formation method | DW extract | 78.38±8.34 | 95.52±4.47 |
| Ethanolic extract | 34.79±2.52 | 100.00±0.00 | |
| Cell migration method | DW extract | 394.37±85.22 | 52.11±8.38 |
| Ethanolic extract | 124.60±5.74* | 90.10±5.74* | |
| *p<0.05 significant when compared with DW extract. Values were Mean±SE | |||
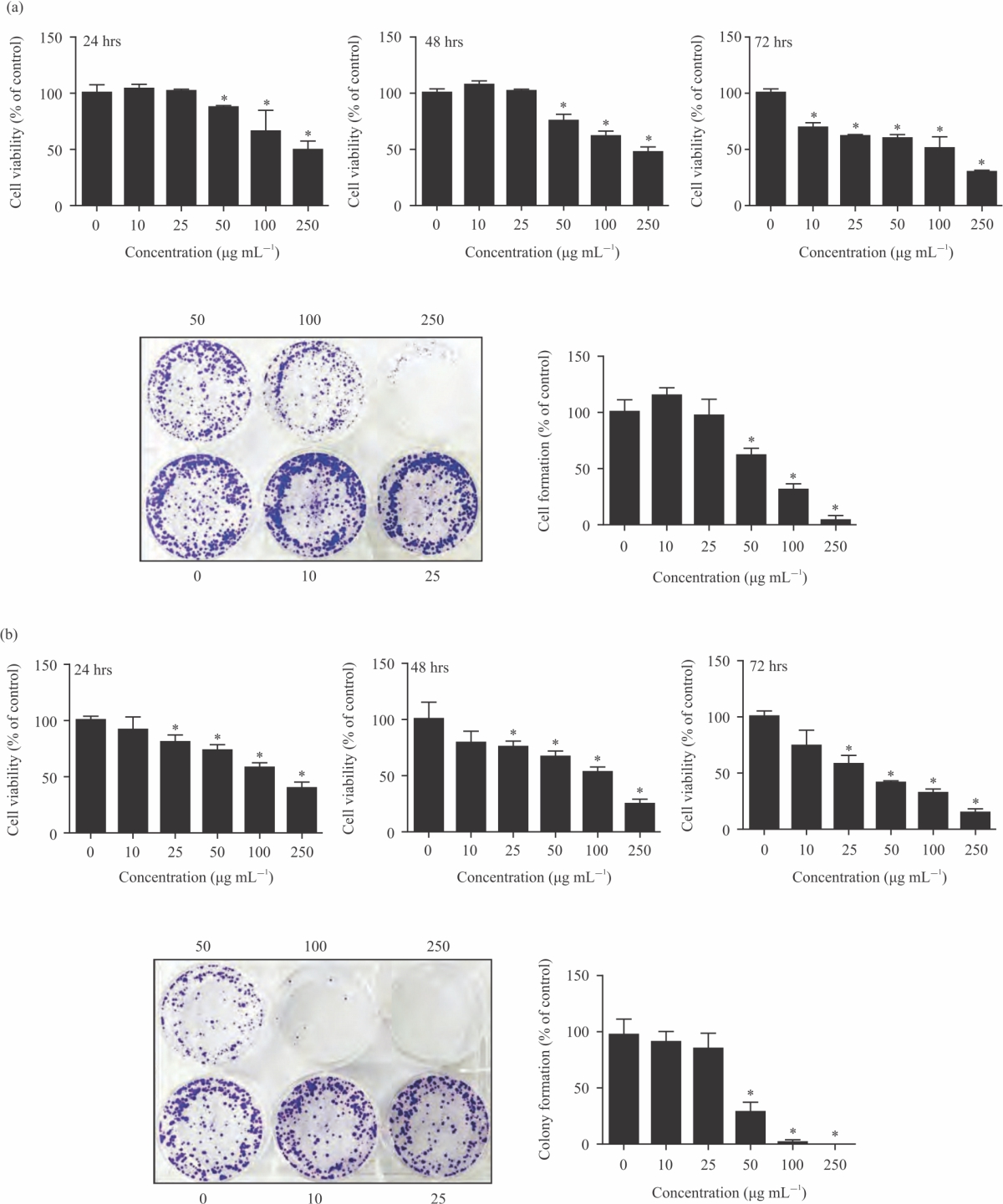 |
| Fig. 1(a-b): | CA effects on cell viability and colony formation. (a-b) The HeLa cancer cells were incubated with DW and ethanolic of CA extracts (0-250 μg mL–1) for 24-72 hrs and then measured cell viability by SRB assay. The cells were incubated with DW and ethanolic CA extracts (0-250 μg mL–1) for 24 hrs (a-b), then cultured further for 15 days, stained with crystal violet and counted Values are expressed as mean±SE. *p<0.05 was significant from control group |
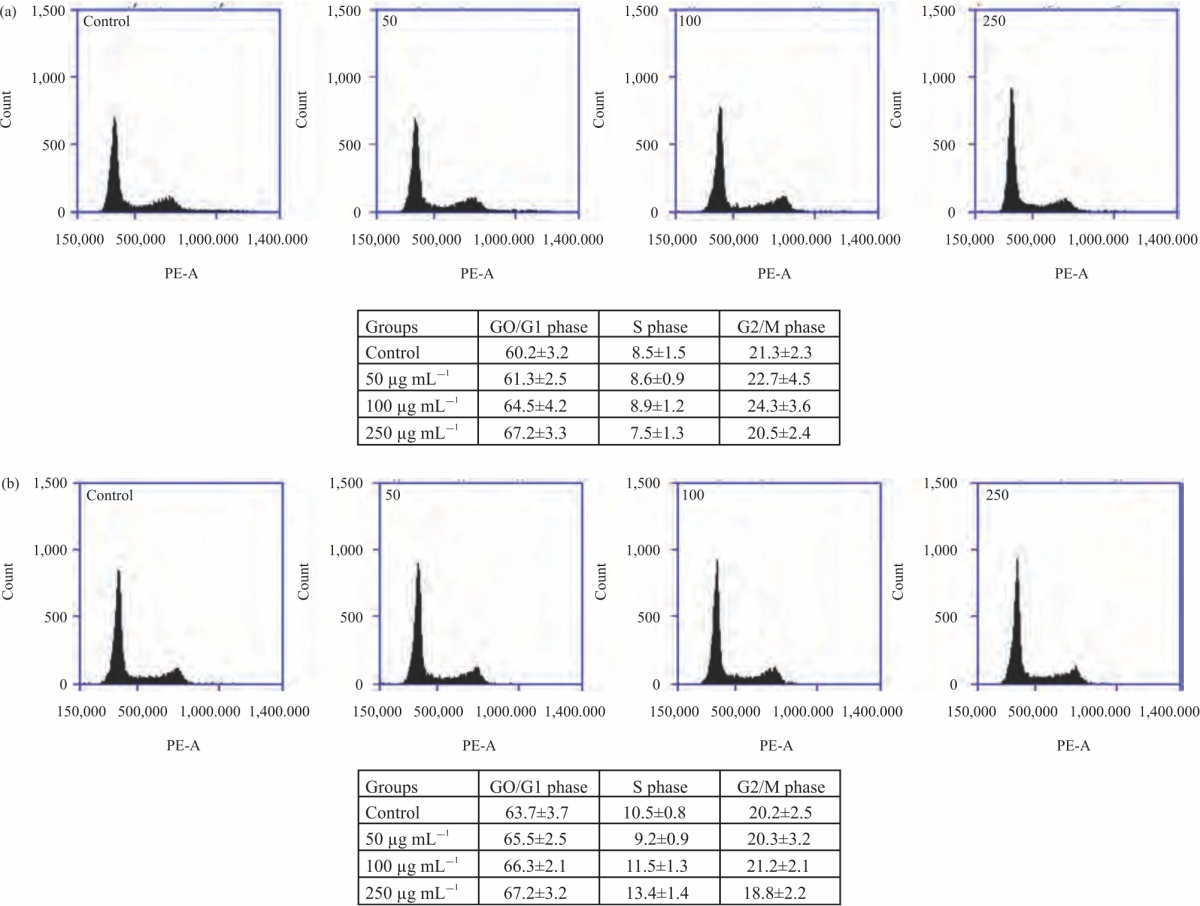 |
| Fig. 2(a-b): | CA effects on cell cycle arrest. (a-b) The HeLa cancer cells were incubated with DW and ethanolic CA extracts (0-250 μg mL–1) for 24 hrs and then measured cell cycle arrest by flow cytometric analysis Values are expressed as Mean±SE. *p<0.05 was significant from control group |
ROS play important roles in mitochondria-mediated apoptosis pathway. Effect of CA extract on ROS level in HeLa cells was measured by DCF-DA staining. Our results showed that DW extract increased ROS levels from 2.0, 4.1 and 5.3%, 7.7 of 0, 50, 100 and 250 μg mL–1 after 24 hrs treatment, respectively. Moreover, ethanolic extract stimulates ROS level from 0.4, 8.8, 12.1 and 28.7% as well and at the dose of 250 μg mL–1 of ethanolic extract divided the indeed two population group as showed in histogram (Fig. 4a-b). CA induced HeLa cell death and apoptosis via induction ROS levels.
