
Gomaa Mostafa-Hedeab
Department of Pharmacology, Medical College, Jouf University, Kingdom of Saudi Arabia
LiveDNA: 966.33513
Manal Ewaiss Hassan
Department of Pathology, Medical College, Jouf University, Kingdom of Saudi Arabia
Dina Sabry
Department of Biochemistry and Molecular Biology, Faculty of Medicine, Cairo University, Egypt
Rehab Ibrahim Ali
Department of Pathology, Medical College, Jouf University, Kingdom of Saudi Arabia
International Journal of Pharmacology
Year: 2020 | Volume: 16 | Issue: 6 | Page No.: 437-446







ABSTRACT
Background and Objective: Diabetes Mellitus (DM) type 1 is characterized by a loss of beta cells; its current therapy depends on the administration of insulin with no available curable therapy till now. We evaluated the effect of Mesenchymal Stem Cells-Derived Exosomes (MSCs-Exs) in the treatment of induced-type I DM in rats; additionally, were compared the effects of the MSCs-Exs versus the undifferentiated MSCs (Un- MSCs) and Vitamin d (Vit. D). Materials and Methods: Seventy-five rats, six weeks old, weighing 150-170 gm were equally divided into five groups, fifteen rats each: group1: healthy control group, group II: diabetic non-treated group, group III: Un-MSCs treated group, group IV: MSCs-Exs treated group and group V: Vit. D treated group. Diabetes was induced by streptozotocin (STZ) injection intraperitoneal (IP) in groups II, III, IV and V. Estimation of serum insulin and glucose levels were done after one, two and three months. At the end of the experiment, animals were sacrificed and pancreatic tissues were obtained for insulin, Smad2, Smad3, Pancreatic And Duodenal Homeobox 1 (PDX1), Paired Box 4 (PAX4) and neuro D genes expression and histopathological examinations. Results: Un-MSCs, Vit. D or MSCs-Exs resulted in lower serum glucose and higher serum insulin with significantly higher Insulin, Smad2, Smad3, PDX1, PAX4 and neuro D gene expressions compared to the diabetic non-treated group. Results were superior in the case of MSCs-Exs compared to either Un-MSCs or Vit. D. Conclusion: MSCs-Exs are found to be better than Un-MSCs or Vit. D in the treatment of experimental type-I DM. MSCs-derived exosomes may represent a novel cell-based treatment agents of type-1 DM treatment.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Gomaa Mostafa-Hedeab, Manal Ewaiss Hassan, Dina Sabry and Rehab Ibrahim Ali, 2020. Anti-diabetic Therapeutic Efficacy of Mesenchymal Stem Cells-derived Exosomes. International Journal of Pharmacology, 16: 437-446.
DOI: 10.3923/ijp.2020.437.446
URL: https://scialert.net/abstract/?doi=ijp.2020.437.446
DOI: 10.3923/ijp.2020.437.446
URL: https://scialert.net/abstract/?doi=ijp.2020.437.446
INTRODUCTION
Diabetes mellitus is worldwide diseases, over 415 million people suffer from diabetes and the incidence is expected to reach1 to 642 million by 2040.
Diabetes is a chronic metabolic disorder characterized by dysfunction of the insulin-secreting pancreatic beta cells. Type 1 DM is characterized by irreversible beta cell destruction occurs and abolishing insulin secretion2 thus necessitates the life-long dependence of insulin injection to regulate blood glucose level; thus type 1 diabetic patients are strictly dependent on daily insulin injection to regulate blood glucose level3.
MSCs are present in many tissues as bone marrow, synovium and adipose tissues4, representing an important source for regenerative medicine that can differentiate into different types of cells with its function5. Previous reports of MSCs showed its immunomodulatory actions6. It acts as an immune and inflammatory system through the suppression of natural killer, T-lymphocytes and dendritic cells7.
MSCs can be segregated from bone marrow which represents the first source reported8, then adipose tissues were discovered as an alternative rich source9 and subsequent MSCs have been successfully segregated synovium10.
MSCs are potent tissue regenerators11 that can differentiate to several types of cells like epithelial cells, stromal cells and hematopoietic cells12.
Exosomes are secreted vesicles with very small structures measured in nanoscale, can transport different types of proteins and lipid components to neighboring cells13. MSCs-Ex therapeutic efficacy is thought duet to its pleiotropic effects and the interacting factors and vesicles of its structure14, its mechanism of action are thought to be through its inward budding to multivesicular bodies15. which is then going to the inside cell via its fusion to the cell membrane or taken up by cells16.
Exosome was previously discovered in immature red blood cells17. Recently, it has been shown that SC-derived exosomes enhance cell proliferation and maintenance of its survival by the relocation of different types of proteins and non-coding types of RNAs18. Exosomes are considered as the active ingredient of MSCs-based therapy19.
Vit. D deficiency is shown to be linked to various types of diseases including DM20. Active Vit. D, (1,25OH2D) can induce insulin synthesis through binding to the Vit. D response element in the human insulin gene promoter in pancreatic B cell21. Vit. D also improves pancreatic Beta-cell function and insulin resistance through its role in ca homeostasis22. Vit. D improves insulin resistance in type-2 diabetic rats23, also Vit. D deficiency was found to be more common in diabetic patients24. Moreover, Zeitz et al.25, showed impaired insulin secretion was reported in mice with defective Vit. D receptor.
