
Li Tan
Department of Pharmacology, School of Pharmacy, Chengdu University of Traditional Chinese Medicine, 611137 Chengdu, People`s Republic of China
Nan Lei
Department of Pharmacology, School of Pharmacy, Chengdu University of Traditional Chinese Medicine, 611137 Chengdu, People`s Republic of China
Man He
Department of Pharmacology, School of Pharmacy, Chengdu University of Traditional Chinese Medicine, 611137 Chengdu, People`s Republic of China
Meng Zhang
Department of Pharmacology, School of Pharmacy, Chengdu University of Traditional Chinese Medicine, 611137 Chengdu, People`s Republic of China
Qiang Sun
Department of Pharmacology, School of Pharmacy, Chengdu University of Traditional Chinese Medicine, 611137 Chengdu, People`s Republic of China
Sha Zeng
Department of Pharmacology, School of Pharmacy, Chengdu University of Traditional Chinese Medicine, 611137 Chengdu, People`s Republic of China
Li Chen
Department of Pharmacology, School of Pharmacy, Chengdu University of Traditional Chinese Medicine, 611137 Chengdu, People`s Republic of China
Lijuan Zhou
Sichuan Academy of Chinese Medical Sciences, 610041 Chengdu, People`s Republic of China
Xianli Meng
Department of Pharmacology, School of Pharmacy, Chengdu University of Traditional Chinese Medicine, 611137 Chengdu, People`s Republic of China
Haibo Xu
Department of Pharmacology, School of Pharmacy, Chengdu University of Traditional Chinese Medicine, 611137 Chengdu, People`s Republic of China
LiveDNA: 86.30006
International Journal of Pharmacology
Year: 2020 | Volume: 16 | Issue: 1 | Page No.: 53-62







ABSTRACT
Background and Objective: It is well known that hedgehog signaling pathway plays a critical role in embryonic development, tissue patterning and carcinogenesis. Scutellarin, a bioactive component in Scutellaria barbata D. Don, is efficacious to various cancers with elusive mechanism to be defined. The objective of the present study was to examine the effect of scutellarin on human colorectal cancer and elaborate the underlying mechanism via hedgehog signaling pathway. Materials and Methods: After treatment of human colorectal carcinoma cells HCT116 with scutellarin or GANT61, an inhibitor of hedgehog signaling pathway activity as a positive control, at indicated concentrations, the effects of scutellarin or GANT61 on the proliferation, migration and anchorage-independent growth of HCT116 cells were assessed by MTT assay, wound healing assay and soft agar colony formation assay, followed by appraisal of the expressions of mRNA and protein of c-Myc, sonic hedgehog (SHh), Patched 1 (Ptch1) and Gli1 in HCT116 cells with quantitative RT-PCR and western blotting, respectively. Results: Scutellarin inhibited the proliferation, migration and anchorage-independent growth of HCT116 cells and mitigated the expressions of mRNA and protein of c-Myc, SHh, Ptch1 and Gli1 in HCT116 cells in a dose-dependent pattern. Furthermore, GANT61 exhibited strong suppression in above assays as well and the potency of scutellarin at 20 μg mL–1 was generally equal to that of GANT61 at 20 μM. Conclusion: These findings highlighted scutellarin as a promising therapeutic agent for human colorectal cancer, with potential down regulation of hedgehog signaling pathway activity.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Li Tan, Nan Lei, Man He, Meng Zhang, Qiang Sun, Sha Zeng, Li Chen, Lijuan Zhou, Xianli Meng and Haibo Xu, 2020. Scutellarin Protects against Human Colorectal Cancer in vitro by Down Regulation of Hedgehog Signaling Pathway Activity. International Journal of Pharmacology, 16: 53-62.
DOI: 10.3923/ijp.2020.53.62
URL: https://scialert.net/abstract/?doi=ijp.2020.53.62
DOI: 10.3923/ijp.2020.53.62
URL: https://scialert.net/abstract/?doi=ijp.2020.53.62
INTRODUCTION
Noncommunicable diseases (NCDs) are now responsible for the majority of global deaths. Cancer is a major public health problem worldwide and is the second leading cause of death in the United States. Colorectal cancer (CRC) is one of the most common malignancies in the world. In both sexes combined, colorectal cancer is the fourth most frequent cancer and the second leading cause of cancer death1. Therefore, prevention of cancer occurrence and high-quality health care for cancer patients is paramount to expand life expectancy.
