
Fakhria Al-Joufi
Department of Pharmacology, Faculty of Pharmacy, Jouf University, Saudi Arabia
LiveDNA: 966.33279
International Journal of Pharmacology
Year: 2020 | Volume: 16 | Issue: 5 | Page No.: 398-406







ABSTRACT
Background and Objective: Hypercholesterolemia is a risk factor for many cardiac and hepatic disorders. Visnagin (VIS) is a coumarin with an anti-inflammatory effect; however, its anti-dyslipidemia activity has not been investigated. This study evaluated the effect of VIS on hypercholesterolemia, oxidative stress and inflammatory response in high cholesterol diet (HCD)-fed rats. Materials and Methods: Rats received an HCD and VIS (30 and 60 mg kg–1) for 10 weeks and samples were harvested. Results: HCD increased serum cholesterol, triglycerides, LDL, VLDL, transaminases, CK-MB and pro-inflammatory cytokines and decreased HDL-cholesterol. VIS ameliorated serum lipids, liver and heart functions and cardiovascular risk indices in HCD-fed rats. Besides, VIS suppressed hepatic lipid peroxidation and increased the levels of cellular antioxidants. The effects of VIS were associated with increased hepatic LDL receptor (LDLR) and decreased HMG-CoA reductase (HMGR) expression. Conclusion: These results demonstrate the anti-hypercholesterolemic activity of VIS in rats. VIS ameliorated serum lipids, inflammation, liver and heart function and oxidative stress and modulated hepatic HMGR and LDLR. Therefore, VIS is a promising candidate for the treatment of hypercholesterolemia.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Fakhria Al-Joufi, 2020. Prevention of Metabolic, Redox and Lipid Biosynthesis Alterations by Visnagin in High Cholesterol-Fed Rats. International Journal of Pharmacology, 16: 398-406.
DOI: 10.3923/ijp.2020.398.406
URL: https://scialert.net/abstract/?doi=ijp.2020.398.406
DOI: 10.3923/ijp.2020.398.406
URL: https://scialert.net/abstract/?doi=ijp.2020.398.406
INTRODUCTION
Dyslipidemia is a disorder of lipid metabolism and its prevalence has dramatically increased in recent years. This metabolic abnormality is caused by the consumption of high fat/cholesterol foods as well as other environmental and genetic factors1-3. Hypercholesterolemia is a dyslipidemic condition associated with the development of cardiovascular diseases (CVD), steatosis, hypertension and other diseases4,5. Hypercholesterolemia and other dyslipidemias increase hepatic both lipid production and accumulation resulting in fatty liver, injury, dysfunction, fibrosis and other complications1,6. Different studies have shown elevated reactive oxygen species (ROS) levels and oxidative stress in different cells, including hepatocytes, endothelium and cardiac cells. Increased ROS causes oxidative stress, lipid peroxidation (LPO) and cell death1,7. Accordingly, several researchers reported oxidative stress in the liver and the heart of rats received a High Cholesterol Diet (HCD)3,8-10. The relationship between inflammation and dyslipidemia/ hypercholesterolemia has been acknowledged in recent studies3,8-10. Tumor necrosis factor (TNF)-α and other inflammatory mediators such as interleukin (IL)-6 have increased in HCD-fed rodents3,8,9, demonstrating the development of an inflammatory response. In addition, oxidation of Low-Density Lipoprotein (LDL) with ROS stimulates inflammation and the production of more ROS11. Inflammation in hypercholesterolemia was associated with various pathological conditions, including steatosis and atherosclerosis12. Thus, the management of hypercholesterolemia and its linked excess ROS and cytokines can prevent its deleterious complications in different organs, particularly liver and heart.
Many plants and natural products with antioxidant activity showed beneficial effects against abnormal lipid metabolism in different conditions3,8-10,13-15. Visnagin (VIS) is a furanochromone (Fig. 1) that belongs to coumarins and possesses multiple pharmacologic properties16.
