
Xiaoying Duan
Third Grade Laboratory of State Administration of Traditional Chinese Medicine, The First Affiliated Hospital of Henan University of Traditional Chinese Medicine, Zhengzhou, China
LiveDNA: 86.29774
Yuanfang Cheng
Third Grade Laboratory of State Administration of Traditional Chinese Medicine, The First Affiliated Hospital of Henan University of Traditional Chinese Medicine, Zhengzhou, China
Feng Sang
Third Grade Laboratory of State Administration of Traditional Chinese Medicine, The First Affiliated Hospital of Henan University of Traditional Chinese Medicine, Zhengzhou, China
Lina Liu
Third Grade Laboratory of State Administration of Traditional Chinese Medicine, The First Affiliated Hospital of Henan University of Traditional Chinese Medicine, Zhengzhou, China
Zhen Liu
Third Grade Laboratory of State Administration of Traditional Chinese Medicine, The First Affiliated Hospital of Henan University of Traditional Chinese Medicine, Zhengzhou, China
Lin Cui
Third Grade Laboratory of State Administration of Traditional Chinese Medicine, The First Affiliated Hospital of Henan University of Traditional Chinese Medicine, Zhengzhou, China
Haixia Li
School of Pharmaceutical Sciences, Zhengzhou University, Zhengzhou, China
International Journal of Pharmacology
Year: 2020 | Volume: 16 | Issue: 6 | Page No.: 470-478







ABSTRACT
Background and Objective: A novel delivery system of matrine (MT) using Wheat Germ Agglutinin (WGA), an ideal bio-adhesive material, had been developed by us. The present work was aimed to evaluate the targeting function, anti-colon cancer efficacy and mechanisms of this novel delivery system on cell level. Materials and Methods: After WGA-modified matrine nanoparticles (WGA-MT-NPs) were prepared with our optimizedmethods, the Encapsulation Efficiency (EE) of MT and modification rate of WGA were measured by HPLC and folin-phenol method, respectively. Subsequently, Transmission Electron Microscopy (TEM) was used to further confirm the modification of WGA and HT-29 human colon cancer cells and NCM460 normal colonic epithelial cells were applied to assess the targeting function of WGA-MT-NPs. Furthermore, in HT-29 cells, the cell growth inhibition was determined by tetrazolium salt reagent (MTT) method and the apoptosis and cell cycle arrest were measured usingflowcytometry assay. Results: The EE and modification rate of WGA were 89.99±5.08% and 87.57±8.61%, respectively. And, TEM revealed that WGA was effectively assembled on the surfaces of nanostructures. Moreover, WGA-MT-NPs possessed the specific affinity towards HT-29 cells, rather than NCM460 cells. In HT-29 cells, the growth inhibition rate was significantly improved by WGA-MT-NPs compared with MT-NPs. Furthermore, the apoptosis of HT-29 cells was significantly higher in WGA-MT-NPs than in MT-NPs. Conclusion: WGA-MT-NPs significantly increased the anti-tumor effect of MT-NPs via stronger affinity towards HT-29 cells and higher promotion of apoptosis. Thus, WGA-MT-NPs may provide a promising new strategy for anti-colon cancer.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Xiaoying Duan, Yuanfang Cheng, Feng Sang, Lina Liu, Zhen Liu, Lin Cui and Haixia Li, 2020. Enhanced Targeting Function and Anti-colon Cancer Efficacy by Wheat Germ Agglutinin-modified Nanoparticles for Matrine Delivery. International Journal of Pharmacology, 16: 470-478.
DOI: 10.3923/ijp.2020.470.478
URL: https://scialert.net/abstract/?doi=ijp.2020.470.478
DOI: 10.3923/ijp.2020.470.478
URL: https://scialert.net/abstract/?doi=ijp.2020.470.478
INTRODUCTION
Colorectal Cancer (CRC) is the fourth most common cancer for morbidity and the second most common for mortality in human beings1. Matrine (MT), a natural alkaloid, has favorable anti-colon cancer effect, such as inhibiting the proliferation and inducing the apoptosis on SW1116, HT-29, LoVo and SW480 colon cancer cells2-5. However, due to the poor permeability of MT across the intestinal epithelial cells6, the absolute oral bioavailability of MT is only 17% in rat7, which hinders MT from achieving its full potential as an anti-colon cancer agent8.