The hedgehog signaling pathway was first discovered in a seminal study on embryonic development of Drosophila2. It has become a widely held view that the hedgehog signaling pathway is highly conserved and tightly controls not only organismal patterning and growth, but also adult tissue homeostasis in varieties of species. There are three types of ligands initiating hedgehog signaling, such as sonic hedgehog (SHh), Indian hedgehog and desert hedgehog, of which sonic hedgehog is most widely expressed and most potent in activating hedgehog signaling pathway3. Hedgehog ligands are secreted lipoproteins4, expressed as precursors mainly by certain epithelial cells and processed by autocatalytic cleavage of the C-terminal intein domain5, yielding an active N-terminal fragment retained6. Following binding to the 12-transmembrane receptors, such as Patched 1 (Ptch1) and Patched 2 (Ptch 2)7-9, active hedgehog ligands stimulate the hedgehog pathway, giving rise to relieving suppression of a transmembrane protein named Smoothened (SMO) by PTCH receptor. Rescued SMO protein accumulates to destroy the repression of suppressor of fused homolog (SUFU) on Gli proteins, which are transcription factors with three isoforms including Gli1, Gli2 and Gli3. It is well established that Gli1 is a dominant activator of hedgehog transcriptional program10. Ultimately, dissociated Gli proteins translocate into cellular nuclei to promote expressions of downstream genes, including hedgehog-interacting protein (HIP), Patched and proto-oncogene c-Myc11,12.
To date, it is being brought to light that hedgehog signaling pathway is greatly associated with tumorigenesis, metastases and malignancies of various cancer types, including basal cell carcinoma (BCC), medulloblastoma, rhabdomyosarcoma, pancreatic ductal adenocarcinoma, lung cancer, prostate cancer and colorectal cancer, etc13,14.
Scutellarin is a type of flavonoid glucuronide with multiple pharmacological activities and widely exists in a vast array of natural herbs. Scutellarin was first discovered as a dominant chemical constituent in the breviscapine of Erigeron breviscapus (Vant.) Hand-Mazz. which is now applied in clinical settings for cardiovascular disorders and cancer in China15. Scutellarin is also contained in high amount in Scutellaria barbata D. Don, a traditional Chinese medicinal herb for cancer treatment with high performance16. Accumulating reports indicated that scutellarin shows strong beneficial activity against a number of cancer types, including human colorectal cancer, hepatocellular carcinoma, lymphoma, human prostate cancer and tongue squamous carcinoma16. However, the underlying mechanism by which scutellarin protects against cancer, in particular colorectal cancer, is yet to be elucidated. In the current study, pharmacological effect of scutellarin on colorectal cancer cells was investigated, providing an insight via hedgehog signaling pathway into the in-depth mechanism.
MATERIALS AND METHODS
This study was carried out in the Department of Pharmacology, School of Pharmacy, Chengdu University of Traditional Chinese Medicine, from January, 2019 to July, 2019.
Drugs and reagents: Human colorectal carcinoma cells HCT116 was originally derived from American Type Culture Collection (Manassas, VA, USA). Scutellarin (C21H18O12) at a purity of over 98% supplied by Must Biotechnology Corporation Ltd. (Chengdu, China) was dissolved in dimethyl sulfoxide (DMSO) and stored in the dark at 4. The GANT61, an inhibitor of hedgehog signaling pathway activity as a positive control, was purchased from MedChemExpress (New Jersey, USA), dissolved in DMSO and stored in aliquots at -20°C. Prior to the assays below, scutellarin and GANT61 were diluted with cell culture medium to indicated final concentrations, with less than 0.1% DMSO contained in the complete medium. The MTT cell proliferation kit was purchased from Beyotime Biotechnology Inc. (Shanghai, China). Multisource RNA Miniprep Kit was bought from Axygen (Massachusetts, USA). PrimeScript™ RT reagent Kit and SYBR® Premix DimerEraser™ were both provided by Takara Biomedical Technology (Beijing, China). Antibodies to sonic hedgehog (SHh), Gli1, c-Myc and GAPDH were obtained from Santa Cruz Biotechnology Inc. (Dallas, Texas, USA). All other reagents were readily available commercially.
Cell culture and treatment: The HCT116 cells were maintained with McCoy's 5A medium supplemented with 10% of heat-inactivated fetal bovine serum (FBS), 0.22% sodium bicarbonate, 100 U mL–1 of penicillin, 100 μg mL–1 of streptomycin. Approximately 24 h post-seeding in culture plate, HCT116 cells were treated with scutellarin or GANT61 at a range of concentrations for various spells, followed by evaluation of cell proliferation, cell migration, soft agar colony formation, expressions of mRNA and protein in HCT116 cells, as detailed below.
MTT assay of cell proliferation: The MTT assay of cell proliferation was conducted twice in quadruplicate, as previously described17. Briefly, after treatment of HCT116 cells grown in a 96-well plate with scutellarin or GANT61, MTT solution in a volume equal to 10% of the culture liquid was aseptically added, followed by incubation for 4 h. After removal and disposal of the culture fluid, MTT solvent in an amount equal to the original culture medium volume was added, followed by gentle stirring in a gyratory shaker to dissolve the MTT formazan crystals and colorimetric readout with a spectrophotometer (Thermo Scientific Varioskan Flash, Type 3001) set to 570 nm.
Wound healing assay: Wound healing assay was employed to appraise cell migration, as described elsewhere18, with minor modification. The HCT116 cells were seeded into a 24-well tissue culture plate and grown to 80% confluence as a monolayer. A straight would line was scratched on the cell monolayer with a p200 pipette tip under an angle of around 30 degrees, followed by gentle wash twice to remove the detached cells. The HCT116 cells were maintained with fresh culture medium containing 1% FBS, scutellarin or GANT61 for additional 24 h, with concomitant capture of migration images using the 10× objective of a microscope at time 0, 12 and 24 h. Cell migration was analyzed with ImageJ 1.48v. The relative migration index was represented in terms of the percentage of migrated cell surface area in total surface area. The experiment was carried out in quadruplicate and repeated 2 times.