The discovery of a curable treatment for type-1 DM is an important target and still receiving updates elsewhere; Vit. D, MSC and MSCs-Ex represent promising agents that are widely used in experimental work, that could be hopefully used in the treatment of DM.
The current study was designed to evaluate the effect of MSCs-Exs in the treatment of induced-type I diabetes mellitus (DM) in rats; additionally, we compared the effects of the MSCs-Exs versus the MSCs and Vit. D in the regeneration of β islets of Langerhans in experimental Type-I DM.
MATERIALS AND METHODS
Study area: This study was carried out at Molecular Biology Lab. Biochemistry Department, Medical College, Cairo University, Egypt from January 2018 to March 2019.
Experimental animals and induction of diabetes: Seventy-five male albino rats, six weeks old, weighing 150-170 gm were included in the current experiment. Rats were maintained on stainless steel cages in an animal house with an air conditioner with 12/12-hour daylight/dark with free access to food and water; they kept for two weeks before starting the experiment for acclimatization. Rats then divided into five groups, fifteen rats each as follows: group1: healthy control group, group II: diabetic non-treated group, group III: Un-MSCs treated group (3×106 cells per rat) intravenously26, group IV: MSCs-Exs treated group (0.4 μg mL–1, 100 μL) IV via tail vein, twice-weekly27 and group V: Vit. D treated group (0.1 μg/kg/day) orally daily28.
Induction of diabetes: Diabetes was induced by injection of streptozotocin in a dose of 55 mg kg–1 intravenous (once), control rats were injected with vehicle. Confirmation of the occurrence of diabetes was done by regular measuring of blood glucose level; the diabetic rat was considered when blood glucose exceeds 300 mg dL–1.
Bone marrow stem cells isolation and characterization: The process of BM isolation is done following the method described by Abd Elaziz et al.9. In brief, phosphate-buffered saline (PBS) was used to flush BM cells' tibia and fibula rats' bones. The results of BM cells flushed were then centrifuged then the upper layer was discarded. Isolated BM-MSCs were cultured, propagated and supplemented with 10% fetal bovine serum (FBS), 0.5% penicillin/ streptomycin and incubated at 37°C and 5% CO2 until reaching 80-90% confluence26.
The isolated MSCs were identified by identification of its morphology character, adherence, surface markers, Cell surface antigen expression of BM-MSCs was analyzed by flow cytometry in which cells were incubated with fluorescein-isothiocyanate-conjugated (anti-CD45) or phycoerythrin cyanine-5-conjugated (anti-CD90) or rat immunoglobulin G (IgG) isotype control antibodies for 30 min at 4°C in PBS.
Exosomes isolation: The release of MSCs derived exosomes (diameter <100 nm) is a cellular process which is a method of communication that occurs in all living cells. To get rid of cells and debris; MSCs' conditioning media was subjected to centrifugation at 3000 g for 15 min at room temperature. The cell-free supernatant was recentrifuged at 100,000×g (ultracentrifuge of Beckman Coulter Optima L 90 K) for 1 h at 4°C to precipitate Exs29. The protein content was quantified by the Bradford method (BioRad, Hercules, CA). The pellet was resuspended in 100 μL of glutaraldehyde for TEM analysis and characterization, or 600 μL of Lysis/Binding Buffer (Zymo Research Quick-gRNA™ MiniPrep kit, USA) and stored at -80°C.
Exosomes characterization: Exosomes were diluted using 145 μL Phosphate Buffer Saline (PBS) containing 0.2% paraformaldehyde (w/v). Ten microliter was administered to a formvar-carbon-coated 300 mesh grid (Electron Microscopy Sciences, Hatfield, USA) for 7 min, 1.75% uranyl acetate (w/v) was used for staining. Samples were left to dry at room temperature for 2 h then imaged by transmission electron microscopy (TEM) (CM-10, Philips, Eindhoven, The Netherlands) at 100 kV afterward30, Fig. 1.
Biochemical investigations: Serum glucose and insulin levels from all studied groups were assessed according to the time chart of the study and according to kits instruction (Cell Biolabs, Inc. San Diego, CA 92126) and (Mercodia AB, Sweden, No: 34-0130) respectively.
Animals scarification: Retro-orbital blood samples were withdrawn from each animal at the end of the 1st, 2nd and 3rd months of the experimental period, for insulin and glucose blood level estimations31.
 | |
| Fig. 1: | TEM of EXs; they were spheroid (their size is less than 100 nm) and have a well-defined membrane with heterogeneous contents |
| Table 1: | Primers sequence of all the studied genes |
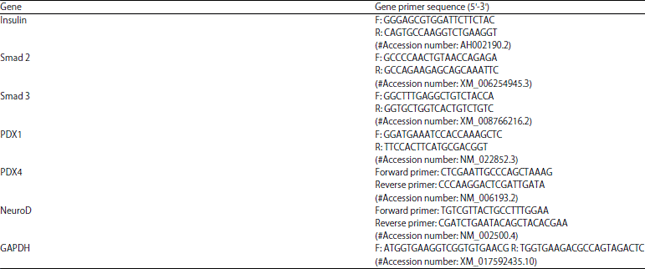 | |
| F: Forward primer and R: Reverse primer | |
At the end of the experiment, rats were sacrificed under general anesthesia. The pancreas of each rat was taken, half for histopathological examination and the other half for gene expression assay. The protocol of this study was approved by the ethical committee, college of medicine, Cairo University under approval number (CU-III-F-64-18).