The aberrant protein glycosylation of mucin in intestinal mucosal tissue is a frequent pathological alteration associated with the onset, progression and poor prognosis of CRC9,10. Wheat Germ Agglutinin (WGA), one of the lectins which are a family of glycan-specific proteins and relatively non-toxic and inexpensive10, can specifically recognize and bind to sugar moieties on cell surfaces of mucus and further to become the “glyco” portion of glycoconjugates on the cell membranes11,12. Thus, WGA is the ideal bio-adhesive material with target-finding properties and ability of reaching the underlying epithelia13,14 and has been used to predict the presence of a colorectal neoplasm9,10. We postulated that modifying WGA on matrine nanoparticles would overcome the MT’s problem of poor permeability to enhance its anti-coloneffect.
In the present study, based on our previous report of a novel delivery system for MT using WGA, we further confirmed the modification of WGA by TEM. Then, targeting function, the antitumor efficacy and mechanisms of the system were evaluated on cell level for exploring whether WGA could enhance the antitumor effect of MT-NPs as expected.
MATERIALS AND METHODS
Materials: Polyvinyl Alcohol (PVA), Wheat Germ Agglutinin (WGA), Carbodiimide (EDAC) N-hydroxysuccinimide (NHS), Folin-Ciocalteu's phenol Reagent, poly (lactic-co-glycolic acid) (PLGA), matrine (MT) and tetrazolium salt reagent (MTT) were purchased from companies with good quality and reputation.
HT-29 colon cancer cell lines (Shanghai Institute of Biochemistry and Cell Biology), NCM-460 human normal colonic epithelial cell line (Guangzhou Jini Ou Biotechnology Co., Ltd.), Annexin V-FITC/PI Apoptosis Detection Kit (Beijing Kang Century Biotech Co., Ltd.), DNA content detection kit (Nanjing KGI Biological Technology Development Co., Ltd.) and Dil red fluorescent membrane probe (Bi Yun Tian Biotechnology Co., Ltd), etc. were used. The entire research was conducted at Henan University of Traditional Chinese Medicine in the period from December, 2017-January, 2019.
Preparation of WGA modified matrine nanoparticles (WGA-MT-NPs): WGA-MT-NPs were prepared using the double emulsification-solvent evaporation method followed by the carbodiimide method as previously described15,16. Briefly, after the hot water solution of MT was dropped into the acetone solution of PLGA under magnetic stirring, the resulting emulsion was injected rapidly into the water solution of PVA under stirring condition. Subsequently, the resulting solution wascontinuously stirred to remove acetone and filtered to obtain MT nanoparticles (MT-NPs).After PBS solution of EDAC and NHS added into the MT-NPs solution to activate for 4 hrs, EDC and NHS were removed by centrifugation. Then PBS solution of WGA was added to modify for 14 hrs, WGA-MT-NPs were obtained by centrifugation and wash to remove free lectin and lyophilized.
Characterization: HPLC and nano-laser particle size analyzer were used to measure encapsulation efficiency, average particle size and potential value, respectively. The folin-phenol method was used to measure the unbound WGA lectin as per manufacturer’s instructions (Sigma):
![]()
And transmission electron microscopy (TEM) was used to determine the image of WGA-MT-NPs.
Interactions between WGA-MT-NPS and HT-29 cells as well as NCM460 cells: The affinity of nanoparticles towards colon cells was measured using fluorescein isothiocyanate (FITC) labeled WGA, according to the method reported by Tammam et al.17 with modification. Briefly, after HT-29 cells or NCM4460 cells (1×105 mL–1) were seeded and cultured for 24 hrs, WGA-MT-NPs solution (200 μg mL–1 of MT) was added. Cells were then incubated for 25 min, washed with PBS, stabilized for 10 min with 4% paraformaldehyde at room temperature, washed again with PBS and immediately added the plasma membrane fluorescent dye Dil 500 μL pre-balanced at room temperature to incubate for 15 min. Discarding Dil, cells were washed and imaged with confocal microscopy.