Soft agar colony formation assay: Following 0.5 mL of 1.4% agarose was mixed with 0.5 mL of cell culture medium (2×) containing scutellarin or GANT61 at various concentrations, the mixture was added to one well of a 6-well plate and set for a base layer. After 0.5 mL of 0.7% agarose was mixed with 0.5 mL of cell culture medium (2×) containing scutellarin or GANT61 at various concentrations and 5000 cells, the mixture was seeded on the base layer and set for a top layer, followed by incubation at 37°C with 5% CO2 and 95% humidity for 2 weeks. Then the colonies were stained with 0.01% crystal violet, photographed and counted under a microscope. The assay was performed twice in quadruplicate19.
Quantitative RT-PCR analysis: After treatment with scutellarin or GANT61 for 48 h, HCT116 cells were harvested and processed for total RNA isolation with a Multisource RNA Miniprep Kit (Axygen), followed by reverse transcription with a PrimeScript™ RT reagent Kit (Takara) and quantitative real-time polymerase chain reaction with a SYBR® Premix DimerEraser™ kit (Takara) on Bio-Rad iCycler iQ Real-Time PCR Detection System in a 96-well plate format, in light of the manufacturer's protocol. β-actin was used as an endogenous reference gene to generate normalized relative expression values for the genes of interest20. Primer sequences for specific genes are shown in Table 1. Fold differences in the values for the treated group relative to the control is analyzed with the comparative Ct method (△△Ct algorithm). Assays were run in 3 replicates.
| Table 1: | Primer sequences for qRT-PCR |
 | |
Western blot analysis: After treatment with scutellarin or GANT61 for 72 h, HCT116 cells were harvested and processed for protein extraction, followed by BCA Protein Assay (Pierce, #23227) for protein quantitation as per the user manual. Cell lysates were separated with sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS–PAGE) and transferred to a PVDF membrane. The membranes were blocked with 5% non-fat milk and probed with primary antibodies against SHh (50 KDa) (Proteintech Group, Inc), Gli1 (160 KDa) (Cell Signaling Technology, Inc.), c-Myc (48.8 KDa) (MultiSciences Biotech Co,. Ltd.) and GAPDH (37KDa) (Zen Bio Science, Inc.) as a whole cell loading control at 1:1000 dilution. After incubation with horseradish peroxidase (HRP)-conjugated secondary antibodies at 1:2000 dilution, proteins were detected with ECLTM western blotting detection reagents (GE Healthcare, #RPN2209), imaged on ChemiDoc XRS+ system (Bio-Rad, Hercules, California, USA) and analyzed with Bio-Rad Quantity One 1-D analysis software20. The assay was carried out in quadruplicate.
Statistical analysis: The data were analyzed using one-way analysis of variance (ANOVA) followed by Student's t-test with SPSS 19.0 software. The results are presented as Mean±SEM (standard error of the mean) unless otherwise indicated. A 2-sided value of p<0.05 was considered statistically significant.
RESULTS
Scutellarin suppresses the proliferation of HCT116 cells: The effect of scutellarin on the proliferation of HCT116 cells was first measured. After treatment of HCT116 cells with scutellarin or GANT61 for 24 h as shown in Fig. 1a, scutellarin at concentrations of 10 and 20 μg mL–1 and GANT61 at 20 μM all significantly inhibited the proliferation of HCT116 cells and the effect of scutellarin at 20 μg mL–1 was statistically equal to that of GANT61. Following 48 h of treatment, the proliferation of HCT116 cells was heavily mitigated by scutellarin at concentrations of 5, 10 and 20 μg mL–1 and GANT61 at 20 μM. The potency of scutellarin was in a dose-dependent mode and no difference in the efficacy between scutellarin at 20 μg mL–1 and GANT61 was appreciable (Fig. 1b).
Scutellarin retards the migration of HCT116 cells: Migration is the 1st step of the more complex process of tumor cell invasion. Using the wound healing assay, the migratory activity of HCT116 cells treated with scutellarin was assessed. As shown in Fig. 2a and c, scutellarin at concentrations of 5, 10 and 20 μg mL–1 and GANT61 at 20 μM all significantly retarded the migration of HCT116 cells treated for 12 h, compared with the control.