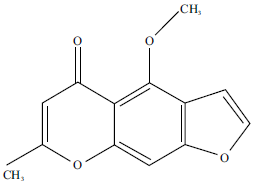 | |
| Fig. 1: | Chemical structure of visnagin |
VIS exerted strong antioxidant and anti-inflammatory activities in different animal models16,17. In microglial cells, VIS prevented lipopolysaccharides (LPS)-induced inflammation and suppressed TNF-α and IL-618. Other studies have demonstrated the anti-inflammatory potential of VIS where it ameliorated acute pancreatitis in mice19. Also, VIS improved the antioxidant defenses and attenuated oxidative stress in a mouse model of pancreatitis19. Although studies have shown promising pharmacological activities of VIS, its effect on dyslipidemia has not been previously explored. This study investigated the role of VIS in ameliorating hypercholesterolemia, oxidative stress and inflammatory response in HCD-fed rats. In addition, the effect of VIS on the expression of HMG-CoA reductase (HMGR) and LDL receptor (LDLR) has been evaluated.
MATERIALS AND METHODS
Study area: This study was conducted at the Department of Pharmacology, Faculty of Pharmacy, Jouf University, Saudi Arabia during the period from February to December 2019.
Induction of hypercholesterolemia and experimental design: Total 24 (8-week-old) male albino rats weighing 160-180 g were included. The animals were housed under a 12 h light/dark cycle and 23±2°C and supplied food and water ad libitum for one week before starting the experiment. Protocols involving the use of animals were performed in line with the guidelines of the National Institutes of Health (NIH publication No. 85-23, revised 2011).
Hypercholesterolemia was induced by feeding the animals an HCD consists of the normal diet and 2% cholesterol for 10 weeks3,9, whereas control rats received normal diet for the same period. The rats were divided into 4 groups (six rats each) as follows:
| Group 1: | Control |
| Group 2: | HCD |
| Group 3: | HCD-fed rats treated with 30 mg kg–1 VIS (Sigma, USA)19 daily by oral gavage for 10 weeks |
| Group 4: | HCD-fed rats treated with 60 mg kg–1 VIS19 daily by oral gavage for 10 weeks |
At the end of the experiment, overnight fasted rats were anesthetized and blood was collected via cardiac puncture for serum preparation. The animals were dissected and the liver was removed and frozen at -80°C.
| Table 1: | Primers sequences |
 | |
Assay of lipids and cardiovascular risk indices: Serum lipids (total cholesterol, triglycerides and HDL-cholesterol) were assayed using assay kits (Biosystems, Spain). The following equations were used to calculate LDL-cholesterol, VLDL-cholesterol and cardiovascular risk indices20:
VLDL-cholesterol = Triglycerides/5
LDL-cholesterol = Total cholesterol-(HDL-cholesterol+VLDL-cholesterol)
Cardiovascular risk index 1 = Total-cholesterol/HDL-cholesterol
Cardiovascular risk index 2 = LDL-cholesterol/HDL-cholesterol
Assay of transaminases, creatine kinase (CK)-MB and pro-inflammatory cytokines: Serum alanine aminotransferase (ALT), aspartate aminotransferase (AST) and CK-MB were assayed using Biosystems (Spain) kits. Serum IL-6 and TNF-α were determined by R and D Systems (USA) ELISA kits.
Assay of LPO and antioxidant: The liver was homogenized (10% w/v) in cold phosphate-buffered saline (PBS), centrifuged and the clear homogenate was separated. Thiobarbituric acid reactive substances (TBARS), a marker of LPO21, reduced glutathione (GSH)22, superoxide dismutase (SOD)23 and catalase24 were assayed in the supernatant.
Assay of gene expression levels: Gene expression of HMGR and LDLR was determined by qRT-PCR as previously described25. Briefly, total RNA was isolated using an RNA isolation kit (Qiagen, Germany) and quantified at 260 nm using a nanodrop. RNA samples with A260/A280 nm>1.8 were reverse transcribed using a reverse transcription kit (Applied Biosystems, USA). cDNA was amplified using SYBR Green master mix (Qiagen, Germany) and primers in Table 1. Gene expression was calculated using the 2ΔΔCt method26.