Effect on HT-29 human colon cancer cell proliferation: HT-29 cells (1×105 mL–1) were seeded and cultured in a 96-well plate for 24 hrs. WGA-MT-NPs and MT-NPs with MT concentrations of 25, 50, 100, 200 and 400 μg mL–1 were separately added. After culturing for 24, 48 and 72 hrs, the MTT method was used to detect cell proliferation as described previously18.
Effects on HT-29 human colon cancer cell cycle and apoptosis: HT-29 cells (2×105 mL–1) were seeded and cultured for 24 hrs. WGA-MT-NPs or MT-NPs were added at final concentration 100, 150 and 200 μg mL–1 of MT. After 48 hrs, cells were digested with 0.25% trypsin, washed with 4°C pre-cooled PBS buffer. Then, cells were treated according to manufacturer’s instructions for detecting cell cycle and cell apoptosis by flow cytometry. The apoptotic rate was thesumofthe percentageof earlyandlate apoptosis.
Statistical analysis: Statistical analysis was performed by SPSS19.0 software using the one way ANOVA, p<0.05 and <0.01 represented significant and extremely significant differences, respectively.
RESULTS
MT encapsulation efficiency and WGA modification rate: The encapsulation efficiency of WGA-MT-NPs was 89.99±5.08% according to the result of HPLC (Fig. 1a). And, according to the standard curve of WGA: y = 4.789×10–4 x+0.7107, R = 0.9996 (Fig. 1b), the modifying efficiency of WGA for MT-NPs was determined as 87.57±8.61%.
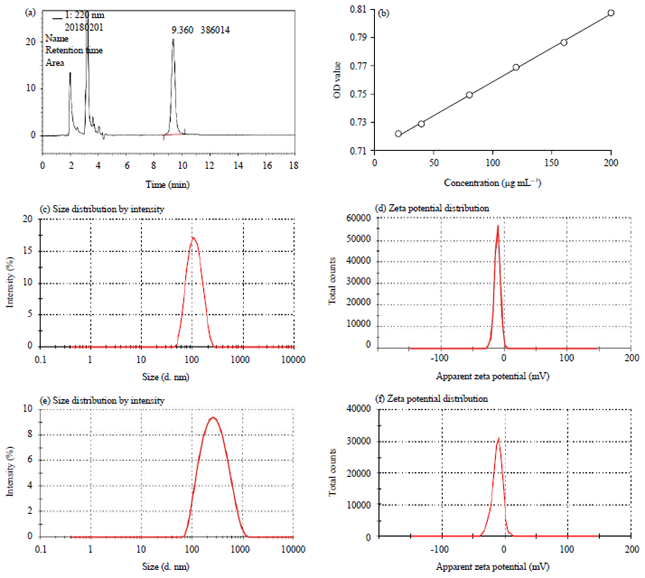 | |
| Fig. 1(a-f): | HPLC chromatogram, (a, b) WGA-MT-NPs and standard curve of WGA, (c, d) MT-NPs size distribution and zeta potential and (e,f) WGA-MT-NPs size distribution and zeta potential |
 | |
| Fig. 2(a-b): | TEM images of (a) MT-NPs and (b) WGA-MT-NPs |
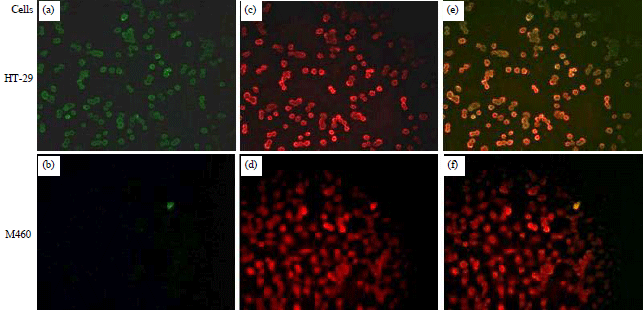 | |
| Fig. 3(a-f): | Confocal images of WGA-MT-NPs affinity towards HT-29 cancer cells and NCM460 normal cells |
| a-b: FITC-WAG, c-d: Dil red fluorescent membrane, e-f: Merged channels | |
Size distribution, zeta potential and morphology: As shown in Fig. 1c-f, the sizes and potential of MT-NPs and WGA-MT-NPs were 102.93±5.53 nm, -12.73±0.68 mV and 220.10±13.80 nm, -10.50±0.70 mV, respectively. TEM images further revealed their morphology. The MT-NPs particles were a distinct spherical or near-spherical shape (Fig. 2a). However, WGA-MT-NPs (Fig. 2b) were significantly changed after modified by WGA, became irregular with some viscous material attached to MT-NPs core.