CA extract on cell migration: To measure the inhibitory effects of CA extract effect on cell migration, wound healing was carried out. CA extract was compared with the untreated control group, the wound healing capacity of HeLa cells decreased markedly (Fig. 5a-b, Table 2) after exposing to DW and ethanolic extract. Resulting at 48 hrs healing time, the induction of wound sizes of DW extract had significant detection at 250 μg mL–1 and ethanolic extract had more potency with significant detection at 100 and 250 μg mL–1. This data showed that both of CA extract inhibited cell migration in HeLa cervical cancer cells.
DISCUSSION
The present study indicated that edible leaf from CA noteworthy on cervical cancer cells through suppressing cell growth, stimulating cell apoptosis and inhibiting cell migration by dose-dependent manner with both DW and ethanolic extract.
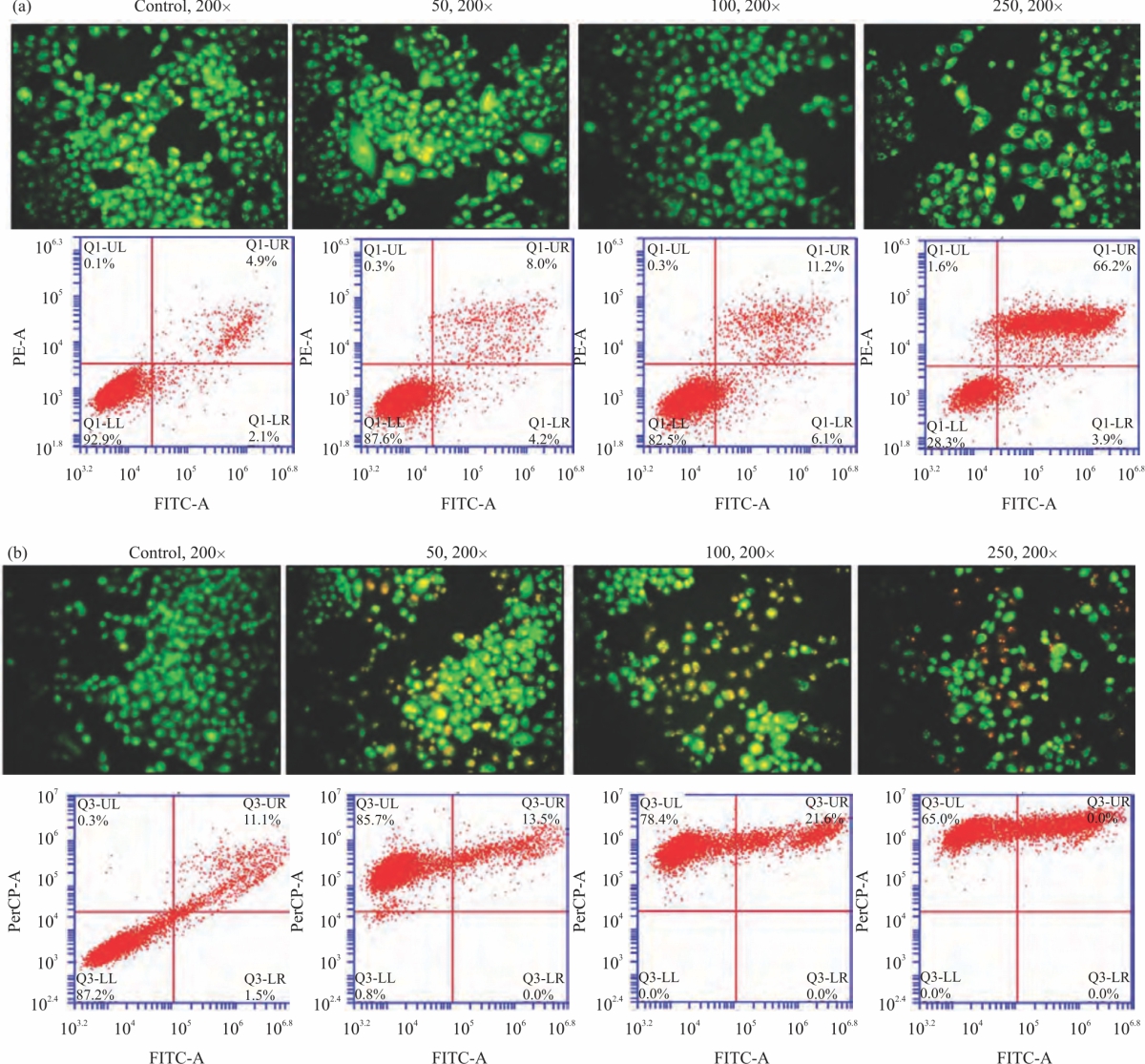 |
| Fig. 3(a-b): | CA effects on cancer cell apoptosis. (a-b) The HeLa cancer cells were incubated with DW and ethanolic CA extracts (0-250 μg mL–1) for 24 hrs, then measured morphology by fluorescent inverted microscopy (20× magnification) and apoptosis by flow cytometric analysis |