Statistical analysis: All results were expressed as the Mean±standard error of the mean (SEM) and the differences between mean values of multiple groups were analyzed using one-way ANOVA followed by Tukey’s test on Graphpad Prism 7. Statistical significance was considered at P less than 0.05.
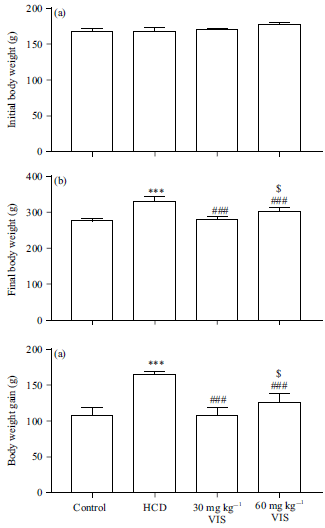 | |
| Fig. 2(a-c): | Effect of visnagin on body weight changes in HCD-fed rats, (a) Initial body weight, (b) Final body weight and (c) Body weight gain |
| Data are Mean±SEM (n = 6), ***p<0.001 compared to control, ###p<0.001 compared to HCD, $p<0.05 compared to 30 mg kg–1 visnagin | |
RESULTS
Effect of VIS on body weight changes in HCD-fed rats: The initial body weight of all groups wasn’t significantly different as illustrated in Fig. 2a. After 10 weeks, HCD induced significant body weight gain (Fig. 2b-c). Treatment with VIS decreased body weight gain in HCD-fed rats.
Effect of VIS on serum lipids in HCD-fed rats: HCD supplementation caused hypertriglyceridemia (Fig. 3a) and hypercholesterolemia (Fig. 3b), increased serum LDL (Fig. 3c) and VLDL (Fig. 3d) significantly (p<0.001).
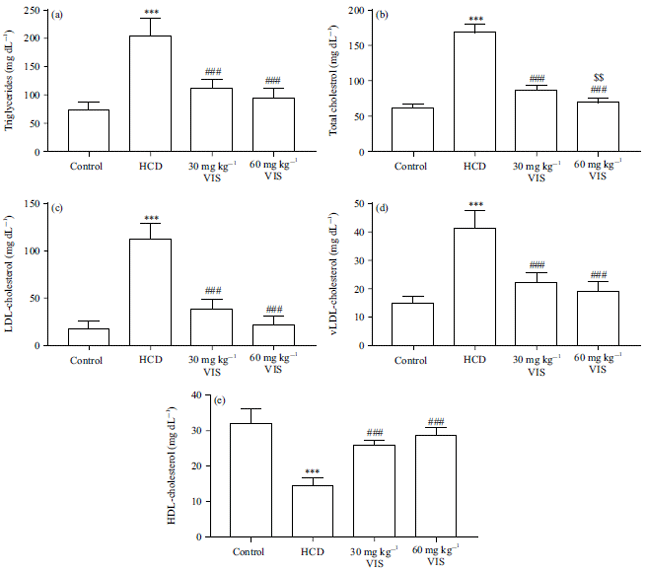 | |
| Fig. 3(a-e): | Visnagin decreased serum (a) Triglycerides, (b) Total cholesterol, (c) LDL-cholesterol and (d) vLDL-cholesteroland increased (e) HDL-cholesterol in HCD-fed rats |
| Data are Mean±SEM (n = 6), ***p<0.001 compared to Control, ###p<0.001 compared to HCD, $$p<0.01 compared to 30 mg kg–1 visnagin | |
On the other hand, serum HDL was decreased significantly (p<0.001) in rats fed an HCD (Fig. 3e). VIS markedly decreased triglycerides and total-, LDL and VLDL cholesterols, whereas increased serum HDL in HCD-challenged rats. The 60 mg kg–1 VIS dose decreased total cholesterol significantly (p<0.01) as compared to the lower dose.
Effect of VIS on HMGR and LDLR expression in HCD-fed rats: The expression of HMGR in the liver of the HCD-fed rat showed a significant increase (p<0.001) as compared to the normal group (Fig. 4a). The low dose of VIS (30 mg) decreased the expression of HMGR hepatic significantly. The higher dose of VIS was more effective in decreasing HMGR expression when compared with the HCD-fed rats (p<0.001) and the lower dose (p<0.01). On the other hand, HCD decreased hepatic LDLR in rats whereas treatment with VIS significantly restored it (Fig. 4b). The effect of VIS on LDLR expression was dose-dependent.
