
Ismaeel Bin-Jaliah
Department of Physiology, College of Medicine, King Khalid University, P.O. Box 641, Abha, 61421, Aseer, Saudi Arabia
LiveDNA: 966.36118
International Journal of Pharmacology
Year: 2021 | Volume: 17 | Issue: 5 | Page No.: 319-327







ABSTRACT
Background and Objective: Exposure to chronic stress is harmful to vital organs such as the brain and the heart. The potential inhibitory effects of the polyphenolic compound quercetin on Chronic Unpredictable Stress (CUS)-induced depression and biomarkers of brain injury associated with the inhibition of Nitrosative Stress (iNOS) and the apoptotic axis p53-Bax-caspase-3 has not been investigated before. Materials and Methods: Rats were either exposed to a variety of unpredictable stressors daily before being sacrificed after 3 weeks or were treated for 3 weeks with quercetin (50 mg kg–1 b.wt./day). Animals were then culled and brain tissues were harvested. Results: CUS significantly (p<0.05) induced iNOS, MDA, p53, Bax and caspase-3, which were significantly inhibited by quercetin. Whereas, quercetin significantly increased brain tissue levels of SOD and Bcl-2. In addition, CUS caused a significant increase in animal immobility and a decrease in climbing ability and sucrose consumption, which were reverted by quercetin. Furthermore, a significant (p<0.0001) correlation between either cerebral cortex brain injury and biomarkers of apoptosis and survival, p53 and Bcl-2, or between p53 and Bcl-2 and biomarkers of nitrosative stress and depression were observed. Conclusion: Quercetin protects against CUS-induced cerebral cortex injury and depression, which is associated with the inhibition of the p53-Bax-caspase-3 axis and biomarkers of nitrosative and oxidative stress.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ismaeel Bin-Jaliah, 2021. Quercetin Inhibits Chronic Stress-Induced Depression Associated with the Inhibition of Nitrosative Stress and Apoptosis. International Journal of Pharmacology, 17: 319-327.
DOI: 10.3923/ijp.2021.319.327
URL: https://scialert.net/abstract/?doi=ijp.2021.319.327
DOI: 10.3923/ijp.2021.319.327
URL: https://scialert.net/abstract/?doi=ijp.2021.319.327
INTRODUCTION
Repeated stress is a risk factor for depression, general anxiety disorder and neurological abnormalities in both humans and experimental animal models1,2 and about 10% of the US population is inflicted with depression3. In addition, exposure to stressors for a prolonged period can affect other systems such as the endocrine and cardiovascular systems4,5. Indeed, chronic stress caused cardiac arrest and death in more than a quarter of the mice that had their anti-stress receptor, serotonin knocked out6. Also, mental stress, depression and anxiety are recognised amongst many risk factors that triggermyocardialinfarction7.
To cope with stress, the body mobilizes physiological and psychological resources leading to alterations in the dynamic regulations of the autonomic, neuroendocrine and immune systems8. Accumulated evidence shows that chronic stress such as those arising from natural disasters, wars, economic deprivation or induced by experimental procedures, is a risk factor for the central and peripheral nervous systems9,10. For example, (i) exposure to various forms of psychological or occupational stressors resulted in the stimulation of the hypothalamic-pituitary-adrenal axis which leads to the increase in the secretion of the stress hormone cortisol9 and (ii) oxidative damage to neurons has been implicated in the pathogenesis of depression11. Furthermore, tissue oxidative stress is believed to be the main cause behind damage that occurs in animals and humans exposed to traumatic events including chronic stress12. Stress-induced overproduction of Reactive Oxygen Species (ROS) caused apoptosis and enhanced levels of lipid peroxidation and peroxynitrite that damages DNA in the brain13. In addition, the role of lowered levels of antioxidants and high generation of ROS and nitrosative stress in the pathogenesis of depression have been reported, which have pointed to neuroinflammation, increased apoptosis rate and altered neurogenesis/ neuroplasticity14.
Quercetin is a flavonol antioxidant compound that is found in many vegetables, fruits and grains15,16. Quercetin has many pleiotropic effects that demonstrated effective protection to the cardiovascular, kidney and liver17-19. Quercetin was also reported to protect against diabetes-induced depression in mice20. However, little is known about the protective effects of quercetin against chronic stress-induced brain injury.
Therefore, this study aimed to investigate the effects of quercetin on CUS-induced depression in rats and to monitor brain tissue levels of the p53-Bax-caspase-3 axis of apoptosis, the survival cell signalling and nitrosative stress in the presence and absence of quercetin.
MATERIALS AND METHODS
Study area: The study was carried out at the Research Center, College of Medicine, King Khalid University, Abha, Saudi Arabia from September-December, 2019.
Animals: All rat studies were overseen and approved by the Medical Research Ethical Committee at King Khalid University, Abha, Saudi Arabia. Ref no: (Rec. No. 2014-06-09). Male Wistar rats (total 24 rats) at 8 weeks and weighing 150-200 g were used for these studies. They had free access to food and water and were housed under a constant temperature of 23±1°C with a 12 hrs light/dark cycle.
Experimental design: After a one-week adaptation period, rats were randomly assigned into 4 groups (n = 6 each) as follows: 1. Control group: Received normal saline. 2. Quercetin treated group (QUR): Rats received QUR (50 mg kg–1)20. 3. Chronic unpredictable stress (CUS) group: A model group and were exposed to CUS protocol, as detailed below and received normal saline. 4. CUS+QUR treated group: were exposed to CUS with a concomitant daily dose of QUR (50 mg kg–1). All treatments were administered as 1 mL, i.p. for three consecutive weeks daily.
Chronic unpredictable stress (CUS) protocol: CUS protocol was induced as previously described21 with modification. Briefly, a set of chronic unpredictable stressors were used to induce brain injury in rats that lasted for 3 weeks.