Higher affinity between WGA-MT-NPs and HT-29 cells: The Fig. 3 reveals the affinity of WGA-MT-NPs. After the FITC-WGA conjugates were used to prepare WGA-MT-NPs, HT-29 cells had obvious green fluorescence, while NCM460 normal colon cells only had very little green fluorescence. The overlap of the FITC green fluorescence and the red fluorescence of Dil, a lipophilicmembranestain, in HT-29 cells had evident co-localization, showing that WGA-MT-NPs had a significant stronger affinity towards HT-29 cells membrane compared with NCM460 cells.
Effects of WGA-MT-NPs on HT-29 cells proliferation: The cell inhibition in vitro is shown in Fig. 4. Within the concentration at 25-100 μg mL–1 of MT, the inhibition rate of WGA-MT-NPs on HT-29 cells were 3-6 times compared to that of MT-NPs. Withan increaseof MT concentration to 200 μg mL–1, WGA-MT-NPs inhibition rate had almost reached its maximum, which was up to 81.6% at 48 hrs.
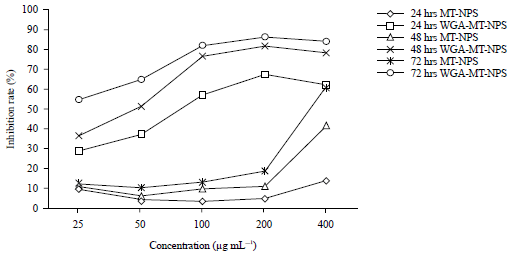 | |
| Fig. 4: | WGA-MT-NPs enhanced the growth inhibition of MT-NPs in HT-29 cells |
| The effect on HT-29 human colon cancer cell proliferation was detected by MTT method | |
Whereas the inhibition rate at 48 hrs was only 41.6% even if the MT-NPs concentration reached 400 μg mL–1. The result suggested that with the help of the WGA, the effect of WGA-MT-NPs at low concentrations nearly equal at high concentrations of MT-NPs.
Effect of WGA-MT-NPs and MT-NPs on HT-29 cell cycle and apoptosis: Results in Fig. 5 show the effects of the nanoparticles on the HT-29 cell cycle. Compared with the control group, treatment with 100, 150 and 200 μg mL–1 both MT-NPs and WGA-MT-NPs were able to significantly increase the cells at the G1 phase and decrease the cells at S phase (p<0.05 and <0.01) with a dose-dependent effect. However, no statistically significant differences were observed between the WGA-MT-NPs group and the MT-NPs group.
As shown in Fig. 6, both MT-NPs and WGA-MT-NPs can induce HT-29 cell apoptosis. Compared with the control group, the 100, 150 and 200 μg mL–1 of MT-NPs groups caused significant apoptosis with an apoptotic rate of 19.06, 22.73 and 30.20%, respectively (p<0.01). The corresponding WGA-MT-NP caused more severe apoptosis with an apoptotic rate of 26.82, 38.30 and 55.70%, respectively. The significant differences were found between the WGA-MT-NPs group and the same concentration MT-NPs group at 150 and 200 μg mL–1.
DISCUSSION
The modification of WGA was confirmed by the increase of the average diameter of nanoparticles (Fig. 1c, e) and the change of shape from near-spherical to non-uniform (Fig. 2a, b), which may be due to WGA grafting on the surface of MT-NPs, its hydrophilic properties can make the protein’s macromolecular chains extend to the water phase and cause the hydrated radius of the nanoparticles to increase. Increased size after WGA modification in the present study is in agreement with Liu et al.19 report.