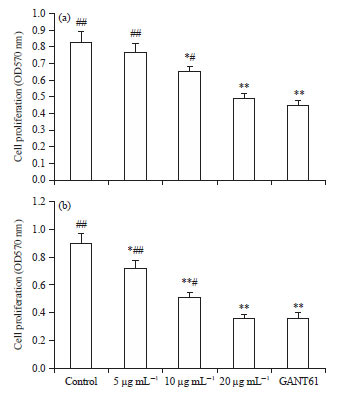 | |
| Fig. 1(a-b): | Scutellarin suppresses the proliferation of HCT116 cells. (a) HCT116 cells with scutellarin at concentrations of 5, 10 and 20 μg mL–1 or GANT61 at 20 μM for 24 h and (b) 48 h |
Data are the Mean±SEM of 8 separate experiments, *p<0.05, **p<0.01 vs. the control, #p<0.05, ##p<0.01 vs. GANT61 | |
In addition, there was no difference in retardation between 20 μg mL–1 of scutellarin and 20 μM of GANT61. In comparison with the vehicle-treated control, treatment of HCT116 cells with scutellarin at a variety of concentrations or GANT61 for 24 h drastically diminished the migration of HCT116 cells. Furthermore, attenuation of cell migration by scutellarin appeared to be dose dependent and no difference in the effectiveness between scutellarin at 20 μg mL–1 and GANT61 at 20 μM was noted (Fig. 2b, c).
Scutellarin impedes the colony formation of HCT116 cells: As anchorage-independent growth monitored by soft agar colony formation assay is a capability of cellular malignant transformation, the potency of scutellarin on malignant transformation in HCT116 cells was then evaluated. The soft agar colony formation of HCT116 cells was sharply impeded by treatment for 2 weeks with scutellarin at concentrations of 5, 10 and 20 μg mL–1 or GANT61 at 20 μM, compared with the control. Moreover, this action of scutellarin was in a dose-dependent pattern, with no difference in impediment between scutellarin at 20 μg mL–1 and GANT61 at 20 μM (Fig. 3a, b).
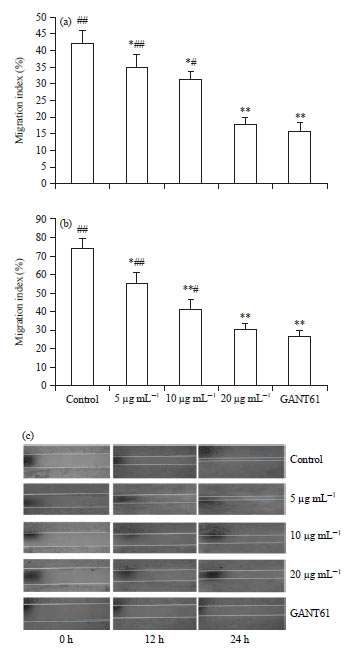 | |
| Fig. 2(a-c): | Scutellarin retards the migration of HCT116 cells. (a) HCT116 cells with scutellarin at concentrations of 5, 10 and 20 μg mL–1 or GANT61 at 20 μM for 12 h, (b) 24 h and (c) Representative images from 8 separate experiments with similar results |
Data are the Mean±SEM of 8 independent tests, *p<0.05, **p<0.01 vs. the control, #p<0.05, ##p<0.01 vs. GANT61 | |
Scutellarin lessens the mRNA level in HCT116 cells: Next, whether scutellarin influenced the expressions of cancer-associated genes in HCT116 cells was ascertained. Following treatment of HCT116 cells with scutellarin for 48 h, the level of c-Myc mRNA was reduced by scutellarin at 5, 10 and 20 μg mL–1, in a dose-dependent manner, with the potential of scutellarin at 20 μg mL–1 equal to that of GANT61 at 20 μM (Fig. 4a).
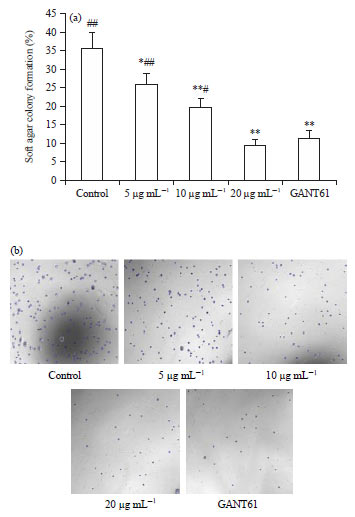 | |
| Fig. 3(a-b): | Scutellarin impedes the colony formation of HCT116 cells. (a) HCT116 cells with scutellarin at concentrations of 5, 10 and 20 μg mL–1 or GANT61 at 20 μM for 2 weeks, soft agar colony formation assay was done as detailed in the materials and methods section and (b) Images from 8 independent experiments with similar results |
Data are the Mean±SEM of 8 separate experiments, *p<0.05, **p<0.01 vs. the control, #p<0.05, ##p<0.01 vs. GANT61 | |
In comparison with the vehicle-treated control, scutellarin at concentrations of 10 and 20 μg mL–1 significantly decreased SHh mRNA expression, with 20 μg mL–1 of scutellarin more efficacious than 20 μM of GANT61 (Fig. 4b). Additionally, scutellarin manifested weak attenuation of Ptch1 mRNA transcription, which was just alleviated by scutellarin at 20 μg mL–1 and GANT61 at 20 μM and no difference in suppression between these 2 groups was observed (Fig. 4c). However, Gli1 mRNA level in HCT116 cells was significantly eliminated by scutellarin at 5, 10 and 20 μg mL–1, in a dose-dependent fashion and drastically by GANT61 at 20 μM (Fig. 4d).