Effect of VIS on liver and heart function in HCD-fed rats: Serum transaminases and CK-MB were assessed to determine the effect of VIS on liver and heart function in HCD-fed rats. As shown in Fig. 5a and b, HCD increased serum ALT and AST activities, respectively (p<0.001). Similarly, HCD-fed rats showed increased serum CK-MB activity as compared to the control (p<0.001, Fig. 5c). VIS (30 and 60 mg kg–1) decreased serum ALT, AST and CK-MB significantly in HCD-fed rats. The effect of VIS on AST and CK-MB was dose-dependent.
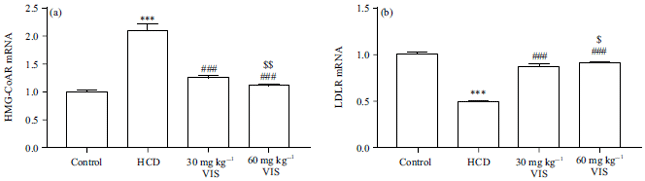 | |
| Fig. 4(a-b): | Visnagin decreased (a) HMGR and (b) Increased LDLR expression in liver of HCD-fed rats |
| Data are Mean±SEM (n = 6), ***p<0.001 compared to Control, ###p<0.001 compared to HCD, $p<0.05 and $$p<0.01 compared to 30 mg kg–1 visnagin | |
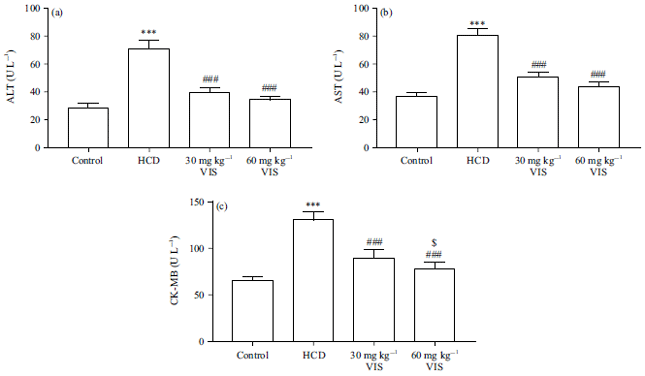 | |
| Fig. 5(a-c): | Visnagin decreased serum (a) ALT, (b) AST and (c) CK-MB in HCD-fed rats |
| Data are Mean±SEM (n = 6), ***p<0.001 compared to Control, ###p<0.001 compared to HCD, $p<0.05 compared to 30 mg kg–1 visnagin | |
Effect of VIS on cardiovascular risk induces in HCD-fed rats: Given the role of HCD in causing dyslipidemia and cardiac dysfunction, the effect of VIS on cardiovascular risk indices was determined. HCD feeding increased total cholesterol- and LDL-cholesterol/HDL-cholesterol significantly as depicted in Fig. 6a and 6b, respectively. VIS exerted cardioprotective effect by decreasing these ratios significantly in HCD-fed rats.
Effect of VIS on pro-inflammatory cytokines in HCD-fed rats: Serum IL-6 (Fig. 7a) and TNF-α (Fig. 7b) were increased in HCD-fed rats significantly as compared to the normal group (p<0.001). Both the 30 and 60 mg kg–1 doses of VIS decreased these serum cytokines in HCD-fed rats. The effect of VIS on serum IL-6 and TNF-α was dose-dependent.
Effect of VIS on redox status in HCD-fed rats: Liver TBARS were increased significantly in HCD-fed rats as compared with the normal as represented in Fig. 8a (p<0.001). On the contrary, GSH, SOD and catalase were declined in the liver of rats received HCD as depicted in Fig. 8b-d. VIS significantly decreased TBARS and increased antioxidants in the liver of HCD-fed rats with a dose-dependent effect on catalase activity (Fig. 8d).