RNA extraction: RNA was extracted from purified EXos by Trizol Isolation Kit (Trizol Quick-gRNA™ kit, USA), following the manufacturer’s instruction. Nanodrop ND-1000 (Thermo Fisher Scientific) were used to measure RNA concentration.
mRNA expression analysis by qRT-PCR: 50 ng of total RNA was used for reversal-transcribed to cDNA with TaqMan® Reverse Transcription Kit (Applied Biosystem). cDNA was pre-amplified with Megaplex™ RT Primers, (Thermo Fisher Scientific) using a Biometra Thermal Cycler (Thermo Fisher Scientific). The qRT-PCR reaction mix was composed of 10 ng of cDNA, 50 nM miScript Universal Primer (Qiagen), 20 nM mRNA-specific primer, 10 μL QuantiTect SYBR Green PCR Master Mix (Qiagen) and nuclease-free water (Qiagen) to reach a final reaction volume of 20 μL. The expression of mRNAs of studied genes (Insulin, Smad2, Smad3, PDX1, PAX4 and neuro D) in all groups were assessed (Table 1). Each sample was run in duplicate to ensure quantitation results and negative control to exclude contamination. GAPDH was used as an endogenous control to normalize mRNA results by 2–ct method. The Real-Time Thermal Cycler StepOne plus (Applied biosystem, Foster City, USA) was used for analysis.
Histopathological examination: At room temperature, pancreatic tissue from each animal was obtained and fixed in 10% formalin solution for 72 hours and then embedded in paraffin blocks. From each block, histological sections of 4 μm thickness were prepared, mounted to glass slides, stained by hematoxylin and eosin (H and E).
Slides were examined by light microscopy first by scanning and low power to assess the adequacy of specimens and take initial impression about architecture and ratio between exocrine and endocrine compartments, number and size of islets, presence of leukocyte infiltrate, vascular changes, cell density within the islets and any deposits. High power was used to assess cellular changes within the islets32.
Statistical analysis: Our results were collected, tabulated, analyzed using SPSS 17 for Windows. Data were expressed as Mean±SEM. Groups were compared using the One-Way ANOVA test, followed by LSD as a post hoc test, paired T-test was used when needed. The result considered significant if P value <0.05.
RESULTS
This study was carried on 75 rats divided into 5 groups, 15 rats each as follows: Control group, STZ (diabetic non-treated group), Un-MSC-treated group, MSCs-Exs and Vit. D-treated groups.
Administration of STZ significantly decrease serum insulin and increase serum glucose levels compared to the control group.
Un-MSC, MSCs-Exs groups and Vit. D significantly increase serum insulin and significantly decreased serum glucose levels compared to the STZ group. The EX-MSC-treated group showed a significant increase in serum insulin and a significant decrease in glucose levels compared to the MSC-treated group or Vit. D-treated group (Fig. 2, 3).
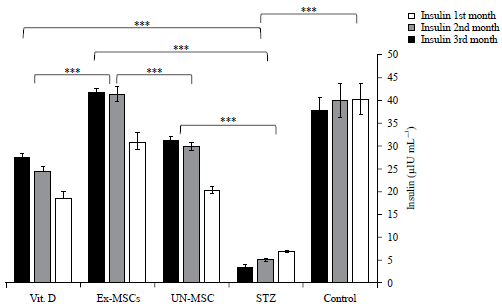 | |
| Fig. 2: | Serum insulin assay among studied groups over three months |
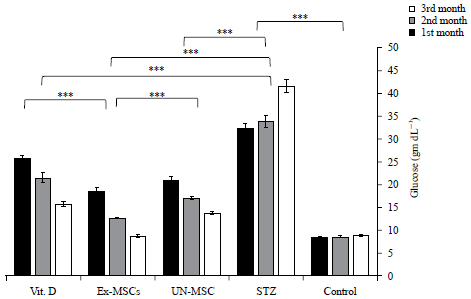 | |
| Fig. 3: | Glucose level among studied groups over three months |
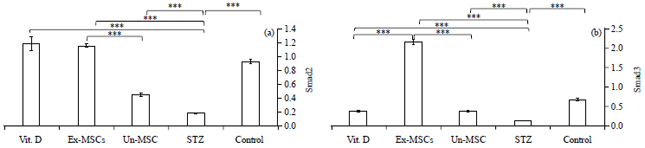 | |
| Fig. 4(a-b): | Quantitative pancreatic (a) Smad2 and (b) Smad3 gene expression of among studied groups |
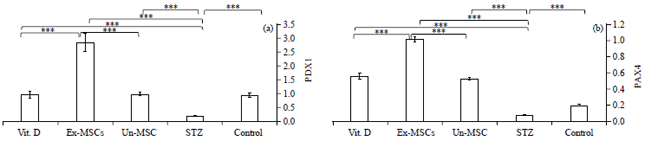 | |
| Fig. 5(a-b): | Quantitative pancreatic (a) PDX1 and (b) PAX4 gene expression among studied groups |
Quantitative estimation of pancreatic Smad2 and Smad3 gene expression levels was done using Real-Time PCR (Fig. 4). Smad2, 3 significantly decreased in the STZ-treated group compared to the untreated group. Un-MSC, Vit. D and Ex-MSC treated groups significantly upregulated pancreatic Smad2 gene expression (Fig. 4a). Ex-MSC treated group showed a significant increase in Smad3 expression compared to either Vit. D or Un-MSC-treated group (Fig. 4b).