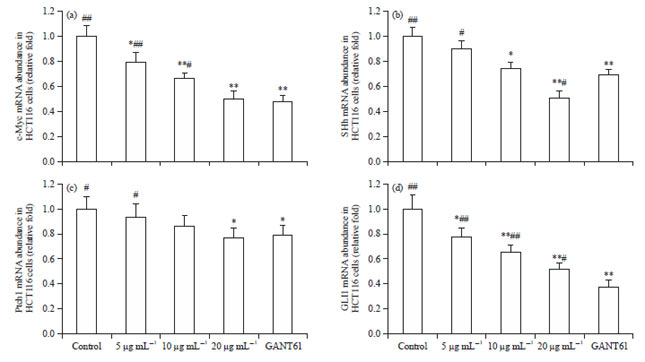 | |
| Fig. 4(a-d): | Scutellarin lessens the mRNA level in HCT116 cells. After treatment of HCT116 cells with scutellarin at concentrations of 5, 10 and 20 μg mL–1 or GANT61 at 20 μM for 48 h, qRT-PCR was performed to analyze the (a) mRNA level of c-Myc, (b) SHh, (c) Ptch1 and (d) GLI1 |
Data are the Mean±SEM of 6 separate experiments, *p<0.05, **p<0.01 vs. the control, #p<0.05, ##p<0.01 vs. GANT61 | |
Scutellarin lowers the protein level in HCT116 cells: To validate the action of scutellarin on the mRNA expressions of cancer-implicated genes, the function of scutellarin on protein level in HCT116 cells was determined. After treatment of HCT116 cells for 72 h with scutellarin at 5, 10 and 20 μg mL–1 or GANT61 at 20 μM, scutellarin dose dependently mitigated c-Myc protein translation, with equality in mitigation between 20 μg mL–1 of scutellarin and 20 μM of GANT61 (Fig. 5a, d). Compared with the control, SHh protein expression was significantly hindered by scutellarin at 10 and 20 μg mL–1 and 20 μM of GANT61, with 20 μg mL–1 of scutellarin more effective than 20 μM of GANT61 (Fig. 5b, d). Moreover, scutellarin at concentrations of 5, 10 and 20 μg mL–1 and GANT61 at 20 μM hugely diminished the expression of Gli1 protein in HCT116 cells and the efficacy of scutellarin revealed to be in a dose-dependent pattern (Fig. 5c, d).
DISCUSSION
Mounting evidence indicates that traditional Chinese medicine may be superior to western medicine in the prevention and treatment of many types of diseases20,21, particularly including cancer17, generally attributed to its less adverse reaction and acting on multiple pathological targets. Scutellarin, a major active flavonoid compound in a traditional Chinese medicinal herb Scutellaria barbata D. Don, is efficacious to varieties of cancers, with the underlying mechanism to be illuminated.
In the study, scutellarin displayed impediment to the proliferation of HCT116 cells in a dose-dependent fashion. This type of function of scutellarin was concurrent with other reports that scutellarin inhibits the proliferation of human hepatocellular carcinoma HepG2 Cells and MHCC97-H cells22,23, non-small cell lung cancer (NSCLC) A549 cells24, renal cell carcinoma (RCC) ACHN cells and 786-O cells25, breast carcinoma MCF-7 cells26 and human prostate cancer PC3 cells27.
Subsequently, whether scutellarin influenced the behavior of colorectal cancer cells was explored. It was discovered that scutellarin dose dependently retarded the migration of HCT116 cells. This result was in accordance with previous statements that scutellarin encumbers the migration of human hepatocellular carcinoma HepG2 Cells22, oral squamous cell carcinoma (OSCC) HSC-4 cells and SAS cells28 and malignant melanoma A375 cells29.
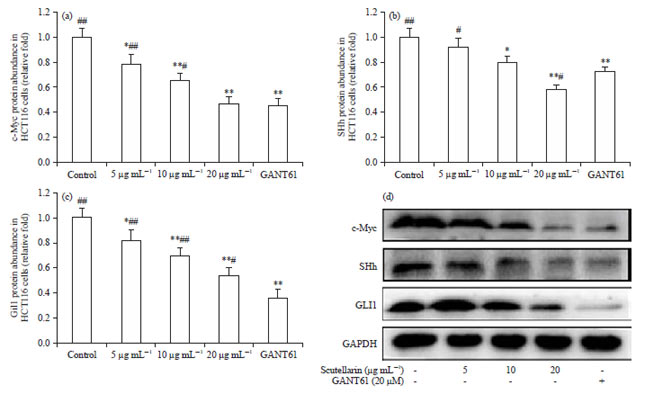 | |
| Fig. 5(a-d): | Scutellarin lowers the protein level in HCT116 cells. After treatment of HCT116 cells with scutellarin at concentrations of 5, 10 and 20 μg mL–1 or GANT61 at 20 μM for 72 h, western blotting was conducted to appraise the (a) Protein level of c-Myc, (b) Shh, (c) GLI1 and (d) Original blots representative of 4 similar experiments, with GAPDH as a loading control |
Data are the Mean±SEM of 4 independent experiments, *p<0.05, **p<0.01 vs. the control, #p<0.05, ##p<0.01 vs. GANT61 | |
Furthermore, clonogenic survival effect of scutellarin on HCT116 cells was detected. Scutellarin markedly diminished the numbers of non-adherent sphere cells, which coincided with other literatures that scutellarin hinders soft agar colony formation of NSCLC A549 cells24, human hepatocellular carcinoma HepG2 Cells and MHCC97-H cells23, RCC ACHN cells and 786-O cells25, human prostate cancer PC3 cells27 and breast carcinoma MCF-7 cells26.