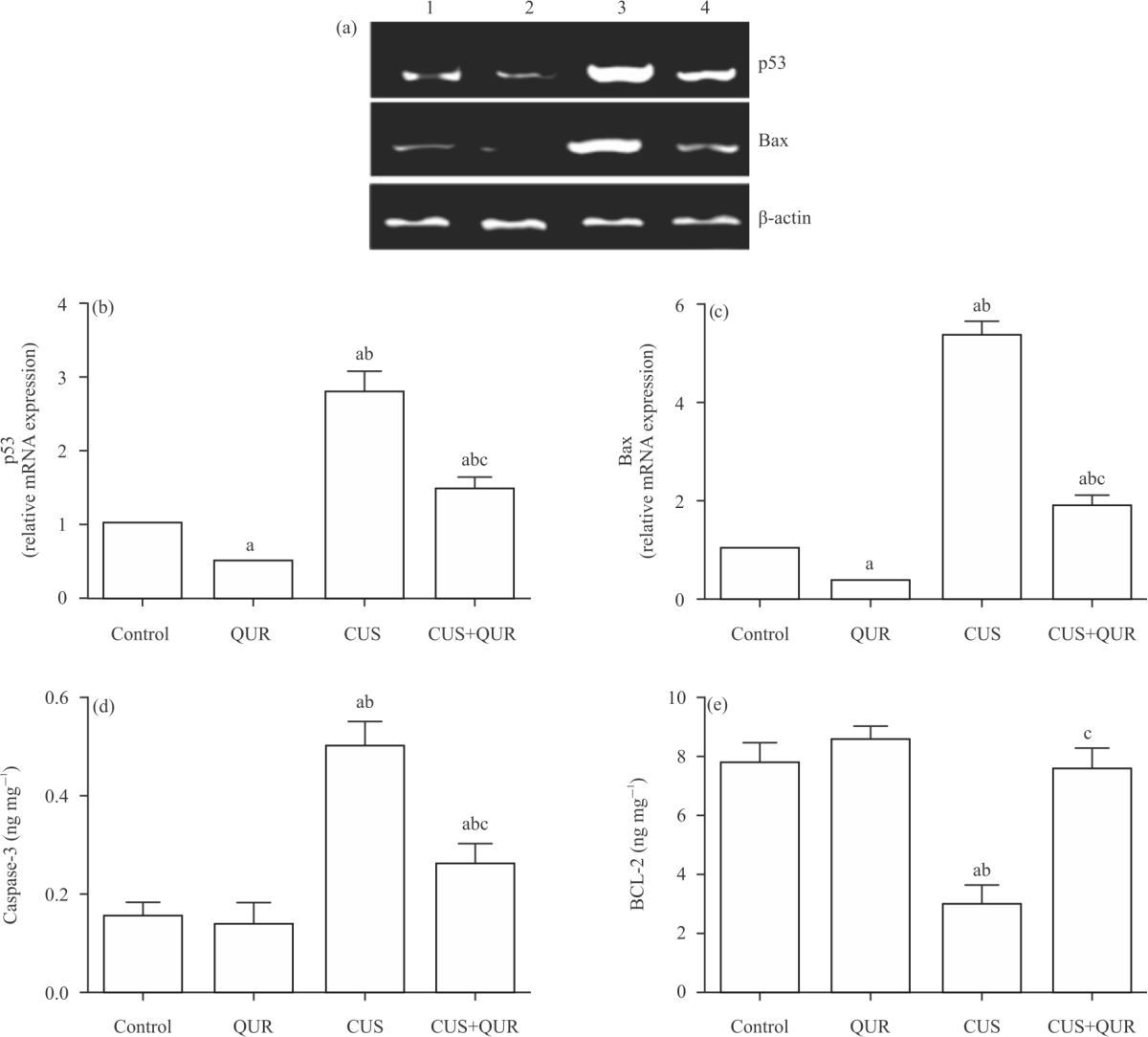
Assessment of depressive-like behaviours; forced swimming and sucrose preference tests: The experiment of assessing depressive-like behaviour in the form of a forced swimming test was carried out as previously described22 to induce brain injury in rats for three weeks. All scoring was done by a single trained person, blind to experimental conditions. The sucrose preference test procedure was performed as previously described23. The sucrose preference was calculated using the following formula:
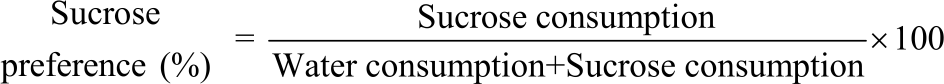 |
Preparation of brain tissues for analysis: At the end of the experimental period, blood samples were collected by cardiac puncture under anaesthesia (sodium thiopental at 40 mg kg–1 b.wt.) and rats were then culled using cervical dislocation. Brain tissues were harvested and washed with iced Phosphate-Buffered Saline (PBS). The brains were cut into longitudinal sections. Parts of the brains were homogenized in a cold phosphate buffer, pH 7.4, containing EDTA. The supernatant obtained was distributed in separate tubes and stored at -70°C for biochemical analysis. Other parts of the brains were stored at -70°C for RNA extraction.
Detection of p53 and Bax messenger RNAs by reverse transcriptase-polymerase chain reaction: As previously described24, total RNAs were isolated from freshly dissected rats’ brains using the RNeasy Mini Kit (Qiagen Pty, Victoria, Australia). The RNA was reverse-transcribed for a single strand cDNA synthesis (Invitrogen) and amplified by PCR for the tumour suppressor p53, apoptosis regulator Bax and β-actin. The PCR products were separated by 2% agarose gel electrophoresis and visualised by ethidium bromide. Gel images were scanned and quantified by densitometry using the NIH image software.