PDX1 and PAX4 gene expression significantly decreased in the STZ group as compared to the control untreated group (Fig. 5). Un-MSC treated and EX-MSC-treated groups significantly increase PDX1 and PAX4 compared to the STZ group. While Vit. D treated group significantly upregulated only PAX4 compared to the SZT-treated group. EX-MSC-treated group significantly increase PDX1 and PAX4 gene expression levels compared to either Vit. D or Un-MSC treated groups (Fig. 5a, b).
Neuro D gene expression level was significantly decreased in the STZ-treated group compared to the untreated group; while Un-MSC, Vit. D and Ex-MSC treated groups significantly upregulated Neuro D gene expression compared to the STZ-treated group.
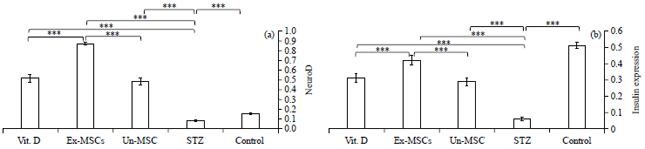 | |
| Fig. 6(a-b): | Quantitative pancreatic (a) Neuro D and (b) Insulin genes expression among studied groups |
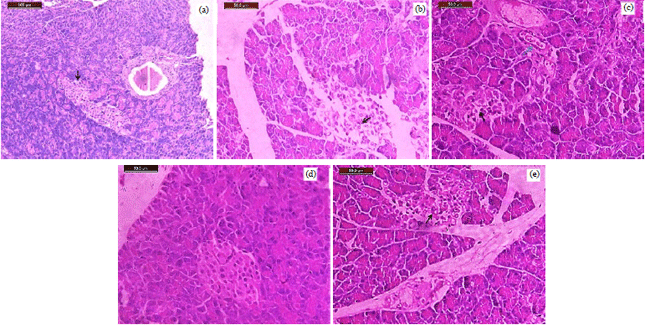 | |
| Fig. 7(a-e): | (a): Pancreas of the control group (2 weeks) showing normal architecture of endocrine and exocrine pancreatic cells. Black arrow indicated normal β cells of islets of Langerhans (100x), (b): Pancreas of STZ group showing massive vacuolation and necrosis of islets of Langerhans cells. Black arrow indicated massive destruction β cells in STZ induced rat (200x), (c): Pancreas of STZ+Un-MSCs group showing blue arrow at dilated blood vessels with inflammatory cells. Black arrow indicated regenerated β cells of islets of Langerhans (200x), (d): Pancreas of STZ+MSCs-Exs group showing a normal architecture of islets of Langerhans cells, Black arrow indicated regenerative β cells in STZ induced rat (200x), (e): Pancreas of STZ+Vit. D group showing vacuolation and necrosis of islets of Langerhans cells. Black arrow indicated destructive β cells in STZ induced rat (200x) |
EX-MSC treated group significantly upregulated Neuro D gene expression in pancreatic tissues compared to Vit. D and Un-MSC treated groups (Fig. 6a).
The pancreatic insulin gene expression level was significantly decreased in the STZ-treated group compared to the control group. Administration of Un-MSC, Vit. D and EX-MSC significantly increase insulin gene expression levels in the treated groups. EX-MSC-treated groups significantly increase insulin gene expression compared to Vit. D or Un-MSC treated groups (Fig. 6b).
Histopathology result: Pancreatic specimens from rats of each group were examined by light microscopy on low and high power. The STZ-treated group showed massive destruction of pancreatic tissues especially islets of Langerhans which decreased in number and size. Cells within islets showed necrosis and vacuolation. There is an inflammatory infiltrate. Islets still decreased in size with vasodilatation, edema, inflammatory infiltrate and signs of regeneration in specimens of the Un-MSCs-treated group. The pancreas of the STZ+ Vit. D group showed edema, vacuolation and necrosis of cells of islets of Langerhans. The pancreas of MSCs-Exs showed almost restoration of normal morphology with normal number size and number of islets and normal cellular morphology and disappearance of the destruction (Fig. 7).
DISCUSSION
In the present study, a single dose of STZ induced diabetes in the injected rats as proved by a significant decrease in serum insulin level and a significant increase in serum glucose compared to the control group. This finding is confirmed by histopathological examination which showed complete degeneration of beta cells and necrosis and islet of Langerhans.
The traditional treatment of type-1 diabetes is done by the administration of external insulin. Previously published studies from our lab aimed to specify and innovate diabetic treatment33.
In the current work, administration of Un-MSC, Ex-MSC, or Vit. D ameliorates STZ-induced diabetes effect by a significant increase in serum insulin and decreases serum glucose levels in rats of the treated groups.