Since the alterations to cellular phenotype, including proliferation, migration and colony formation, are generally the manifestations of cellular genotype changes, if scutellarin influenced the expressions of some genes in HCT116 cells was next pursued. In the study, the mRNA transcription and protein translation of c-Myc in HCT116 cells were both largely attenuated by scutellarin in a mode of dose dependence. It is well known that c-Myc is a proto-oncogene coding for a transcription factor, which can activate the expressions of downstream target oncogenes, giving rise to carcinogenesis30. Constitutive upregulation of c-Myc is commonly observed in carcinoma of the colon, stomach, cervix, breast and lung31,32, thus c-Myc is viewed as a promising target for anti-cancer drugs and down regulation of c-Myc level by scutellarin is quite beneficial for therapeutic intervention in colorectal cancer. Furthermore, over expression of c-Myc is also associated with cancer stem cells-initiated malignancies, scutellarin may possibly exert anti-cancer property via repression of the self-renewal and differentiation of cancer stem cells33.
A growing body of evidence suggested that hedgehog signaling pathway is implicated in cancer stem cell maintenance, carcinogenesis, progression and invasiveness34. Activation of hedgehog signaling pathway by binding of hedgehog ligand to patched receptor eventually renders Gli protein to translocate into cellular nuclei and heighten the expressions of downstream genes including c-Myc and some key components of hedgehog signaling pathway, such as Patched and HIP35. The importance of hedgehog signaling pathway in cancerous pathogenesis places the novel drug targeting hedgehog pathway in high demand. Therefore, whether scutellarin affected the activity of hedgehog signaling pathway in colorectal cancer was quested. Intriguingly, scutellarin diminished SHh, Ptch1 and Gli1 expression in HCT116 cells in a dose-dependent manner, suggesting that down regulation of hedgehog pathway activity is potentially the underlying mechanism by which scutellarin protects against colorectal cancer.
In the study, GANT61 was employed as a positive control. GANT61 is a compound with inhibitory activity for Gli1 as well as Gli2-induced transcription, by reduction of Gli1/2 DNA binding. It was documented that GANT61 efficiently suppresses tumor growth and proliferation in a Gli-dependent manner36. In the study, GANT61 hugely mitigated the proliferation, migration, transformation and the expressions of c-Myc, SHh, Gli1 and Ptch1 in HCT116 cells, which was consistent with a number of other data regarding the pharmacodynamic property37 of GANT61. In the above assays, the efficacy of scutellarin at 20 μg mL–1 was statistically equivalent to that of GANT61 at 20 μM, except weaker on Gli1 expression and stronger on SHh expression. Given that some patients receiving GANT61 report serious adverse events and non-durable response, scutellarin may be more advantageous than GANT61 in terms of cancer treatment.
On the whole, signal transduction in the context of hedgehog pathway is quite complex and many other molecules, for instance Ptch2, Smoothened, SUFU, Gli2 and Gli3, also play vital roles in modulation of hedgehog signaling pathway activity. The effects of scutellarin on these molecular targets need to be addressed to clarify the in-depth mechanism for cancer treatment. Besides canonical hedgehog signaling pathway, non-canonical hedgehog signaling pathway discovered in recent years takes a complementary role in carcinogenesis38, on which the impact of scutellarin is in urgent need of illumination.
CONCLUSION
Taken together, scutellarin may protect against human colorectal cancer, potentially by down regulation of hedgehog signaling pathway activity, as evidenced by its abilities of reducing the proliferation, migration and anchorage-independent growth of HCT116 cells and diminishing the expressions of c-Myc, SHh, Ptch1 and Gli1 in HCT116 cells.
SIGNIFICANCE STATEMENT
Traditional Chinese medicine since ancient times has utilized a large arsenal of natural products which nowadays are regarded as potential drug sources, from which scutellarin is a promising anti-cancer agent by down regulation of hedgehog signaling pathway activity. As lead compounds targeting hedgehog signaling cascade are of particular interest, substantial efforts should be made to develop new agents, such as scutellarin, targeting various components of hedgehog signaling pathway. With intensive delineation of the pharmacological mechanisms, utilization of scutellarin for therapeutic intervention in clinical cancer will be further underpinned in the foreseeable future. This study demonstrated the association of hedgehog signaling cascade with colorectal cancer and may provide a therapeutic strategy for various cancers by down regulation of hedgehog signaling pathway activity in clinical settings.