Determination of tissue levels of nitrosative and oxidative stress and apoptosis and survival biomarkers: ELISA kits were used according to the manufacturers’ instructions to determine the tissue levels of malondialdehyde (MDA) for lipid peroxidation (Cat No. NWK-MDA01, NWLSS, USA), Superoxide Dismutase (SOD) (Cat. No. 706002, Cayman Chemical, Ann Arbor, MI, USA),Glutathione Peroxidase (GPx) (Cat. No. 703102, Cayman Chemical, Ann Arbor, MI, USA), caspase 3 (Cat. No. R5814), B-cell lymphoma 2 (Bcl-2) (Cat. No. R6813) and inducible Nitric Oxide Synthase (iNOS) (Cat. No R6663) were purchased from STZ ELISA (USA).
Statistical and morphometric analysis: Statistical analysis was performed by using the Graph pad Prism statistical software package (version 6). The data were expressed as mean±standard deviation (SD).One-way analysis of variance (ANOVA) was performed followed by Tukey’s t-test. Using the "Leica Qwin 500 C" image analyzer (Cambridge, UK), the degree of cerebral cortex damage was obtained in 10 non-overlapping high-power fields/ rat of H and E-stained sections. Quantitative data were tabulated as a means and Standard Deviations (SD) and compared using analysis of variance (ANOVA) followed by post hoc analysis (Tukey test). A significant difference was considered when p-value 0.05. Calculations were made on SPSS software (version 19).
RESULTS
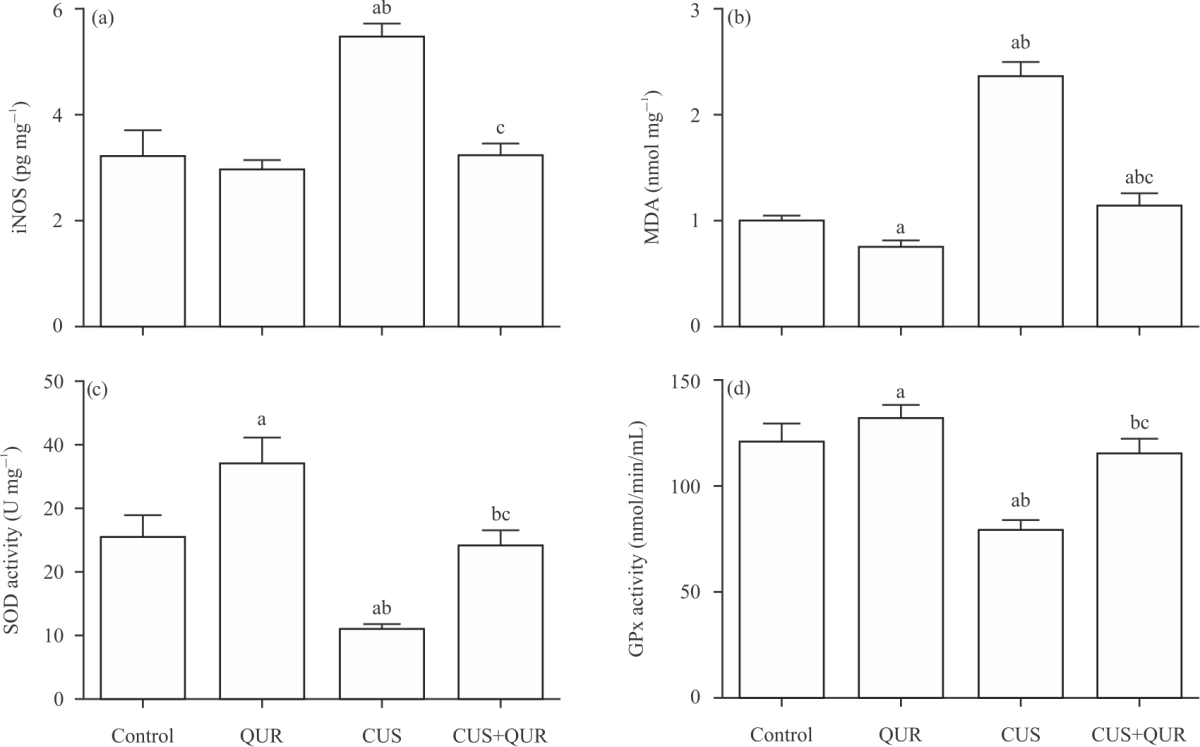
Quercetin inhibits CUS-induced brain injury biomarkers and depression: To test the hypothesis that quercetin in this modified model of CUS can protect against brain injury and depression caused by CUS, we measured iNOS, MDA as a by-product of lipid peroxidation and the antioxidants SOD and Gpx using the ELISA method in brain tissue homogenates of all groups of rats. Compared with the control untreated group, CUS significantly (p<0.0001) augmented iNOS (Fig. 1a) and MDA (Fig. 1b) and ameliorated SOD (Fig. 1c) and Gpx (Fig. 1d) tissue levels, which were significantly (p<0.0001; CUS vs CUS+QUR) protected by quercetin. However, the levels of MDA in the treated group (CUS+QUR) were still significant (p<0.046) to the two control groups (Fig. 1b) which mean, partial inhibition of MDA was achieved by quercetin. We further assessed depression-like behaviour levels in these animals in the presence and absence of quercetin. As shown in Fig. 2, CUS significantly (p<0.0001) inhibited mobility (Fig. 2a; increased immobility) and climbing ability (Fig. 2b) that were significantly (p<0.0001; CUS vs. CUS+QUR) protected by quercetin. However, exposing rats to CUS for 3 weeks did not affect their swimming ability (Fig. 2c). Sucrose consumption (Fig. 2d) was inhibited by CUS and protected with quercetin. The degree of protection exerted by quercetin was partial in (A).