WGA-MT-NPs had better affinity on HT-29 cells, rather normal colon cells (Fig. 3a, b). It was reported that the cell surface of colon cancer epithelial cells, such as Caco-2 and HT-29, had a large number of glycoproteins, while there was an apparent lack of these glycoproteins on normal colon cells20,21, thus CRC could be successfully distinguished from normal cells by targeting this glycosylation with affinity capillary electrophoresis22. Our result also accounts for targeting glycoproteins, the binding sites of WGA, could recognize colon cancer cells.
Protein glycosylation is the attachment of carbohydrate to the amino acid residue of the protein backbone23,24. O-GlcNAcylation in which N-acetyl glucosamine (GlcNAc) is added to the serine or threonine residue of protein, N-linked and O-linked modifications are the commonly occurring types of glycan modifications of protein25. Altered mucin expression in conjunction with differential glycosylation has been strongly associated with malignant pathologies of colon26. The aberrant glycosylation, such as an increase in core fucosylation and GlcNAc branching on N-glycans, alteration of O-glycans and O-GlcNAcylation, have been reported in CRC20. Also, WGA can bind GlcNAc-related oligosaccharides and has applications as a commercial reagent to detect glycans containing GlcNAc modified residues27 and WGA was used to determine the mucin secreted by colon carcinoma Caco-2 and mucus-producing HT29-MTX-E12 cell monolayers28. And, a lectin-based reverse-phase protein microarray method had been developed for screening the glycosylation pattern of receptors isolated from thepatientsofcoloniccarcinoma29.
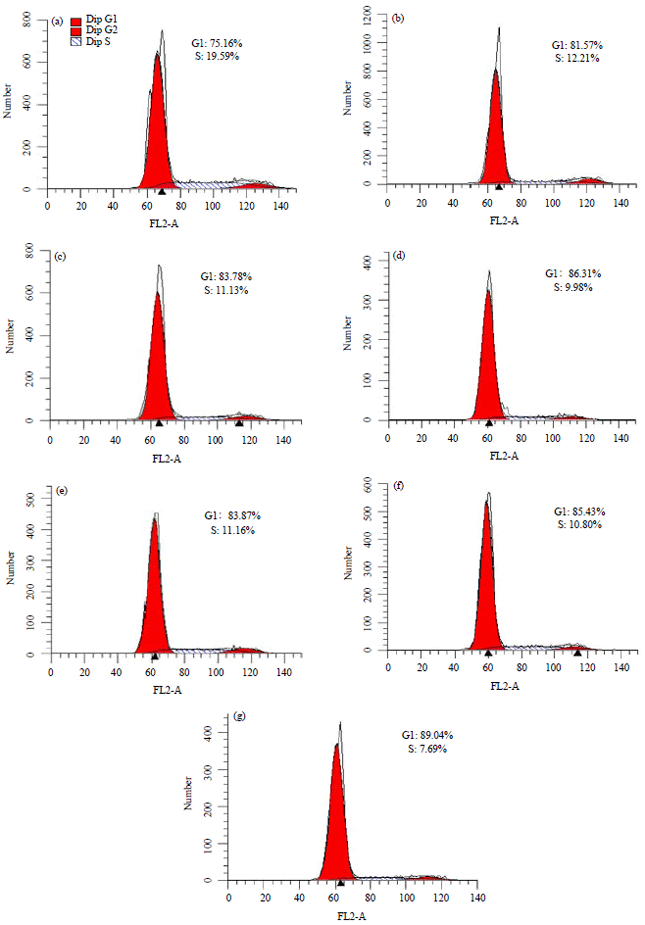 | |
| Fig. 5(a-g): | Effect on HT-29 human colon cancer cell cycle |
a: Control, b-d: 100, 150 and 200 μg mL–1 of MT-NPs, e-g: 100, 150 and 200 μg mL–1 of WGA-MT-NPs | |
 | |
| Fig. 6: | Effect on HT-29 colon cancer cell apoptosis |
Percent of apoptosis cells included the early and late apoptosis cells. #p<0.05, ##p<0.01 vs the control group, **p<0.01 vs the same concentration of MT-NPs group | |
All of these studies may further explain the mechanisms of WGA targeting CRC. These mechanisms may contribute to the targeting function of WGA-MT-NPs.