ACKNOWLEDGMENT
This work was sponsored by National Natural Science Foundation of China (Nos. 81173598 and 81573813), the Excellent Talent Program of Chengdu University of Traditional Chinese Medicine (No. YXRC2019002) and the Fund of Scientific Research Innovation Team Construction in Sichuan Provincial University (No. 18TD0017).
REFERENCES
- Bray, F., J. Ferlay, I. Soerjomataram, R.L. Siegel, L.A. Torre and A. Jemal, 2018. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: Cancer J. Clin., 68: 394-424.
CrossRefDirect Link - Nusslein-Volhard, C. and E. Wieschaus, 1980. Mutations affecting segment number and polarity in Drosophila. Nature, 287: 795-801.
CrossRefPubMedDirect Link - Carpenter, R.L. and H. Ray, 2019. Safety and tolerability of sonic hedgehog pathway inhibitors in cancer. Drug Saf., 42: 263-279.
CrossRefPubMedDirect Link - McMahon, A.P., P.W. Ingham and C.J. Tabin, 2003. Developmental roles and clinical significance of hedgehog signaling. Curr. Top. Dev. Biol., 53: 1-114.
CrossRefDirect Link - Perler, F.B., 1998. Protein splicing of inteins and hedgehog autoproteolysis: Structure, function and evolution. Cell, 92: 1-4.
CrossRefPubMedDirect Link - Porter, J.A., D.P. von Kessler, S.C. Ekker, K.E. Young, J.J. Lee, K. Moses and P.A. Beachy, 1995. The product of hedgehog autoproteolytic cleavage active in local and long-range signalling. Nature, 374: 363-366.
CrossRefDirect Link - Marigo, V., R.A. Davey, Y. Zuo, J.M. Cunningham and C.J. Tabin, 1996. Biochemical evidence that patched is the Hedgehog receptor. Nature, 384: 176-179.
CrossRefDirect Link - Stone, D.M., M. Hynes, M. Armanini, T.A. Swanson and Q. Gu et al., 1996. The tumour-suppressor gene patched encodes a candidate receptor for Sonic hedgehog. Nature, 384: 129-134.
CrossRefDirect Link - Smyth, I., M.A. Narang, T. Evans, C. Heimann and Y. Nakamura et al., 1999. Isolation and characterization of human patched 2 (PTCH2), a putative tumour suppressor gene in basal cell carcinoma and medulloblastoma on chromosome 1p32. Hum. Mol. Genet., 8: 291-297.
CrossRefDirect Link - Carballo, G.B., J.R. Honorato, G.P.F. de Lopes and T. Spohr, 2018. A highlight on Sonic hedgehog pathway. Cell Commun. Signal., Vol. 16.
CrossRefDirect Link - Bariwal, J., V. Kumar, Y. Dong and R.I. Mahato, 2019. Design of Hedgehog pathway inhibitors for cancer treatment. Med. Res. Rev., 39: 1137-1204.
CrossRefDirect Link - Sari, I.N., L.T.H. Phi, N. Jun, Y.T. Wijaya, S. Lee and H.Y. Kwon, 2018. Hedgehog signaling in cancer: A prospective therapeutic target for eradicating cancer stem cells. Cells, Vol. 7, No. 11.
CrossRefDirect Link - Raleigh, D.R. and J.F. Reiter, 2019. Misactivation of Hedgehog signaling causes inherited and sporadic cancers. J. Clin. Invest., 129: 465-475.
CrossRefDirect Link - Wu, C., X. Zhu, W. Liu, T. Ruan and K. Tao, 2017. Hedgehog signaling pathway in colorectal cancer: Function, mechanism and therapy. Onco Targets Ther., 10: 3249-3259.
CrossRefPubMedDirect Link - Wang, L. and Q. Ma, 2018. Clinical benefits and pharmacology of scutellarin: A comprehensive review. Pharmacol. Ther., 190: 105-127.
CrossRefDirect Link - Chledzik, S., J. Strawa, K. Matuszek and J. Nazaruk, 2018. Pharmacological effects of scutellarin, an active component of genus Scutellaria and Erigeron: A systematic review. Am. J. Chin. Med., 46: 319-337.
CrossRefDirect Link - Wan, S., L. Tan, N. Lei, Y. Shi and M. He et al., 2018. Compound bieshe kangai inhibits proliferation and induces apoptosis in HCT116 human colorectal cancer cells. Trop. J. Pharm. Res., 17: 2163-2168.
CrossRefDirect Link - Xu, H., M. McCann, Z. Zhang, G.H. Posner, V. Bingham, M. El-Tanani and F.C. Campbell, 2009. Vitamin D receptor modulates the neoplastic phenotype through antagonistic growth regulatory signals. Mol. Carcinog., 4: 758-772.
CrossRefDirect Link - Xu, H., D. Wang, C. Peng, X. Huang and M. Ou et al., 2014. Rabbit sera containing compound danshen dripping pill attenuate leukocytes adhesion to TNF-alpha–activated human umbilical vein endothelial cells by suppressing endothelial ICAM-1 and VCAM-1 expression through NF-kappaB signaling pathway. J. Cardiovasc. Pharmacol., 63: 323-332.