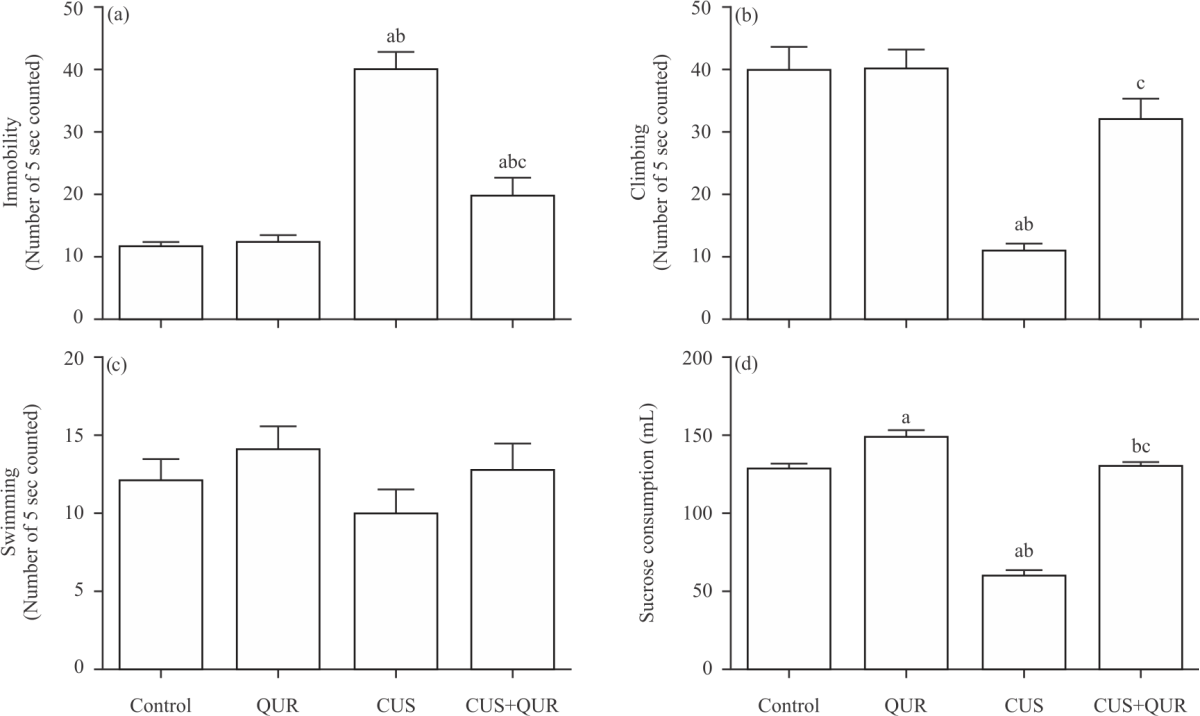
Quercetin protects CUS-modulated apoptosis and survival biomarkers in brain tissues: We then tested the apoptotic p53-Bax-caspase-3 axis cell signalling in brain tissue CUS strongly (i) augmented p53 and Bax cDNA message (Fig. 3a-c); (ii) augmented caspase-3 protein expression (Fig. 3d) and (iii) inhibited the survival protein Bcl-2 (Fig. 3e), which were effectively (p<0.0001; CUS vs. CUS+QUR) protected by quercetin. However, complete protection was only seen in (E).
Correlation between p53 or Bcl-2 and biomarkers of depression and nitrosative and oxidative stress: The correlation between either p53 or Bcl-2 scorning and tissue levels of nitrosative and oxidative stress and depression biomarkers were determined to further support the link between p53 and Bcl-2 and brain injury and to further confirm and characterize the role of QUR as being stable and an appropriate agent in brain injury rats. A significant (p<0.0001) correlation was observed between p53 and iNOS (r = 0.849) (Fig. 4a), p53 and MDA (r = 0.918) (Fig. 4b), p53 and climbing ability (r = -0.898) (Fig. 4c), p53 and sucrose consumption (r = - 0.915) (Fig. 4d), Bcl-2 and iNOS (r = -0.936) (Fig. 4e), Bcl-2 and MDA (r = - 0.946) (Fig. 4f), Bcl-2 and climbing ability (r = 0.921) (Fig. 4g) and Bcl-2 and sucrose consumption (r = 0.955) (Fig. 4h).
 |
| Fig. 1(a-d): | Quercetin protects the modulation of nitrosative and oxidative stress and anti-oxidants biomarkers caused by CUS Brain tissue levels of (a) iNOS, (b) MDA, (c) SOD and (d) GPx were measured at the end of the experiment in all groups of rats; control, QUR, CUS and CUS+QUR groups. Results represent the mean (±SD); n = 6 for each group. All shown p-values are significant; ap<0.05 versus control, b p<0.05 versus QUR, cp<0.05 versus CUS. CUS: Chronic uncontrolled stress, QUR: Quercetin, iNOS: Inducible nitric oxide synthase, MDA: Malondialdehyde, SOD: Superoxide dismutase, GPx: Glutathione peroxidase |
 |
| Fig. 2(a-d): | Quercetin inhibits CUS-induced depression-like behaviour levels in rats Markers of depression, (a) Immobility, (b) Climbing ability, (c) Swimming ability and (d) Sucrose consumption were assessed at the end of the experiment in the animal groups used in this study; Control, QUR, CUS and CUS+QUR groups. Results represent the mean (±SD), n = 6 for each group. ap<0.05 versus control,p<0.05 versus QUR, cp<0.05 versus CUS. CUS: Chronic uncontrolled stress, QUR: Quercetin |
 |
| Fig. 3(a-e): | Quercetin protects the modulation of apoptotic and survival biomarkers caused by CUS (a-c) Brain tissue levels of p53 and Bax gene expression were measured using RT-PCR analysis, (d) Caspase-3 and (e) Bcl-2 protein expression measured by ELISA at the end of the experiment in all groups of rats; control, QUR, CUS and CUS+QUR groups. For (A): Lane 1: Control, Lane 2: QUR, Lane 3: CUS and Lane 4: CUS+QUR groups. Results represent the mean (±SD); n = 6 for each group. ap<0.05 versus control, bp<0.05 versus QUR, cp<0.05 versus CUS. CUS: Chronic uncontrolled stress, QUR: Quercetin, p53: Tumour suppressor p53, Bax: Apoptosis regulator Bax, Bcl-2: B-cell lymphoma 2 |
Quercetin partially protects brain tissue against CUS-induced injury: Given the results described above that showed substantial protection by quercetin to CUS-induced depression, apoptosis and nitrosative and oxidative stress, we assessed the degree of potential protection to brain tissue injury provided by quercetin in response to CUS. Compared to normal tissue structure in the control group (Fig. 5a), CUS caused profound tissue damage as demonstrated by substantial neuronal (N) damage with shrunken cells, disrupted atrophic nuclei (n), perineuronal vacuolation (arrows) and swollen glial cells (G) are seen (Fig. 5b). Whereas, tissue prepared from the cerebral cortex of the brain of the treatment group (CUS+QUR) showed partially preserved brain architecture (Fig. 5c-d), as shown by mixed populations of cells; intact and damaged.