Conventional chemotherapeutic drugs are the most commonly prescribed agents used for cervical cancer treatment; however, toxic display and resistance capability related with these agents is a matter of concern. Accordingly, many new compounds are now considered for examining and developing greater anticancer agents, displaying less toxicity and needing high efficacy. The natural compounds are investigated and interested to explore the anticancer agents. CA, Thai vegetable, is mostly found in the North-East part of Thailand and has been reported against several chronic diseases including cancer.
Inhibition of cancer cell viability is one of the major hallmarks of anticancer ability of natural products or vegetables to defeat cancer cells. The SRB data indicated that CA extract suppresses the proliferation of cervical HeLa cells in a dose- and time-dependent manner, especially in ethanolic extract. Consistently, the colony formation assay presented the reduction of colony formation after treatment with CA extract by dose-dependent manner as well.
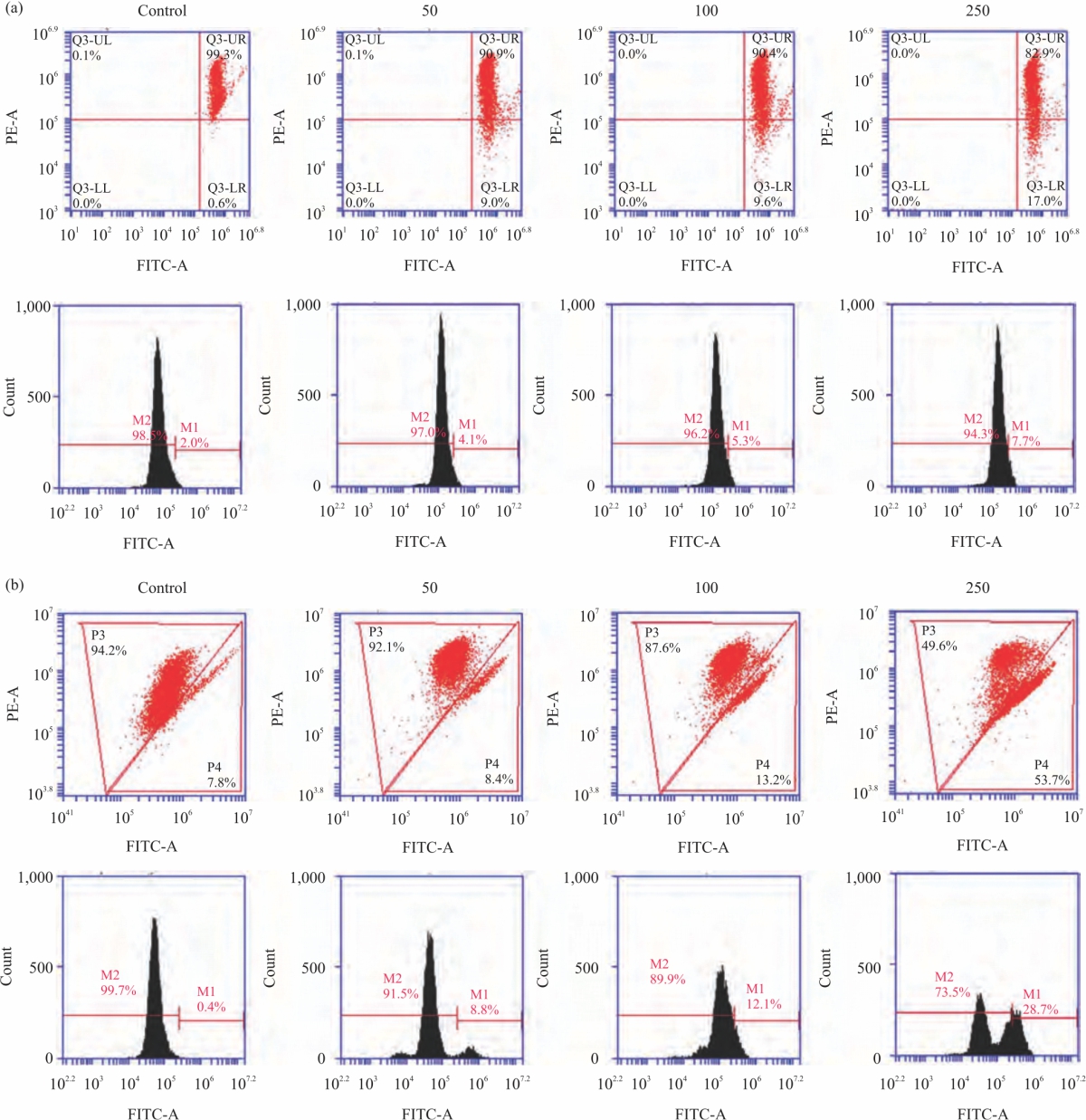 |
| Fig. 4(a-b): | CA effects on mitochondrial membrane potential (MMP) and ROS level. (a-b) The HeLa cancer cells were incubated with DW and ethanolic CA extracts (0-250 μg mL–1) for 24 hrs and then measured MMP by flow cytometric analysis. The cells were incubated with DW and ethanolic CA extracts (0-250 μg mL–1) for 24 hrs (a-b) and then measured ROS level by flow cytometric analysis Values are expressed as mean±SE. *p<0.05 was significant from control group |
Ethanolic extract had the anticancer activities more than DW extract with showing low IC50 values. Less information of CA on anticancer actions, consequently, CA has a broad spectrum on several cancer cells was very interested. Our previous study demonstrated that CA was powerfully suppressed breast cancer cells growth, IC50 values 26.28±1.11 μg mL–1 for 48 hrs, greater than cervical cancer cells, IC50 values 94.73±18.25 μg mL–1 for 48 hrs, as showing with lower IC50 values6.