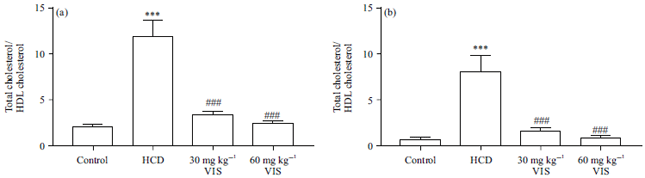 | |
| Fig. 6(a-b): | Visnagin decreased cardiovascular risk induces in HCD-fed rats |
Data are Mean±SEM (n = 6), ***p<0.001 compared to Control, ###p<0.001 compared to HCD | |
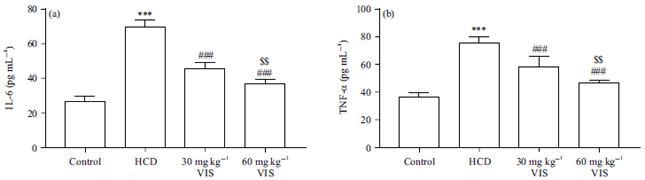 | |
| Fig. 7(a-b): | Visnagin decreased serum (a) IL-6 and (b) TNF-α in HCD-fed rats |
Data are Mean±SEM (n = 6), ***p<0.001 compared to Control, ###p<0.001 compared to HCD, $$p<0.01 compared to 30 mg kg–1 visnagin | |
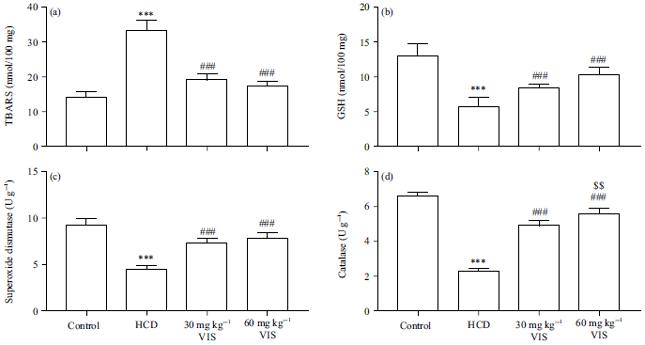 | |
| Fig. 8(a-b): | Visnagin decreased (a) TBARS (b) Increased GSH, (c) Superoxide dismutase and (d) Catalase in liver of HCD-fed rats |
| Data are Mean±SEM (n = 6), ***p<0.001 compared to Control, ##p<0.01 and ###p<0.001 compared to HCD, $$p<0.01 compared to 30 mg kg–1 visnagin | |
DISCUSSION
This study showed the ameliorative effect of VIS on serum lipids, liver and heart function, cardiovascular indices and hepatic TBARS in hypercholesterolemic rats. Besides, VIS increased hepatic antioxidant and LDLR expression and downregulated HMGR. Hypercholesterolemia is a risk factor of CVD and causes damage to different organs, including the liver1. It provokes oxidative stress and chronic inflammation which plays a role in worsening dyslipidemia and leads to cell death1,7. Therefore, agents with anti-hyperlipidemic, antioxidant and anti-inflammatory effects would protect against the damaging consequences of hypercholesterolemia. VIS is a coumarin which can suppress oxidative stress and attenuate inflammatory responses; however, its effect on hypercholesterolemia and its associated derangements is not known. This study is the first to show the anti-hypercholesterolemic potential of VIS as well as its ability to prevent inflammation and oxidative injury.