We further determined the correlation between the score of the cerebral cortex brain injury and the tissue levels of p53 and Bcl-2 to support the link between brain injury and apoptosis. As displayed in Fig. 5(e-f), a positive correlation was scored between brain tissue injury and p53 (r = 0.919; p<0.0001) (Fig. 5e) and a negative correlation between brain tissue injury and Bcl-2 (r = -0.879; p<0.0001) (Fig. 5f).
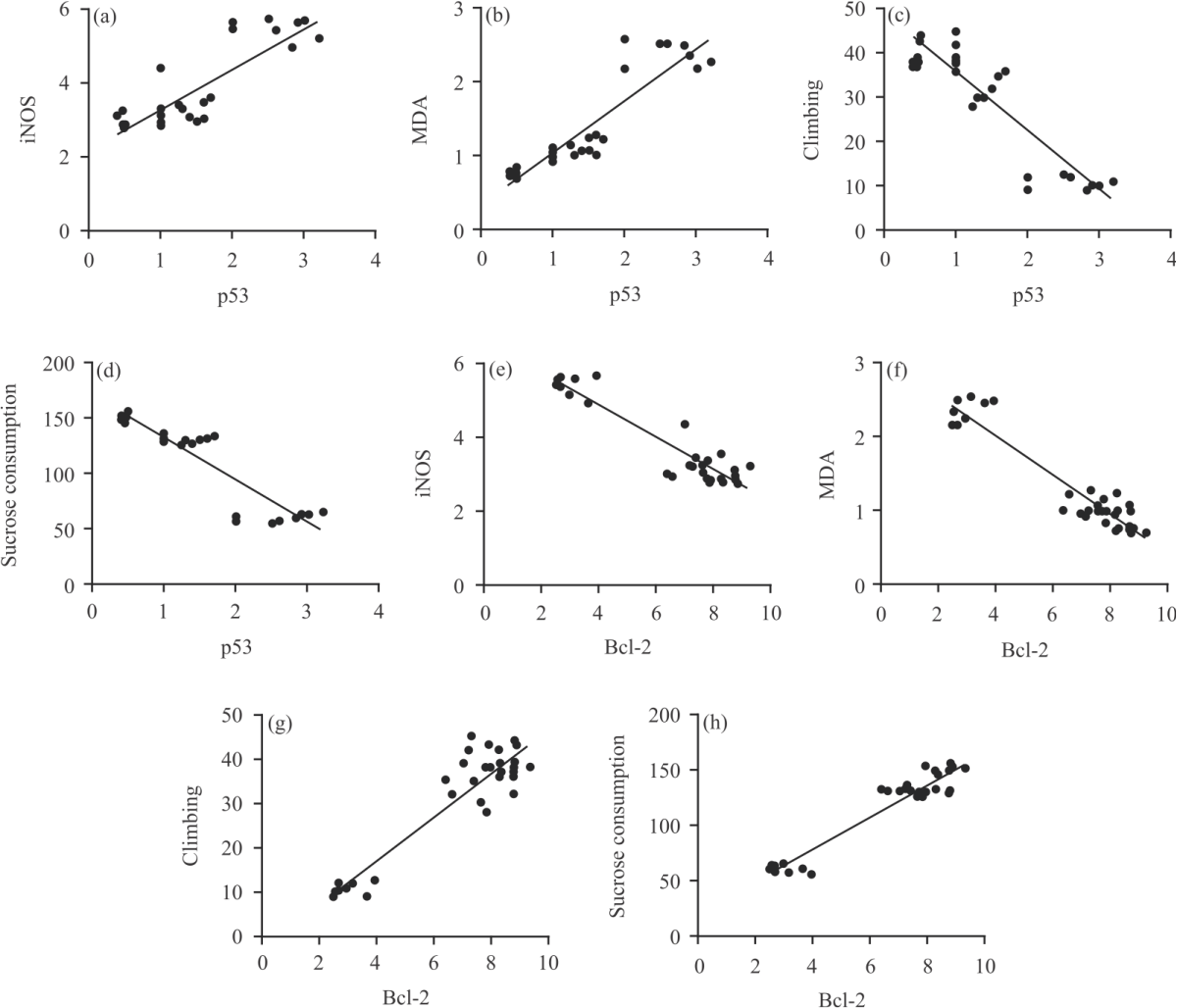 |
| Fig. 4(a-h): | Correlation between p53 or Bcl-2 and markers of depression and iNOS and MDA (a-d) P53 and Bcl-2 brain homogenate levels were measured in all groups of rats and the correlation between p53 and iNOS, p53 and MDA, p53 and climbing ability and p53 and sucrose consumption, respectively and (e-h) Correlation between Bcl-2 and iNOS, MDA, Bcl-2 and climbing ability and Bcl-2 and sucrose consumption, respectively |
DISCUSSION
One of the main findings of our study was that exposure of rats to CUS for 3 weeks led to the activation of a p53-Bax-caspase-3 axis of apoptosis in brain tissue, which was inhibited by quercetin. We were required to generate this animal model of CUS-induced depression to investigate the apoptotic pathways induced by CUS in the presence and absence of quercetin. Also, our protective regimen of quercetin was used to assess brain tissue levels of the survival (anti-apoptotic) protein Bcl-2 and biomarkers of nitrosative and oxidative stress, which demonstrated beneficial effects of quercetin. Furthermore, our data show (i) a positive correlation between p53 score and the brain tissue levels of iNOS and MDA; (ii) a negative correlation between p53 score and climbing ability and sucrose consumption and (iii) a significant correlation between cerebral cortex injury score and p53 and Bcl-2 support our conclusion mentioned above. Our results were thus consistent with our working hypothesis that CUS can substantially induce the activity of brain tissue p53-Bax-caspase-3 axis, which is an apoptotic pathway located downstream of the oxidative and nitrosative stress in the cell signalling pathways25,26. This axis was blocked by quercetin in a rat model of CUS-induced depression. Therefore, the findings of this study confirm those previously published that showed both types of stress, acute and chronic, induced several pathological manifestations of brain injuries in animal models27,28, such as behaviour and oxidative status of restraint-stressed mice29, psychological stress-induced enhancement of brain lipid peroxidation via nitric oxide30 and quercetin pre-treatment protects against stress-induced anxiety and depression-like behaviour and improves memory in male mice31.