Although Vit. D is not well established as a treatment option of type-1 diabetes, yet it represents promising pharmacological agents in the management of DM. Hence, we select Vit. D as a promising drug to be compared against other cell lines-based therapy in the current work.
The reports of Meta-analysis demonstrated the effect of Vit. D in the treatment of diabetes is controversial; where George et al.17, concluded that Vit. D decrease fasting plasma glucose level and improve insulin sensitivity comparing to the placebo group34,28,24,23,20, while the vice was reported by Seida et al.35. In a recent metanalysis, Vit. D supplementation significantly reduces HbA1c36.
The protective effect of Vit D observed her run with23, which may be due to its induction of glucose transport and activation of the insulin receptor gene transcription37, or through decrease the activation of c-Jun N-terminal protein kinase that is involved in triglyceride metabolism38.
Exogenous MSCs have been reported to have an anti-inflammatory effect39. MSCs are capable to differentiate into endothelial or epithelial cells, vascular smooth muscle40.
TGF-β is a cytokine involved in the regulation of diverse cellular processes including apoptosis by regulating the balance between apoptotic and pro-apoptotic genes expression41, activation of TGF-β induces phosphorylation of Smad2 and Smad3 with subsequent combination with Smad4 resulting in the initiation of transcription. The formed complex translocated to the nucleus to regulate targeted gene expression through direct binding to the Smad-binding element or indirectly through transcriptional coactivator association.
Currently, the STZ-treated group markedly downregulated pancreatic smad2 and smad3 expression levels. On the other hand, all treated groups significantly upregulated pancreatic smad2 and smad3 expression levels.
Pancreatic duodenal homeobox-1(PDX1) is a transcription factor involved in foregut and pancreas development during fetal life42, It controls the islet cell development and apoptosis.
In the current work, pancreatic PDX1 gene expression was significantly downregulated following STZ administration. PDX1 was upregulated in all treated rats, being more significant in Ex-MSc treated group compared to others. Our results run with a recent study reported that restoration of PDX1 expression inhibits Beta-cell apoptosis and improves the function of beta-cell43. Hani et al.44, that reported inducing of PDX1 gene expression can lead to the improvement of islet cells function and enhance its survival in an in vitro study. Rajaei et al.45 succeeded to generate in vitro cells with Glucose-Responsive insulin-secreting cells properties from PDX1-Overexpressing diabetic patients stem cells. Intense efforts are directed to identify agents that can be able to induce beta cell regeneration and insulin secretion. Paired box (PAX), is one of the transcriptional genes involved in cell organogenesis46, it plays a crucial role in plasticity and pancreas development.
Currently, STZ downregulates PAX4 gene expression, whereas Vit. D, Un-MSC, EX-MSC treated groups upregulate PAX4 in treated rats. EX-MSC-showed superior results compared to Vit. D and Un-MSC treated groups. The current results run with the Mellado-Gil et al.47, who showed the PAX4 overexpressing cells protect against STZ-induced DM in mice. Thus, PAX induced overexpression could be a good tool as a novel curative treatment of DM.
In the current work, the antiapoptotic effect of PDX1 and PAX4 upregulation in the treated groups was furtherly confirmed by histopathological examination of pancreatic tissues of the treated rats that showed improvement of pancreatic architecture and appearance of beta-cell cellular regeneration which run parallel to insulin gene expression.
NeuroD is a crucial transcription factor involved in pancreas development48 and activates the expression of glucokinase enzyme involved in glucose metabolism.
Currently, the NeuroD gene expression level was significantly decreased in the STZ-treated group. This is in coincides with previous studies that revealed; Neuro D is a vital transcription factor expressed in all pancreatic cells49. Neuro D gene expression level was upregulated in Un-MSC, exosome and Vit. D treated groups compared to the STZ-treated group. EX-MSC-treated group significantly increased Neuro D gene expression compared to Vit. D or Un-MSC treated groups.
Previous studies reported that Neuro D can induce insulin synthesis and release and can induce-also-beta-cell maturation and function50. In the present study, Ex-MSC treated group showed superior improvement relative to either UN-MSC or Vit. D.
Exosomes are an extracellular vesicle that contains molecular content of their cells of origin, such as proteins and different types of RNA. It represents a method of communication between cells as it can transfer molecules from one cell to another, thus exosomes can modulate the immune system51, mediating adaptive immune responses to pathogens and tumors52.
Exosomes have been recognized as a potential therapeutic agent as they can elicit potent cellular responses both in vitro and in vivo53. It has a powerful regenerative effect in damaged cells54. Thus exosomes can be considered as an important carrier for different types of RNA involved in many interfering functions55.
The better results obtained from Ex-MSC compared to MSC may be explained due to direct fusion of exosomes to its target cells enabling it to perform more biological effects, the facility of its storage for long period, easy control of its dose, time and route of administration and almost no risk of its rejection by the immune system or tumorigenesis associated with treatment by cell transplantation56.
In the current study, we evaluated the MSCs, MSCs-Ex and Vit. D against type-1 DM which revealed the superior beneficial effect of MSCs-Ex. This may represent a promising therapeutic tool for permanent Type-1 DM recovery. However different doses of MSCs-Ex should be tested to select the appropriate doses and recording of its related side effects before the clinical trial phase which was not done in the current project, we acknowledge this limitation and encourage the researcher to consider it in the upcoming studies.