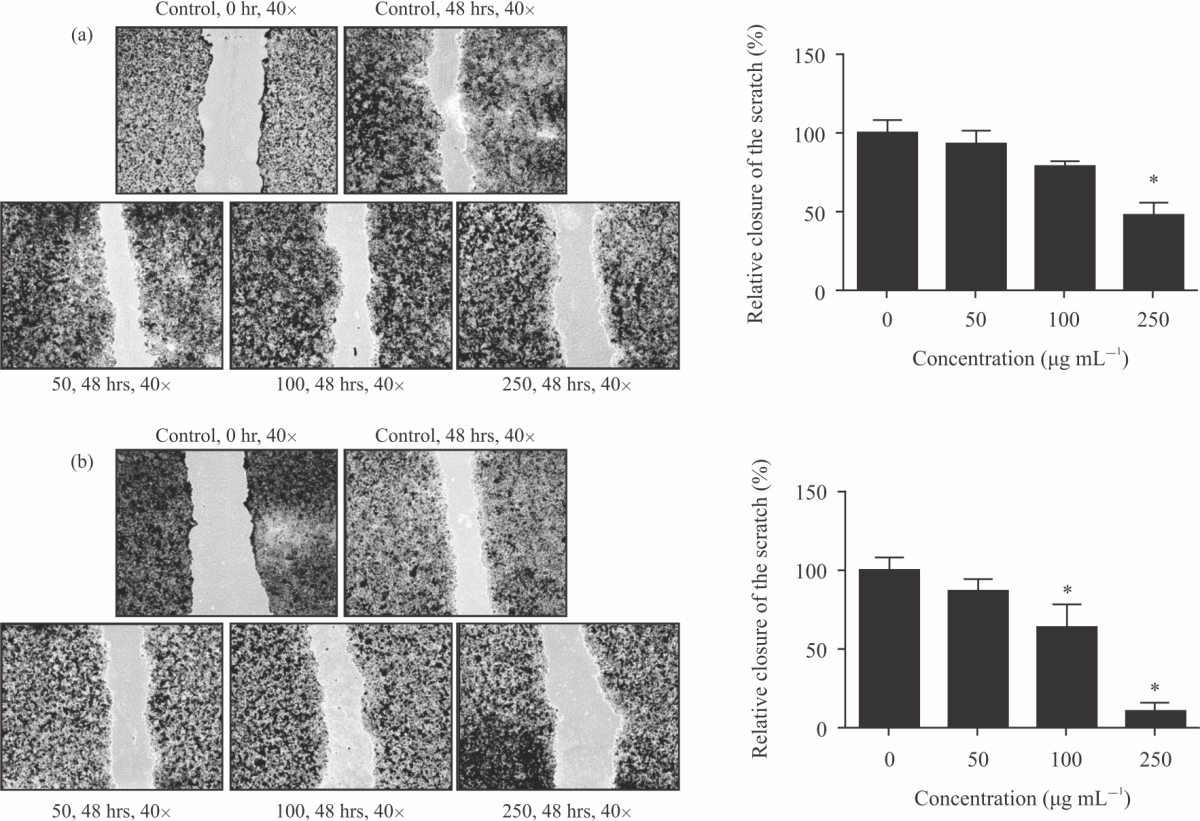 |
| Fig. 5(a-b): | CA effects on cell migration. (a-b) The HeLa cancer cells were made a wound and then incubated with DW and ethanolic CA extracts (0-250 μg mL–1) for 48 hrs. The distance of wound size was measured and calculated Values are expressed as mean±SE. *p<0.05 was significant from control group |
Shailaja et al.9, showed that ethanol extract of CA demonstrated most potent anticancer activity through its lowest GI50 (50% growth inhibition), TGI (amount for 50% Growth Inhibition) concentration on nasopharynx (KB), lung (HOP62), cervix (ME180) and leukemia (K562) cells. Moreover, our data indicated that CA caused induction of cancer cells arrest at G0/G1 phase both of two extract. The mechanism of CA action was reported that CA strongly decreased cyclin D1 level along with inducing caspase 3 and cytochrome c protein levels6. Hence, CA suppressed several cancer cells types with low concentration.
The induction of cancer cells apoptosis is a major mechanism by which many compounds/extracts display their anticancer activities10. CA extracts strongly increased caspase 3 and cytochrome c level and lead to induce breast cancer cells apoptosis6. The present study found that morphological changes in HeLa cells that were a characteristic of apoptotic fraction were detected under fluorescent inverted microscope and ethanolic extract induced more necrosis than DW extract. Following treatment with CA for 24 hrs, of apoptotic phases were carried out by Annexin V-FITC/PI double staining, the data showed that DW extract at the dose of 250 μg mL–1 strongly induces the percentage of late apoptotic cells when compared with control group. On the other hand, some of plant extract can emiss the florescent intensity by itself, therefore the data from CA ethanolic extract had some problem to detect (Fig. 3b); however, it caused in induction the late apoptotic cells by dose-dependent manner.