In cell signalling, nitrosative and oxidative stress are located upstream of the apoptotic biomarkers upon activation in many cases such as brain and vascular injury25,32,33.
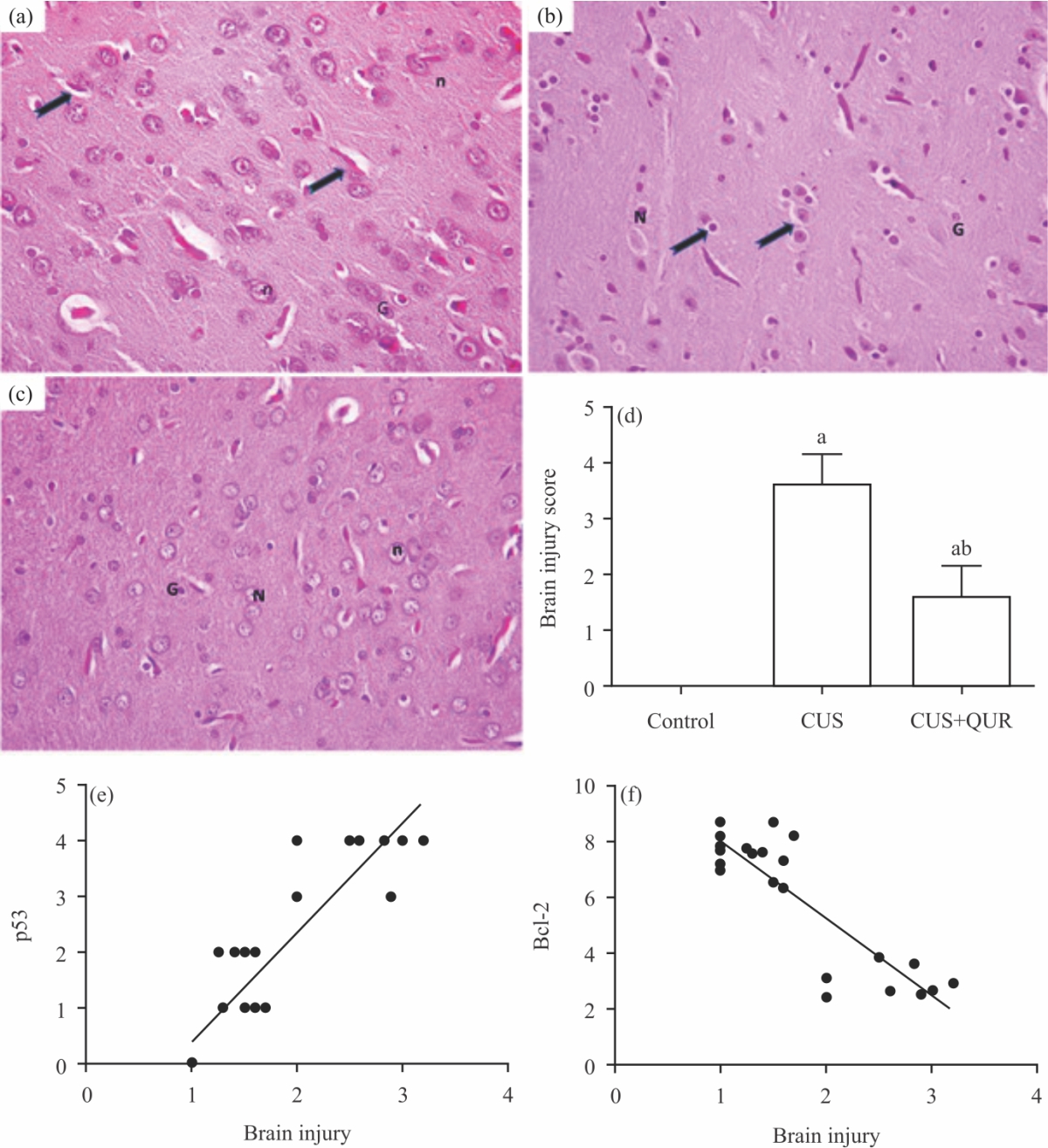 |
| Fig. 5(a-f): | Quercetin partially protects brain architecture against injury induced by CUS (a) H and E stained images (400x) of harvested rat tissues obtained after 3 weeks from the brain cerebral cortex of the control group, (b) CUS group, (c) Protective group, CUS+QUR are visualized by light microscopy. Note that in (A), arrows point to the glial processes and in (B), point to the perineuronal vacuolation. N: Neurons, n: Nucleus, G: Glial cells. Histograms in (d) represent a quantitative analysis of the cerebral cortex brain injury in the three groups of rats. All shown p values are significant, ap<0.05 versus control, b p<0.05 versus QUR and (e-f) Correlation between the brain injury score versus (e) p53 and (f) Bcl-2. CUS: Chronic uncontrolled stress, QUR: Quercetin, p53: Tumour suppressor p53, Bcl-2: B-cell lymphoma 2 |
Oxidative and nitrosative stress and p53 were reported to be involved in Alzheimer disease26. In addition, oxidative stress-activated a specific p53 transcriptional response, which regulates cellular senescence and ageing in mice and deletion of p53 and the redox that generates mitochondrial ROS by genetic means reduced apoptosis and increased longevity of transgenic mice26. Furthermore, overexpression of iNOS caused induction of vascular endothelial cell apoptosis and caspase-3 and 9activities,which were reduced by SMT, a selective iNOS inhibitor33. These reports are in agreement with our data that demonstrate the association between CUS-induced depression and apoptosis and oxidative and nitrosative stress (Fig. 1-4). Also, our data shown here are in agreement with our recently published report24 that showed modulation of these parameters; p53, Bax, Bcl-2 and biomarkers of oxidative and nitrosative stress in acute liver injury. Finally, in vivo study of quercetin on humans is warranted to evaluate whether these animal findings can be translated into therapy. Also, a future study would use quercetin to augment the effects of an antidepressant drug in this model.