CONCLUSION
Vit D, Un-MSC and Ex-MSC showed a protective effect against STZ-induced type 1 DM in rats. The superior result was found in Ex-MSC treated group with evidence of beta cell regeneration providing an important novel source for cell-based permanent treatment of type 1 diabetes.
SIGNIFICANCE STATEMENT
This study discovers the possible effectiveness of Mesenchymal Stem Cell-derived exosomes that can be beneficial for type-1 DM in rats. These results may represent a novel cell-based treatment agent of type-1 diabetes that will help a researcher to explore this field of treatment on their researches. Thus, new believe of permanent recovery from type-1 DM may be accessible.
ACKNOWLEDGMENT
This Research work Was Funded by The Scientific Research Deanship, Jouf University, Sakaka, Saudi Arabia (Project No. 39/603).
REFERENCES
- Czech, M.P., 2017. Insulin action and resistance in obesity and type 2 diabetes. Nat. Med., 23: 804-814.
CrossRefDirect Link - Balamurugan, A., R. Bottino, N. Giannoukakis and C. Smetanka, 2006. Prospective and challenges of islet transplantation for the therapy of autoimmune diabetes. Pancreas, 32: 231-243.
CrossRefDirect Link - Campagnoli, C., I.A.G. Roberts, S. Kumar, P.R. Bennett, I. Bellantuono and N.M. Fisk, 2001. Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver and bone marrow. Blood, 98: 2396-2402.
CrossRefDirect Link - Chen, Y., J.Z. Shao, L.X. Xiang, X.J. Dong and G.R. Zhang, 2008. Mesenchymal stem cells: A promising candidate in regenerative medicine. Int. J. Biochem. Cell Biol., 40: 815-820.
CrossRefDirect Link - Gao, F., S.M. Chiu, D.A.L. Motan, Z. Zhang and L. Chen et al., 2016. Mesenchymal stem cells and immunomodulation: current status and future prospects. Cell Death Dis., 7: e2062-e2062.
CrossRefDirect Link - Aggarwal, S. and M.F. Pittenger, 2005. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood, 105: 1815-1822.
CrossRefDirect Link - Erices, A., P. Conget and J.J. Minguell, 2000. Mesenchymal progenitor cells in human umbilical cord blood. Br. J. Haematol., 109: 235-242.
CrossRefDirect Link - Zuk, P.A., M. Zhu, H. Mizuno, J. Huang and J.W. Futrell et al., 2001. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng., 7: 211-228.
CrossRefDirect Link - De Bari, C., F. Dell'Accio, P. Tylzanowski and F.P. Luyten, 2001. Multipotent mesenchymal stem cells from adult human synovial membrane. Arthritis Rheumatism, 44: 1928-1942.
CrossRefDirect Link - Matsumoto, R., T. Omura, M. Yoshiyama, T. Hayashi and S. Inamoto et al., 2005. Vascular endothelial growth factor-expressing mesenchymal stem cell transplantation for the treatment of acute myocardial infarction. Arteriosclerosis, Thrombosis, Vascular Biol., 25: 1168-1173.
CrossRefDirect Link - Jiang, Y., B.N. Jahagirdar, R.L. Reinhardt, R.E. Schwartz and C.D. Keene et al., 2002. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature, 418: 41-49.
CrossRefDirect Link - Winston, C.N., E.J. Goetzl, J.B. Schwartz, F.M. Elahi and R.A. Rissman, 2019. Complement protein levels in plasma astrocyte‐derived exosomes are abnormal in conversion from mild cognitive impairment to Alzheimer's disease dementia. Alzheimer's Dementia: Diagn., Assess. Dis. Monit., 11: 61-66.
CrossRefDirect Link - Lai, R.C., S.S. Tan, B.J. Teh, S.K. Sze and F. Arslan et al., 2012. Proteolytic potential of the MSC exosome proteome: implications for an exosome-mediated delivery of therapeutic proteasome. Int. J. Proteomics, 2012: 1-14.
CrossRefDirect Link - Raposo, G. and W. Stoorvogel, 2013. Extracellular vesicles: Exosomes, microvesicles and friends. J. Cell. Biol., 200: 373-383.
CrossRefDirect Link - Stahl, P.D. and G. Raposo, 2018. Exosomes and extracellular vesicles: the path forward. Essays Biochem., 62: 119-124.
CrossRefDirect Link - Johnstone, R.M., M. Adam, J. Hammond, L. Orr and C. Turbide, 1987. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem., 262: 9412-9420.
Direct Link - Shi, B., Y. Wang, R. Zhao, X. Long, W. Deng and Z. Wang, 2018. Bone marrow mesenchymal stem cell-derived exosomal miR-21 protects C-kit+ cardiac stem cells from oxidative injury through the PTEN/PI3K/Akt axis. PLoS ONE, Vol. 13.
CrossRefDirect Link - Chen, T., F. Arslan, Y. Yin, S. Tan and R. Lai et al., 2011. Enabling a robust scalable manufacturing process for therapeutic exosomes through oncogenic immortalization of human ESC-derived MSCs. J. Translational Med.