Interestingly, many compounds showed the action on apoptosis induction by stimulation of ROS formation and mitochondrial dysfunction11,12. ROS overproduction or reduced the intracellular antioxidant agents can result in oxidative stress13. Mitochondria is not only the important source of ROS, but also the main target of ROS14. In this study, ethanolic extract significantly increased ROS production and decreased mitochondrial function, especially at the dose of 250 μg mL–1. As ochratoxin A data indicated that this compound stimulated the ROS production and activated mitochondrial damage, shown by decreasing mitochondrial membrane potential (ΔΨM) and reducing in ATP levels as this study demonstrated12. Consequently, apoptotic pathway in mitochondrial was stimulated, represented by increase the levels of apoptotic rate and stimulation of apoptotic proteins such as cleaved form of PARP, caspase 3 and caspase 912. In conclusion, CA activated cervical cancer cells apoptosis.
Cancer/tumor is one of the most common fatal diseases in the world and death is related by metastasis from primary organ to distant organs and the development of secondary tumors15. Anticancer drugs was interested in medicinal products or plants or vegetables that suppress the migration of cancer cell for preventing cancer metastasis to distant organs16. Our previous study demonstrated that CA extract caused inhibitions of breast cancer cells migration by which significant suppressing MMP 2 and MMP 9 protein levels6. The compounds were inhibited the cancer cells migration maybe attenuated the secondary tumors occurred in many cancer types. These findings are supported by natural plants from Thailand, Kaempferia parviflora inhibited the signaling pathways of MAPK and PI3K/AKT and showed consistently with suppression of migration of HeLa cells17. On the other hand, Oroxylum indicum leaf extract inhibited human breast cancer cell migration through reducing gene expression of MMP 9 and ICAMP1 along with decreasing protein expression of MMP918. Further, Phyllanthus emblica Linn extract suppressed invasiveness of breast MDA-MB-231 cancer cells in the in vitro Matrigel invasion assay19.
These mechanisms might possibly be responsible for inhibiting migration in cancer cells and cervical cancer cells are remaining to be determined in later studies.
CONCLUSION
In conclusion, DW and ethanolic extract of CA triggers the cell growth inhibition, apoptosis induction and migratory suppression in HeLa cervical cancer cells. Attenuation of mitochondrial function and induction of ROS formation may be the mechanism of CA action on HeLa cells, in part, to its effect. These results highlight the potential of CA as an effective phytochemical for treatment of cervical cancer and the other types of cancer in further.
SIGNIFICANCE STATEMENT
This study showed that CA extract significantly inhibits cervical cancer cells proliferation, induces cancer cells apoptosis and suppresses cells migration with low dose of CA extract. The mechanism of CA extract generated the ROS formation and decreased mitochondrial function in intracellular of HeLa cells. These results suggested that CA is potential therapeutic agent against cervical cancer.
ACKNOWLEDGMENT
This research project was financially supported by Mahasarakham University 2021 (MSU2021). The authors thank Dr. Adrian R. Plant and Prof. Motoyuki Sumida (Mahasarakham University) for language-editing the manuscript.
REFERENCES
- Liu, M., Z. Wang, Q. Liu, H. Zhu and N. Xu, 2018. Expression of micro-RNA-492 (MIR-492) in human cervical cancer cell lines is upregulated by transfection with wild-type P53, irradiation, and 5-fluorouracil treatment in vitro. Med. Sci. Monit., 24: 7750-7758.
CrossRefDirect Link - Bayu, H., Y. Berhe, A. Mulat and A. Alemu, 2016. Cervical cancer screening service uptake and associated factors among age eligible women in mekelle zone, northern Ethiopia, 2015: A community based study using health belief model. PLoS ONE, Vol. 11.
CrossRefDirect Link - Song, T.T., F. Xu and W. Wang, 2020. Inhibiting ubiquitin conjugating enzyme E2 N by microRNA-590-3p reduced cell growth of cervical carcinoma. Kaohsiung J. Med. Sci., 36: 501-507.
CrossRefDirect Link - Wang, T., J. Feng and A. Zhang, 2020. miR-584 inhibits cell proliferation, migration and invasion in vitro and enhances the sensitivity to cisplatin in human cervical cancer by negatively targeting GLI1. Exp. Ther. Med., 19: 2059-2066.
CrossRefDirect Link - Dong, F. and R. Jiang, 2011. Research progress of the natural products against prostate cancer. Chin. J. Nat. Med., 9: 81-89.