CONCLUSION
In summary, we have demonstrated that induction of CUS-induced depression in rats for three weeks causes the activation of brain tissue p53-Bax-caspase-3 axis of apoptosis associated with the induction of oxidative and nitrosative stress and inhibition of the survival protein Bcl-2 and quercetin can revert these parameters.
SIGNIFICANCE STATEMENT
This study represents a significant contribution in the study of brain injury and depression induced by chronic stress in rats since it investigates the brain tissue apoptotic and survival cell signalling molecules modulated by chronic stress and the potential protection of these molecules and inhibition of brain injury and depression by the antioxidant compound quercetin, which may offer therapeutic potential in humans.
ACKNOWLEDGMENTS
This work was supported by the Research Deanship of King Khalid University, Abha, Saudi Arabia; Grant number No. R.G.P.1/44/42. The author highly appreciates the support of Professors Bahjat Al-Ani, Mohamed A Haidara and Mohamed Samir Zaki from the College of Medicine, King Khalid University, Abha, Saudi Arabia. Thanks are due to Dr. Mariam Al-Ani for proofreading the manuscript.
REFERENCES
- Bondi, C.O., G. Rodriguez, G.G. Gould, A. Frazer and D.A. Morilak, 2008. Chronic unpredictable stress induces a cognitive deficit and anxiety-like behavior in rats that is prevented by chronic antidepressant drug treatment. Neuropsychopharmacology, 33: 320-331.
CrossRefDirect Link - Opel, N., R. Redlich, P. Zwanzger, D. Grotegerd and V. Arolt et al., 2014. Hippocampal atrophy in major depression: A function of childhood maltreatment rather than diagnosis? Neuropsychopharmacology, 39: 2723-2731.
CrossRefDirect Link - Kessler, R.C., P. Berglund, O. Demler, R. Jin and D. Koretz et al., 2003. The epidemiology of major depressive disorder: Results from the national comorbidity survey replication (NCS-R). JAMA, 289: 3095-3105.
CrossRefPubMedDirect Link - Helmreich, D.L., D.B. Parfitt, X.Y. Lu, H. Akil and S.J. Watson, 2005. Relation between the Hypothalamic-Pituitary-Thyroid (HPT) axis and the Hypothalamic-Pituitary-Adrenal (HPA) axis during repeated stress. Neuroendocrinology, 81: 183-192.
CrossRefDirect Link - O’Donnell, K., L. Brydon, C.E. Wright and A. Steptoe, 2008. Self-esteem levels and cardiovascular and inflammatory responses to acute stress. Brain, Behav. Immun., 22: 1241-1247.
CrossRefDirect Link - Carnevali, L., F. Mastorci, E. Audero, G. Graiani and S. Rossi et al., 2012. Stress-induced susceptibility to sudden cardiac death in mice with altered serotonin homeostasis. PLoS ONE, Vol. 7.
CrossRefDirect Link - Dimsdale, J.E., 2008. Psychological stress and cardiovascular disease. J. Am. Coll. Cardiol., 51: 1237-1246.
CrossRefDirect Link - Palego, L., G. Giannaccini and L. Betti, 2021. Neuroendocrine response to psychosocial stressors, inflammation mediators and brain-periphery pathways of adaptation. Cent. Nervous Syst. Agents Med. Chem., 21: 2-19.
CrossRefDirect Link - Alkadhi, K., M. Zagaar, I. Alhaider, S. Salim and A. Aleisa, 2013. Neurobiological consequences of sleep deprivation. Curr. Neuropharmacol., 11: 231-249.
CrossRefDirect Link - Hovatta, I., J. Juhila and J. Donner, 2010. Oxidative stress in anxiety and comorbid disorders. Neurosci. Res., 68: 261-275.
CrossRefPubMedDirect Link - Nunomura, A., T. Tamaoki and N. Motohashi, 2014. Role of oxidative stress in the pathophysiology of neuropsychiatric disorders. Seishin Shinkeigaku Zasshi, 116: 842-858.
Direct Link - Chew, S.H. and S. Toyokuni, 2015. Malignant mesothelioma as an oxidative stress-induced cancer: An update. Free Radical Biol. Med., 86: 166-178.
CrossRefDirect Link - Kalagatur, N.K., E.F. Abd_Allah, S. Poda, K. Kadirvelu, A. Hashem, V. Mudili and C. Siddaiah, 2021. Quercetin mitigates the deoxynivalenol mycotoxin induced apoptosis in SH-SY5Y cells by modulating the oxidative stress mediators. Saudi J. Biol. Sci., 28: 465-477.