CrossRefDirect Link - Mattila, C., P. Knekt, S. Mannisto, H. Rissanen, M. A. Laaksonen, J. Montonen and A. Reunanen, 2007. Serum 25-Hydroxyvitamin D concentration and subsequent risk of type 2 diabetes. Diabetes Care, 30: 2569-2570.
CrossRefDirect Link - Mitri, J. and A.G. Pittas, 2014. Vitamin D and Diabetes. Endocrinol. Metab. Clin. North Am., 43: 205-232.
CrossRefDirect Link - Pittas, A.G., J. Lau, F.B. Hu and B. Dawson-Hughes, 2007. The role of Vitamin D and Calcium in Type 2 diabetes. A systematic review and meta-analysis. J. Clin. Endocrinol. Metab., 92: 2017-2029.
CrossRefDirect Link - Amin, S.N., U.K. Hussein, H.D. Yassa, S.S. Hassan and L.A. Rashed, 2018. Synergistic actions of vitamin D and metformin on skeletal muscles and insulin resistance of type 2 diabetic rats. J. Cell Physiol., 233: 5768-5779.
CrossRefDirect Link - Ross, A.C., 2011. The 2011 report on dietary reference intakes for calcium and vitamin D. Public Health Nutr., 14: 938-939.
CrossRefDirect Link - Zeitz, U., K. Weber, D.W. Soegiarto, E. Wolf, R. Balling and R.G. Erben, 2003. Impaired insulin secretory capacity in mice lacking a functional vitamin D receptor. FASEB J., 17: 1-14.
CrossRefDirect Link - Abdel Aziz, M.T., H.M. Atta, S. Mahfouz, H.H. Fouad and N.K. Roshdy et al., 2007. Therapeutic potential of bone marrow-derived mesenchymal stem cells on experimental liver fibrosis. Clin. Biochem., 40: 893-899.
CrossRefDirect Link - Qu, Y., Q. Zhang, X. Cai, F. Li, Z. Ma, M. Xu and L. Lu, 2017. Exosomes derived from miR-181-5p-modified adipose-derived mesenchymal stem cells prevent liver fibrosis via autophagy activation. J. Cell. Mol. Med., 21: 2491-2502.
CrossRefDirect Link - Akagawa, M., N. Miyakoshi, Y. Kasukawa, Y. Ono and Y. Yuasa et al., 2018. Effects of activated vitamin D, alfacalcidol, and low-intensity aerobic exercise on osteopenia and muscle atrophy in type 2 diabetes mellitus model rats. PLoS ONE, Vol. 13.
CrossRefDirect Link - Menck, K., A. Bleckmann, A. Wachter, B. Hennies and L. Ries et al., 2018. Characterisation of tumour-derived microvesicles in cancer patients’ blood and correlation with clinical outcome. J. Extracellular Vesicles, Vol. 6.
CrossRefDirect Link - Van Der Pol, E., T. Van Leeuwen and R. Nieuwland, 2014. An Overview of Novel and Conventional Methods to Detect Extracellular Vesicles. 1st Edn., Pan Stanford Publishing, Singapore, Pages: 32.
CrossRefDirect Link - Wassef, M.A., H. Fouad, D. Sabry, N. Afifi, A.M. Abbas, W. Mostafa and S.H. Ahmed, 2016. Therapeutic efficacy of differentiated versus undifferentiated mesenchymal stem cells in experimental type I diabetes in rat. Biochem. Biophys. Rep., 5: 468-475.
CrossRefDirect Link - Bancroft, J.D., 2008. Theory and Practice of Histological Techniques. Elsevier Health Sciences, Philadelphia, Pennsylvania, ISBN-13: 9780443102790, Pages: 725.
Direct Link - Mostafa-Hedeab G., A.A. Mohamed, G. Thabet, D. Sabry, R.F. Salam and M.E. Hassen, 2018. Effect of mate 1, mate 2 and oct1 single nucleotide polymorphisms on metformin action in recently diagnosed egyptian type-2 diabetic patients. Biomedical and Pharmacology Journal 11: 149-157.
CrossRefDirect Link - George, P.S., E.R. Pearson and M.D. Witham, 2012. Effect of vitamin D supplementation on glycaemic control and insulin resistance: a systematic review and meta-analysis. Diabetic Med., 29: e142-e150.
CrossRefDirect Link - Seida, J.C., J. Mitri, I.N. Colmers, S.R. Majumdar and M.B. Davidson et al., 2014. Effect of vitamin D3Supplementation on improving glucose homeostasis and preventing diabetes: A systematic review and meta-analysis. J. Clin. Endocrinol. Metab., 99: 3551-3560.
CrossRefDirect Link - Wu, C., S. Qiu, X. Zhu and L. Li, 2017. Vitamin D supplementation and glycemic control in type 2 diabetes patients: A systematic review and meta-analysis. Metab., 73: 67-76.
CrossRefDirect Link - Alvarez, J.A. and A. Ashraf, 2010. Role of vitamin d in insulin secretion and insulin sensitivity for glucose homeostasis. Int. J. Endocrinol.
CrossRefDirect Link - Zhou, Q.G., F.F. Hou, Z.J. Guo, M. Liang, G.B. Wang and X. Zhang, 2008. 1,25-Dihydroxyvitamin D improved the free fatty-acid-induced insulin resistance in cultured C2C12 cells. Diabetes Metab. Res. Rev., 24: 459-464.