Hypercholesterolemia was established by feeding rats an HCD for 10 weeks. Besides the increase in body weight gain, serum cholesterol, triglycerides, VLDL and LDL were increased whereas HDL was declined in HCD-fed rats. In agreement with these results, previous studies have shown increased body weight and serum lipids in the same experimental model3,8,9. This model mimics hypercholesterolemia in humans and is therefore of great value for pharmacological investigations27. VIS prevented the significant weight gain induced by HCD, ameliorated triglycerides and cholesterol and enhanced HDL levels, demonstrating a potent anti-hyperlipidemic effect. Although the anti-dyslipidemia mechanism of VIS has not previously explained, this study showed its modulatory effects on the expression levels of HMGR and LDLR. HCD feeding increased hepatic HMGR and reduced LDLR whereas VIS reversed these effects. HMGR is the rate-limiting enzyme of cholesterol biosynthesis and is an important target for cholesterol-lowering drugs28. HCD stimulates HMGR expression and activation leading to elevated LDL-cholesterol and accumulation of cholesterol in the liver27,29. In agreement with this notion, feeding an HCD for 10 weeks has been recently reported to increase hepatic cholesterol levels9. Thus, suppressing HMGR represents the main contributor to the anti-hypercholesterolemic effect of VIS. Also, increased LDLR contributes to the ameliorative effect of VIS in hypercholesterolemic rats. LDLR is responsible for LDL-cholesterol binding and internalization into hepatocytes leading to decreased serum cholesterol. Thus, increasing the expression of LDLR is an effective way to reduce blood and liver cholesterol30.
The liver injury occurs as a consequence of the increased accumulation of cholesterol as previously reported1,6,9. HCD has been recently reported to cause liver injury shown as steatosis, inflammatory cell infiltration and increased serum transaminases9. In this study, HCD increased serum ALT and AST as a result of liver injury. ALT and AST are indicators for the assessment of liver injury and are considered valuable markers of degenerative and necrotic changes in hepatocytes31. HCD induced cardiac injury as shown by increased serum CK-MB as reported in previous studies3,9. Also, cardiovascular risk indices have been increased in HCD-fed rats, confirming the increased CK-MB levels. Interestingly, VIS ameliorated serum ALT, AST, CK-MB and cardiovascular risk indices. Therefore, VIS has hepatic- and cardioprotective effects in HCD-fed rats. Although reports on the hepatoprotective activity of VIS are scarce, its cardioprotective effect in doxorubicin-induced zebrafish as a model of cardiomyopathy32. Because the hypercholesterolemia-induced liver injury is associated with oxidative stress and the development of chronic inflammation3,8-10, the protective effect of VIS is explained in terms of its anti-hypercholesterolemic as well as radical-scavenging and anti-inflammatory activities. In the present study, HCD caused oxidative stress and inflammation as shown by increased liver LPO and serum IL-6 and TNF-α and decreased liver GSH and antioxidant enzymes. Hypercholesterolemia induces oxidative modification of LDL, protein and glucose33 and stimulated NADPH oxidase to produce ROS and consequently LPO and cell death33. LPO exerts a negative impact on cell membrane fluidity and permeability, leading to the destruction of the membrane33,34. In addition, increased ROS causes oxidation of the antioxidant enzymes and therefore induce oxidative stress34. Excess ROS and oxidized LDL stimulate inflammatory responses by increasing NF-κB signaling and the release of IL-6 and TNF-α as well as many other mediators. This was confirmed by increased serum IL-6 and TNF-α in rats fed the HCD for 10 weeks, indicating chronic inflammation which promotes different pathologies such as atherosclerosis and other CVD12. Interestingly, the anti-hypercholesterolemic effect of VIS was associated with attenuated oxidative stress and inflammation. The ability of VIS to protect against inflammation and oxidative stress has been reported in previous studies. In LPS-challenged microglial cells in vitro, VIS exerted a strong anti-inflammatory effect and decreased the expression of IL-6 and TNF-α18. Similar effects were reported in vivo where VIS ameliorated oxidative stress and inflammation in a mouse model of acute pancreatitis19. VIS suppressed NF-κB and abolished the expression of TNF-α and IL-619. The findings of this study showed the potent antioxidant and anti-inflammatory activities of VIS in hypercholesterolemic rats.
CONCLUSION
This study shows for the first time the anti-hypercholesterolemic activity of VIS in rats. VIS ameliorated serum cholesterol and triglycerides increased HDL-cholesterol and reduced cardiovascular risk. The mechanism of VIS involves its modulatory effect on hepatic HMGR and LDLR. VIS exerted hepato and cardioprotective effects where it decreased serum transaminases and CK-MB in HCD-fed rats. In addition, VIS attenuated oxidative stress and inflammation and enhanced antioxidants in HCD-fed rats. These findings demonstrate that VIS has a potent anti-dyslipidemic effect is a promising agent for the treatment of hypercholesterolemia.