Direct Link - Buranrat, B., S. Boontha, P. Temkitthawon and P. Chomchalao, 2020. Anticancer activities of Careya arborea Roxb on MCF-7 human breast cancer cells. Biologia, 75: 2359-2366.
CrossRefDirect Link - Begum, R., M. Sharma, K.K. Pillai, V. Aeri and M.A. Sheliya, 2015. Inhibitory effect of Careya arborea on inflammatory biomarkers in carrageenan-induced inflammation. Pharm. Biol., 53: 437-445.
CrossRefDirect Link - Natesan, S., S. Badami, S.H. Dongre and A. Godavarthi, 2007. Antitumor activity and antioxidant status of the methanol extract of Careya arborea bark against dalton`s lymphoma ascites-induced ascitic and solid tumor in mice. J. Pharmacol. Sci., 103: 12-23.
CrossRefDirect Link - Shailaja, W., W. Balaji and K. Mahesh, 2019. Identification of bioactive compounds and cytotoxic activity of Careya arborea Roxb. leaves. J. Pharmacogn. Phytochem., 8: 362-365.
Direct Link - Madunić, I.V., J. Madunić, M. Antunović, M. Paradžik and V. Garaj-Vrhovac et al., 2018. Apigenin, a dietary flavonoid, induces apoptosis, DNA damage, and oxidative stress in human breast cancer MCF-7 and MDA MB-231 cells. Naunyn-Schmiedeberg`s Arch. Pharmacol., 391: 537-550.
CrossRefDirect Link - Klungsaeng, S., V. Kukongviriyapan, A. Prawan, S. Kongpetch and L. Senggunprai, 2019. Cucurbitacin B induces mitochondrial-mediated apoptosis pathway in cholangiocarcinoma cells via suppressing focal adhesion kinase signaling. Naunyn-Schmiedeberg's Arch. Pharmacol., 392: 271-278.
CrossRefDirect Link - Li, Q., Z. Dong, W. Lian, J. Cui and J. Wang et al., 2019. Ochratoxin A causes mitochondrial dysfunction, apoptotic and autophagic cell death and also induces mitochondrial biogenesis in human gastric epithelium cells. Arch. Toxicol., 93: 1141-1155.
CrossRefDirect Link - Bhat, P.V., M. Pandareesh, F. Khanum and A. Tamatam, 2016. Cytotoxic effects of ochratoxin A in neuro-2a cells: Role of oxidative stress evidenced by N-acetylcysteine. Front. Microbiol., Vol. 7.
CrossRefDirect Link - Wang, H., Y. Chen, N. Zhai, X. Chen, F. Gan, H. Li and K. Huang, 2017. Ochratoxin A-induced apoptosis of IPEC-J2 cells through ROS-mediated mitochondrial permeability transition pore opening pathway. J. Agric. Food Chem., 65: 10630-10637.
CrossRefDirect Link - Seyfried, T.N. and L.C. Huysentruyt, 2013. On the origin of cancer metastasis. Crit. Rev. Oncogenesis, 18: 43-73.
Direct Link - Steeg, P.S. and D. Theodorescu, 2008. Metastasis: A therapeutic target for cancer. Nat. Clin. Pract. Oncol., 5: 206-219.
CrossRefDirect Link - Potikanond, S., S. Sookkhee, M.N. Takuathung, P. Mungkornasawakul, N. Wikan, D.R. Smith and W. Nimlamool, 2017. Kaempferia parviflora extract exhibits anti-cancer activity against HeLa cervical cancer cells. Front. Pharmacol., Vol. 8.
CrossRefDirect Link - Buranrat, B., S. Noiwetch, T. Suksar and A. Ta-Ut, 2020. Inhibition of cell proliferation and migration by Oroxylum indicum extracts on breast cancer cells via Rac1 modulation. J. Pharm. Anal., 10: 187-193.
CrossRefDirect Link - Ngamkitidechakul, C., K. Jaijoy, P. Hansakul, N. Soonthornchareonnon and S. Sireeratawong, 2010. Antitumour effects of Phyllanthus emblica L.: Induction of cancer cell apoptosis and inhibition of in vivo tumour promotion and in vitro invasion of human cancer cells. Phytother. Res., 24: 1405-1413.
CrossRefDirect Link