CrossRefDirect Link - Xu, H., Q. Wu, C. Peng and L. Zhou, 2013. Study on the antiviral activity of San Huang Yi Gan Capsule against hepatitis B virus with seropharmacological method. BMC Complem. Altern. Med., Vol.13.
Direct Link - Ke, Y., T. Bao, X. Wu, H. Tang and Y. Wang et al., 2017. Scutellarin suppresses migration and invasion of human hepatocellular carcinoma by inhibiting the STAT3/Girdin/Akt activity. Biochem. Biophys. Res. Commun., 483: 509-515.
CrossRefDirect Link - Liu, K., T. Tian, Y. Zheng, L. Zhou and C. Dai et al., 2019. Scutellarin inhibits proliferation and invasion of hepatocellular carcinoma cells via down-regulation of JAK2/STAT3 pathway. J. Cell. Mol. Med., 23: 3040-3044.
CrossRefDirect Link - Cao, P., B. Liu, F. Du, D. Li and Y. Wang et al., 2019. Scutellarin suppresses proliferation and promotes apoptosis in A549 lung adenocarcinoma cells via AKT/mTOR/4EBP1 and STAT3 pathways. Thorac. Cancer., 10: 492-500.
CrossRefDirect Link - Deng, W., W. Han, T. Fan, X. Wang, Z. Cheng, B. Wan and J. Chen, 2018. Scutellarin inhibits human renal cancer cell proliferation and migration via upregulation of PTEN. Biomed. Pharmacother., 107: 1505-1513.
CrossRefDirect Link - Hou, L., L. Chen and L. Fang, 2017. Scutellarin inhibits proliferation, invasion, and tumorigenicity in human breast cancer cells by regulating HIPPO-YAP signaling pathway. Med. Sci. Monit., 23: 5130-5138.
CrossRefPubMedDirect Link - Gao, C., Y. Zhou, Z. Jiang, Y. Zhao and D. Zhang et al., 2017. Cytotoxic and chemosensitization effects of Scutellarin from traditional Chinese herb Scutellaria altissima L. in human prostate cancer cells. Oncol. Rep., 38: 1491-1499.
CrossRefDirect Link - Li, H., D. Huang, Z. Gao, Y. Lv, L. Zhang, H. Cui and J. Zheng, 2010. Scutellarin inhibits cell migration by regulating production of αvβ6 integrin and E-cadherin in human tongue cancer cells. Oncol. Rep., 24: 1153-1160.
CrossRefDirect Link - Li, C.Y., Q. Wang, X. Wang, G. Li, S. Shen and X. Wei, 2019. Scutellarin inhibits the invasive potential of malignant melanoma cells through the suppression epithelial-mesenchymal transition and angiogenesis via the PI3K/Akt/mTOR signaling pathway. Eur. J. Pharmacol., Vol. 858.
CrossRefDirect Link - Caforio, M., C. Sorino, S. Iacovelli, M. Fanciulli, F. Locatelli and V. Folgiero, 2018. Recent advances in searching c-Myc transcriptional cofactors during tumorigenesis. J. Exp. Clin. Cancer Res., Vol. 37.
CrossRefDirect Link - Kumar, D., N. Sharma and R. Giri, 2017. Therapeutic interventions of cancers using intrinsically disordered proteins as drug targets: c-Myc as model system. Cancer Inform., 16: 1-7.
CrossRefDirect Link - Sipos, F., G. Firneisz and G. Muzes, 2016. Therapeutic aspects of c-MYC signaling in inflammatory and cancerous colonic diseases. World J. Gastroenterol., 22: 7938-7950.
CrossRefPubMedDirect Link - Elbadawy, M., T. Usui, H. Yamawaki and K. Sasaki, 2019. Emerging roles of C-Myc in cancer stem cell-related signaling and resistance to cancer chemotherapy: A potential therapeutic target against colorectal cancer. Int. J. Mol. Sci., Vol. 20.
CrossRefDirect Link - Cortes, J.E., R. Gutzmer, M.W. Kieran and J.A. Solomon, 2019. Hedgehog signaling inhibitors in solid and hematological cancers. Cancer Treat. Rev., 76: 41-50.
CrossRefDirect Link - Niyaz, M., K.S. Khan and S. Mudassar, 2019. Hedgehog signaling: An Achilles' heel in cancer. Transl. Oncol., 12: 1334-1344.
CrossRefDirect Link - Stanton, B.Z. and L.F. Peng, 2010. Small-molecule modulators of the Sonic Hedgehog signaling pathway. Mol. BioSyst., 6: 44-54.
CrossRefDirect Link - Wu, J., D. Di, C. Zhao, Y. Liu and H. Chen et al., 2018. Role of glioma-associated GLI1 oncogene in carcinogenesis and cancer-targeted therapy. Curr. Cancer Drug Targets, 18: 558-566.
CrossRefDirect Link - Pietrobono, S., S. Gagliardi and B. Stecca, 2019. Non-canonical hedgehog signaling pathway in cancer: Activation of GLI transcription factors beyond smoothened. Front. Genet., Vol. 10.
CrossRefDirect Link