CrossRefDirect Link - Ortiz, L.A., F. Gambelli, C. McBride, D. Gaupp, M. Baddoo, N. Kaminski and D.G. Phinney, 2003. Mesenchymal stem cell engraftment in lung is enhanced in response to bleomycin exposure and ameliorates its fibrotic effects. Proc. Natl. Acad. Sci., 100: 8407-8411.
CrossRefDirect Link - Uccelli, A., L. Moretta and V. Pistoia, 2008. Mesenchymal stem cells in health and disease. Nat. Rev. Immunol., 8: 726-736.
CrossRefDirect Link - Spender, L.C., D.I.O. Brien, D. Simpson, D. Dutt and C.D. Gregory et al., 2009. TGF-β induces apoptosis in human B cells by transcriptional regulation of BIK and BCL-XL. Cell Death Differ., 16: 593-602.
CrossRefDirect Link - Lai, Y., H. Brandhorst, H. Hossain, A. Bierhaus, C. Chen, R.G. Bretzel and T. Linn, 2009. Activation of NFκB dependent apoptotic pathway in pancreatic islet cells by hypoxia. Islets, 1: 19-25.
CrossRefDirect Link - Kornelius, E., H.H. Li, C.H. Peng, Y.S. Yang and W.J. Chen et al., 2018. Liraglutide protects against glucolipotoxicity-induced RIN-m5F β-cell apoptosis through restoration of PDX1 expression. J. Cell Mol. Med., 23: 619-629.
CrossRefDirect Link - Hani, H., Z.N. Allaudin, M.A. Mohd-Lila, K. Sarsaifi and M. Rasouli et al., 2017. Improvement of isolated caprine islet survival and functionality in vitro by enhancing of PDX1 gene expression. Xenotransplantation, Vol. 24.
CrossRefDirect Link - Rajaei, B., M. Shamsara and M.H. Sanati, 2018. In vitro generation of glucose-responsive insulin-secreting cells from PDX1–overexpressing human-induced pluripotent stem cell derived from diabetic patient. ASAIO J., 64: 819-826.
CrossRefDirect Link - Paixão-Côrtes, V.R., F.M. Salzano and M.C. Bortolini, 2015. Origins and evolvability of the PAX family. Seminars Cell Dev. Biol., 44: 64-74.
CrossRefDirect Link - Mellado-Gil, J.M., C.M. Jiménez-Moreno, A. Martin-Montalvo, A.I. Alvarez-Mercado and E. Fuente-Martin et al., 2016. PAX4 preserves endoplasmic reticulum integrity preventing beta cell degeneration in a mouse model of type 1 diabetes mellitus. Diabetologia, 59: 755-765.
CrossRefDirect Link - Chao, C.S., Z.L. Loomis, J.E. Lee and L. Sussel, 2007. Genetic identification of a novel NeuroD1 function in the early differentiation of islet α, PP and ε cells. Dev. Biol., 312: 523-532.
CrossRefDirect Link - Inoue, Y. and S. Hayashi, 2007. Tissue-specific laminin expression facilitates integrin-dependent association of the embryonic wing disc with the trachea in Drosophila. Dev. Biol., 304: 90-101.
CrossRefDirect Link - Gu, C., G.H. Stein, N. Pan, S. Goebbels and H. Hornberg et al., 2010. Pancreatic β cells require neurod to achieve and maintain functional maturity. Cell Metab., 11: 298-310.
CrossRef - Oushy, S., J.E. Hellwinkel, M. Wang, G.J. Nguyen and D. Gunaydin et al., 2017. Glioblastoma multiforme-derived extracellular vesicles drive normal astrocytes towards a tumour-enhancing phenotype. Phil. Trans. R. Soc. B, Vol. 373.
CrossRefDirect Link - Hough, K.P., D. Chanda, S.R. Duncan, V.J. Thannickal and J.S. Deshane, 2017. Exosomes in immunoregulation of chronic lung diseases. Allergy, 72: 534-544.
CrossRefDirect Link - Di Rocco, G., S. Baldari and G. Toietta, 2016. Towards therapeutic delivery of extracellular vesicles: Strategies for in vivo tracking and biodistribution analysis. Stem Cells Int., 2016: 1-12.
CrossRefDirect Link - Basu, J. and J.W. Ludlow, 2016. Exosomes for repair, regeneration and rejuvenation. Expert Opin. Biol. Ther., 16: 489-506.
CrossRefDirect Link - Wahlgren, J., L. Statello, G. Skogberg, E. Telemo and H. Valadi, 2015. Delivery of small interfering RNAs to cells via exosomes. In: SiRNA Delivery Methods, Shum, K. and J. Rossi, Springer, New York pp: 105-125.
CrossRefDirect Link - Lu, K., H.Y. Li, K. Yang, J.L. Wu, X.W. Cai, Y. Zhou and C.Q. Li, 2017. Exosomes as potential alternatives to stem cell therapy for intervertebral disc degeneration: in-vitro study on exosomes in interaction of nucleus pulposus cells and bone marrow mesenchymal stem cells. Stem Cell Res. Ther., Vol. 8.
CrossRefDirect Link