Modification with WGA significantly increased the growth inhibitory activity of the corresponding MT-NPs group at the same concentration (Fig. 4). In accordance with our results, Pandey et al.30 also reported the WGA-conjugated polylactic-co-glycolic acid nanoparticles of 5-fluorouracil could significantly inhibit the growth of colon cancer cells compared to the non-conjugated NPs and pure drug. Also, recent study has shown that glycosylation and glycan interactions affect clathrin-independent endocytosis31. And, the lipid nanoparticles of bufalin with WGA could also increase bufalin's uptake by colon cancer CaCo-2 cells in vitro32. These results suggested that the improved inhibition effects of MT-NPs by WGA modification may be related to the uptake.
Both WGA-MT-NPs and MT-NPs were able to hinder DNA synthesis and induce apoptosis in HT-29 cells (Fig. 5 and 6). However, compared with the same concentration MT-NPs, WGA-MT-NPs significantly enhanced the apoptosis of HT-29 cells (Fig. 6). This may be because MT can inhibit DNA synthesis and induce apoptosis in HT-29 cells3,33 and with the help of WGA, WGA-MT-NPs enabled more cells to apoptosis. In similar to our results, WGA-EF-NPs (5-fluorouracil and epigallocatechin-3-gallate co-loaded in wheat germ agglutinin-conjugated nanoparticles) exhibited superior anti-tumor activity and pro-apoptotic efficacy compared to the drugs nanoparticles without WGA decoration34. Therefore, the significantly increased apoptosis in HT-29 cells may play a role in WGA-MT-NPs significantly enhancing the antitumor effect of MT-NPs.
CONCLUSION
WGA-MT-NPs significantly increased the antitumor effect of MT-NPs via stronger affinity towards HT-29 cells, higher promotion of apoptosis and maintenance of inhibiting proliferation. Thus, WGA-MT-NPs may provide a promising new strategy for anti-colon cancer. The potential of WGA-MT-NPs for the effective treatment of CRC is worthy of further study in vivo.
SIGNIFICANCE STATEMENT
This study discovered the targeting function and the increased apoptosis effect of WGA modifying matrine nanoparticles, which can be beneficial for improving the anti-colon cancer efficiency of matrine. This study will help the researchers to uncover the critical areas ofcoloncancer treatment with MT’s nano-pharmaceutics that many researchers were not able to explore. Thus a new therapy strategy of MT on colon cancer may be arrived at.
ACKNOWLEDGMENTS
This study was supposed by the Scientific and Technological Projects of the Science and Technology Department of Henan Province (NO: 132102310267). We would like to give our thanks to Professor Sugai Yin in Henan University of TCM and Dr. Jie Zhou in Zhengzhou University for their guidance in technologies.
REFERENCES
- Bray, F., J. Ferlay, I. Soerjomataram, R.L. Siegel, L.A. Torre and A. Jemal, 2018. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: Cancer J. Clin., 68: 394-424.
CrossRefDirect Link - Zhang, S., B. Cheng, H. Li, W. Xu and B. Zhai et al., 2014. Matrine inhibits proliferation and induces apoptosis of human colon cancer LoVo cells by inactivating Akt pathway. Mol. Biol. Rep., 41: 2101-2108.
CrossRefDirect Link - Chang, C., S.P. Liu, C.H. Fang, R.S. He, Z. Wang, Y.Q. Zhu and S.W. Jiang, 2013. Effects of matrine on the proliferation of HT29 human colon cancer cells and its antitumor mechanism. Oncol. Lett., 6: 699-704.
CrossRefDirect Link - Duan, L., L. Deng, D. Wang, S. Ma, C. Li and D. Zhao, 2017. Treatment mechanism of matrine in combination with irinotecan for colon cancer. Oncol. Lett., 14: 2300-2304.
CrossRefDirect Link - Ren, H., S. Zhang, H. Ma, Y. Wang, D. Liu, X. Wang and Z. Wang, 2014. Matrine reduces the proliferation and invasion of colorectal cancer cells via reducing the activity of p38 signaling pathway. Acta Biochim. Biophys. Sin., 46: 1049-1055.
CrossRefDirect Link - Ruan, J., J. Liu, D. Zhu, T. Gong, F. Yang, X. Hao and Z. Zhang, 2010. Preparation and evaluation of self-nanoemulsified drug delivery systems (SNEDDSs) of matrine based on drug–phospholipid complex technique. Int. J. Pharm., 386: 282-290.