SIGNIFICANCE STATEMENT
Hypercholesterolemia is a major risk factor for many disorders, including cardiovascular and liver diseases. This study shows the novel anti-hypercholesterolemic effect of the furanochromone visnagin and its hepatoprotective activity. Visnagin modulates HMGR and LDLR, suppress oxidative stress and attenuate inflammation. Therefore, this furanochromone is a promising candidate for the development of an effective anti-hyperlipidemic agent.
REFERENCES
- Sozen, E., and N.K. Ozer, 2017. Impact of high cholesterol and endoplasmic reticulum stress on metabolic diseases: An updated mini-review. Redox Biol., 12: 456-461.
CrossRefDirect Link - Ordovas, J.M., 2009. Genetic influences on blood lipids and cardiovascular disease risk: tools for primary prevention. Am. J. Clin. Nutr., 89: 1509S-1517S.
CrossRefDirect Link - Bin-Jumah, M.N., 2018. Monolluma quadrangula protects against oxidative stress and modulates LDL receptor and fatty acid synthase gene expression in hypercholesterolemic rats. Oxid. Med. Cell. Longevity, Vol. 2018.
CrossRefDirect Link - Han, Q., S.C. Yeung, M.S.M. Ip and J.C.W. Mak, 2018. Dysregulation of cardiac lipid parameters in high-fat high-cholesterol diet-induced rat model. Lipids Health Dis.
CrossRefDirect Link - Cífková, R., and A. Krajčoviechová, 2015. Dyslipidemia and cardiovascular disease in women. Curr. Cardiol. Rep.
CrossRefDirect Link - Farrell, G.C and C.Z. Larter, 2006. Nonalcoholic fatty liver disease: From steatosis to cirrhosis. Hepatol., 43: S99-S112.
CrossRefPubMedDirect Link - Förstermann, U., 2008. Oxidative stress in vascular disease: causes, defense mechanisms and potential therapies. Nat. Rev. Cardiol., 5: 338-349.
CrossRefDirect Link - Aloud, A.A., 2019. Protective effect of morin, a flavonoid against hypercholesterolemia-induced hepatic and renal toxicities in rats. J. Appl. Sci., 19: 575-584.
CrossRefDirect Link - Abukhalil, M.H., O.E. Hussein, M. Bin-Jumah, S.A.M. Saghir and M.O. Germoush et al., 2020. Farnesol attenuates oxidative stress and liver injury and modulates fatty acid synthase and acetyl-CoA carboxylase in high cholesterol-fed rats. Environ. Sci. Pollut. Res.
CrossRefDirect Link - Aladaileh, S.H., S.A. Saghir, K. Murugesu, A. Sadikun and A. Ahmad et al., 2019. Antihyperlipidemic and antioxidant effects of Averrhoa carambola extract in high-fat diet-fed rats. Biomedicines, Vol. 7, No. 3.
CrossRefDirect Link - Lara-Guzmán, O.J., Á. Gil-Izquierdo, S. Medina, E. Osorio and R. Álvarez-Quintero et al., 2018. Oxidized LDL triggers changes in oxidative stress and inflammatory biomarkers in human macrophages. Redox Biol., 15: 1-11.
CrossRefDirect Link - Kim, E.J., B.H. Kim, H.S. Seo, Y.J. Lee, H.H. Kim, H.H. Son and M.H. Choi, 2014. Cholesterol-induced non-alcoholic fatty liver disease and atherosclerosis aggravated by systemic inflammation. PLoS ONE.
CrossRefDirect Link - Mahmoud, A.M., M.B. Ashour, A. Abdel-Moneim and O.M. Ahmed, 2012. Hesperidin and naringin attenuate hyperglycemia-mediated oxidative stress and proinflammatory cytokine production in high fat fed/streptozotocin-induced type 2 diabetic rats. J. Diabetes Complications, 26: 483-490.