CrossRefDirect Link - Maes, M., P. Galecki, Y.S. Chang and M. Berk, 2011. A review on the Oxidative and Nitrosative Stress (O&NS) pathways in major depression and their possible contribution to the (neuro) degenerative processes in that illness. Progr. Neuro-Psychopharmacol. Biol. Psychiatry, 35: 676-692.
CrossRefPubMedDirect Link - Nishimuro, H., H. Ohnishi, M. Sato, M. Ohnishi-Kameyama and I. Matsunaga et al., 2015. Estimated daily intake and seasonal food sources of quercetin in japan. Nutrients, 7: 2345-2358.
CrossRefDirect Link - Xu, D., M.J. Hu, Y.Q. Wang and Y.L. Cui, 2019. Antioxidant activities of quercetin and its complexes for medicinal application. Molecules, Vol. 24.
CrossRefDirect Link - Chen, Y.W., H.C. Chou, S.T. Lin, Y.H. Chen, Y.J. Chang, L. Chen and H.L. Chan, 2013. Cardioprotective effects of quercetin in cardiomyocyte under ischemia/reperfusion injury. Evid. Complement. Altern. Med., Vol. 2013.
CrossRefDirect Link - Zhang, J., Y. Sheng, L. Shi, Z. Zheng, M. Chen, B. Lu and L. Ji, 2017. Quercetin and baicalein suppress monocrotaline-induced hepatic sinusoidal obstruction syndrome in rats. Eur. J. Pharmacol., 795: 160-168.
CrossRefDirect Link - Yang, H., Y. Song, Y.N. Liang and R. Li, 2018. Quercetin treatment improves renal function and protects the kidney in a rat model of adenine-induced chronic kidney disease. Med. Sci. Monit., 24: 4760-4766.
CrossRefDirect Link - Anjaneyulu, M., K. Chopra and I. Kaur, 2003. Antidepressant activity of quercetin, a bioflavonoid, in streptozotocin-induced diabetic mice. J. Med. Food, 6: 391-395.
CrossRefDirect Link - Harro, J., R. Häidkind, M. Harro, A.R. Modiri and P.G. Gillberg et al., 1999. Chronic mild unpredictable stress after noradrenergic denervation: Attenuation of behavioural and biochemical effects of DSP-4 treatment. Eur. Neuropsychopharmacol., 10: 5-16.
CrossRefDirect Link - Yankelevitch-Yahav, R., M. Franko, A. Huly and R. Doron, 2015. The forced swim test as a model of depressive-like behavior. J. Visualized Exp., Vol. 2.
CrossRefDirect Link - Grippo, A.J., N.R. Sullivan, K.J. Damjanoska, J.W. Crane and G.A. Carrasco et al., 2005. Chronic mild stress induces behavioral and physiological changes, and may alter serotonin 1A receptor function, in male and cycling female rats. Psychopharmacology, 179: 769-780.
CrossRefDirect Link - Al Humayed, S., F. Al-Hashem, M.A. Haidara, A.O. El Karib, S.S. Kamar, S.N. Amin and B. Al-Ani, 2020. Resveratrol pretreatment ameliorates p53-Bax axis and augments the survival biomarker B-cell lymphoma 2 modulated by paracetamol overdose in a rat model of acute liver injury. Pharmacology, 105: 39-46.
CrossRefDirect Link - Cenini, G., R. Sultana, M. Memo and D.A. Butterfield, 2008. Effects of oxidative and nitrosative stress in brain on p53 proapoptotic protein in amnestic mild cognitive impairment and Alzheimer disease. Free Radical Biol. Med., 45: 81-85.
CrossRefDirect Link - Gambino, V., G.D. Michele, O. Venezia, P. Migliaccio and V. Dall'Olio et al., 2013. Oxidative stress activates a specific p53 transcriptional response that regulates cellular senescence and aging. Aging Cell, 12: 435-445.
CrossRefDirect Link - Luo, Y., S. Kuang, L. Xue and J. Yang, 2016. The mechanism of 5-lipoxygenase in the impairment of learning and memory in rats subjected to chronic unpredictable mild stress. Physiol. Behav., 167: 145-153.
CrossRefDirect Link - Zhao, X., F. Cao, Q. Liu, X. Li and G. Xu et al., 2019. Behavioral, inflammatory and neurochemical disturbances in LPS and UCMS-induced mouse models of depression. Behavioural Brain Res., 364: 494-502.
CrossRefDirect Link - Moretti, M., J. Budni, D.B. dos Santos, A. Antunes and J.F. Daufenbach et al., 2013. Protective effects of ascorbic acid on behavior and oxidative status of restraint-stressed mice. J. Mol. Neurosci., 49: 68-79.
CrossRefPubMedDirect Link - Domingues, M., A.M. Casaril, P.T. Birmann, S.R. Bampi and D. de Andrade Lourenço et al., 2019. Effects of a selanylimidazopyridine on the acute restraint stress-induced depressive- and anxiety-like behaviors and biological changes in mice. Behav. Brain Res., 366: 96-107.
CrossRefDirect Link - Samad, N., A. Saleem, F. Yasmin and M.A. Shehzad, 2018. Quercetin protects against stress-induced anxiety- and depression-like behavior and improves memory in male mice. Physiol. Res., 67: 795-808.
CrossRefDirect Link - Bo, S., R. Gambino, V. Ponzo, I. Cioffi and I. Goitre et al., 2018. Effects of resveratrol on bone health in type 2 diabetic patients. A double-blind randomized-controlled trial. Nutr. Diabetes, Vol. 8.
CrossRefDirect Link - Liang, B. and J. Su, 2019. Inducible nitric oxide synthase (iNOS) mediates vascular endothelial cell apoptosis in grass carp reovirus (GCRV)-induced hemorrhage. Int. J. Mol. Sci., Vol. 20.
CrossRefDirect Link