CrossRefDirect Link - Yang, Z., S. Gao, T. Yin, K.H. Kulkarni, Y. Teng, M. You and M. Hu, 2010. Biopharmaceutical and pharmacokinetic characterization of matrine as determined by a sensitive and robust UPLC–MS/MS method. J. Pharm. Biomed. Anal., 51: 1120-1127.
CrossRefDirect Link - Chen, X., J. Yan, S. Yu and P. Wang, 2018. Formulation and in vitro release kinetics of mucoadhesive blend gels containing matrine for buccal administration. AAPS Pharm. Sci. Tech., 19: 470-480.
CrossRefDirect Link - Venkitachalam, S., L. Revoredo, V. Varadan, R.E. Fecteau and L. Ravi et al., 2016. Biochemical and functional characterization of glycosylation-associated mutational landscapes in colon cancer. Sci Rep., Vol. 6.
CrossRefDirect Link - Kuo, J.C.H., A.E.K. Ibrahim, S. Dawson, D. Parashar and W.J. Howat et al., 2016. Detection of colorectal dysplasia using fluorescently labelled lectins. Sci Rep., Vol. 6.
CrossRefDirect Link - Bruschi, M.L., 2019. Lectins and nanostructured drug delivery systems. Curr. Drug Deliv., 16: 268-269.
CrossRefDirect Link - Devi, R.V. and M.R. Basil-Rose, 2018. Lectins as ligands for directing nanostructured systems. Curr. Drug Deliv., 15: 448-452.
CrossRefDirect Link - Glavas-Dodov, M., B. Steffansen, M.S. Crcarevska, N. Geskovski, S. Dimchevska, S. Kuzmanovska and K. Goracinova, 2013. Wheat germ agglutinin-functionalised crosslinked polyelectrolyte microparticles for local colon delivery of 5-FU: In vitro efficacy and in vivogastrointestinal distribution. J. Microencapsulat., 30: 643-656.
CrossRefDirect Link - Murata, M., T. Yonamine, S. Tanaka, K. Tahara, Y. Tozuka and H. Takeuchi, 2013. Surface modification of liposomes using polymer-wheat germ agglutinin conjugates to improve the absorption of peptide drugs by pulmonary administration. J. Pharm. Sci., 102: 1281-1289.
CrossRefDirect Link - Makhlof, A., S. Fujimoto, Y. Tozuka and H. Takeuchi, 2011. In vitro and in vivo evaluation of WGA–carbopol modified liposomes as carriers for oral peptide delivery. Eur. J. Pharm. Biopharm., 77: 216-224.
CrossRefDirect Link - Noviendri, D., I. Jaswir, M. Taher, F. Mohamed and H.M. Salleh et al., 2016. Fabrication of fucoxanthin-loaded microsphere (F-LM) by two steps double-emulsion solvent evaporation method and characterization of fucoxanthin before and after microencapsulation. J. Oleo Sci., 65: 641-653.
CrossRefDirect Link - Tammam, S.N., H.M.E. Azzazy and A. Lamprecht, 2015. A high throughput method for quantification of cell surface bound and internalized chitosan nanoparticles. Int. J. Biol. Macromol., 81: 858-866.
CrossRefDirect Link - Li, H.X., S.Y. Han, X. Ma, K. Zhang, L. Wang, Z.Z. Ma and P.F. Tu, 2012. The saponin of red ginseng protects the cardiac myocytes against ischemic injury in vitro and in vivo. Phytomedicine, 19: 477-483.
CrossRefPubMedDirect Link - Liu, Y., J. Liu, J. Liang, M. Zhang and Z. Li et al., 2017. Mucosal transfer of wheat germ agglutinin modified lipid–polymer hybrid nanoparticles for oral delivery of oridonin. Nanomed. Nanotechnol. Biol. Med., 13: 2219-2229.
CrossRefDirect Link - Biwi, J., C. Clarisse, C. Biot, R.P. Kozak and K. Madunic et al., 2019. OGT controls the expression and the glycosylation of E-cadherin and affects glycosphingolipid structures in human colon cell lines. Proteomics, Vol. 19.
CrossRefDirect Link - Wang, C., P.C. Ho and L.Y. Lim, 2010. Wheat germ agglutinin-conjugated PLGA nanoparticles for enhanced intracellular delivery of paclitaxel to colon cancer cells. Int. J. Pharm., 400: 201-210.
CrossRefDirect Link - Yamamoto, T., K. Sato, S. Wakahara, K. Mitamura and A. Taga, 2020. A method for detecting tumor cells derived from colorectal cancer by targeting cell surface glycosylation with affinity capillary electrophoresis. J. Pharm. Biomed. Anal., Vol. 182.
CrossRefDirect Link - Venturi, G., I.G. Ferreira, M. Pucci, M. Ferracin, N. Malagolini, M. Chiricolo and F. Dall’Olio, 2019. Impact of sialyltransferase ST6GAL1 overexpression on different colon cancer cell types. Glycobiology, 29: 684-695.
CrossRefDirect Link - Nagae, M., Y. Kizuka, E. Mihara, Y. Kitago and S. Hanashima et al., 2018. Structure and mechanism of cancer-associated N-acetylglucosaminyltransferase-V. Nat. Commun., Vol. 9.
CrossRefDirect Link - Barkeer, S., S. Chugh, S.K. Batra and M.P. Ponnusamy, 2018. Glycosylation of cancer stem cells: Function in stemness, tumorigenesis and metastasis. Neoplasia, 20: 813-825.
CrossRefDirect Link - Pothuraju, R., S.R. Krishn, S.K. Gautam, P. Pai and K. Ganguly et al., 2020. Mechanistic and functional shades of mucins and associated glycans in colon cancer. Cancers, Vol. 12.
CrossRefDirect Link - Biari, K.E., Á. Gaudioso, M.C. Fernández-Alonso, J. Jiménez-Barbero and F.J. Cañada, 2019. Peptidoglycan recognition by wheat germ agglutinin. A view by NMR. Nat. Prod. Commun., Vol. 14.
CrossRefDirect Link - Limage, R., E. Tako, N. Kolba, Z. Guo, A. García‐Rodríguez, C.N.H. Marques and G.J. Mahler, 2020. TiO2 nanoparticles and commensal bacteria alter mucus layer thickness and composition in a gastrointestinal tract model. Small, Vol. 16.
CrossRefDirect Link - Robajac, D., M. Križáková, R. Masnikosa, G. Miljuš, M. Šunderić, O. Nedić and J. Katrlík, 2020. Sensitive glycoprofiling of insulin-like growth factor receptors isolated from colon tissue of patients with colorectal carcinoma using lectin-based protein microarray. Int. J. Biol. Macromol., 144: 932-937.
CrossRefDirect Link - Pandey, A.N., K. Rajpoot and S.K.J.K. Jain, 2020. Using 5-fluorouracil-encored plga nanoparticles for the treatment of colorectal cancer: The in vitro characterization and cytotoxicity studies. Nanomed. J., 7: 211-224.
CrossRefDirect Link - Mathew, M.P. and J.G. Donaldson, 2019. Glycosylation and glycan interactions can serve as extracellular machinery facilitating clathrin‐independent endocytosis. Traffic, 20: 295-300.
CrossRefDirect Link - Liu, Y., P. Wang, C. Sun, N. Feng and W. Zhou et al., 2010. Wheat germ agglutinin-grafted lipid nanoparticles: Preparation and in vitro evaluation of the association with Caco-2 monolayers. Int. J. Pharm., 397: 155-163.
CrossRefDirect Link - Gu, Y.Y., M.H. Chen, B.H. May, X.Z. Liao and J.H. Liu et al., 2018. Matrine induces apoptosis in multiple colorectal cancer cell lines in vitro and inhibits tumour growth with minimum side effects in vivo via Bcl-2 and caspase-3. Phytomedicine, 51: 214-225.
CrossRefDirect Link - Wang, R., J. Huang, J. Chen, M. Yang and H. Wang et al., 2019. Enhanced anti-colon cancer efficacy of 5-fluorouracil by epigallocatechin-3- gallate co-loaded in wheat germ agglutinin-conjugated nanoparticles. Nanomed. Nanotechnol. Biol. Med., Vol. 21.
CrossRefDirect Link