CrossRefDirect Link - Germoush, M.O., H.A. Elgebaly, S. Hassan, E.M. Kamel, M. Bin-Jumah and A.M. Mahmoud, 2020. Consumption of terpenoids-rich Padina pavonia extract attenuates hyperglycemia, insulin resistance and oxidative stress and upregulates PPARγ in a rat model of type 2 diabetes. Antioxidants, Vol. 9, No. 1.
CrossRefDirect Link - Elsayed, R.H., E.M. Kamel, A.M. Mahmoud, A.A. El-Bassuony, M. Bin-Jumah, A.M. Lamsabhi and S.A. Ahmed, 2020. Rumex dentatus L. phenolics ameliorate hyperglycemia by modulating hepatic key enzymes of carbohydrate metabolism, oxidative stress and PPARγ in diabetic rats. Food Chem. Toxicol., Vol. 138.
CrossRefDirect Link - Hassanein, E.H.M., A.M. Sayed, O.E. Hussein and A.M. Mahmoud, 2020. Coumarins as modulators of the keap1/Nrf2/ARE signaling pathway. Oxid. Med. Cell. Longevity.
CrossRefDirect Link - Khalil, H.S.A., N.K. Sedky, K.M. Amin, O.M.A. Elhafez and R.K. Arafa, 2019. Visnagin and benzofuran scaffold-based molecules as selective cyclooxygenase-2 inhibitors with anti-inflammatory and analgesic properties: design, synthesis and molecular docking. Future Med. Chem., 11: 659-676.
CrossRefDirect Link - Lee, J.K., J.S. Jung, S.H. Park, S.H. Park and Y.B. Sim et al., 2010. Anti-inflammatory effect of visnagin in lipopolysaccharide-stimulated BV-2 microglial cells. Arch. Pharm. Res., 33: 1843-1850.
CrossRefDirect Link - Pasari, L.P., A. Khurana, P. Anchi, M.A. Saifi, S. Annaldas and C. Godugu, 2019. Visnagin attenuates acute pancreatitis via Nrf2/NFκB pathway and abrogates associated multiple organ dysfunction. Biomed. Pharmacother.
CrossRefDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Beutler, E., O. Duron and B.M. Kelly, 1963. Improved method for the determination of blood glutathione. J. Lab. Clin. Med., 61: 882-888.
PubMedDirect Link - Marklund, S. and G. Marklund, 1974. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem., 47: 469-474.
CrossRefPubMedDirect Link - Cohen, G., D. Dembiec and J. Marcus, 1970. Measurement of catalase activity in tissue extracts. Ann. Biochem., 34: 30-38.
CrossRefPubMedDirect Link - Kim, C.Y., K.S. Chung, S.Y. Cheon, K. Lee and I. Ham et al., 2016. Hypolipidemic effects of HVC1 in a high cholesterol diet-induced rat model of hyperlipidemia. Mol. Med. Rep., 14: 3152-3158.
CrossRefDirect Link - Livak, K.J. and T.D. Schmittgen, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25: 402-408.
CrossRefDirect Link - Shi, J., R. Li, Y. Liu, H. Lu, L. Yu and F. Zhang, 2019. Shuangyu tiaozhi granule attenuates hypercholesterolemia through the reduction of cholesterol synthesis in rat fed a high cholesterol diet. BioMed. Res. Int.
CrossRefDirect Link - Oates, J.A., A.J.J. Wood and S.M. Grundy, 1988. HMG-CoA reductase inhibitors for treatment of hypercholesterolemia. N. Engl. J. Med., 319: 24-33.
CrossRefDirect Link - Min, H.K., A. Kapoor, M. Fuchs, F. Mirshahi, H. Zhou et al., 2012. Increased hepatic synthesis and dysregulation of cholesterol metabolism is associated with the severity of nonalcoholic fatty liver disease. Cell Metab., 15: 665-674.
CrossRefDirect Link - Zelcer, N., C. Hong, R. Boyadjian and P. Tontonoz, 2009. LXR regulates cholesterol uptake through idol-dependent ubiquitination of the LDL receptor. Sci., 325: 100-104.
CrossRefDirect Link - McGill, M.R., 2016. The past and present of serum aminotransferases and the future of liver injury biomarkers. EXCLI J., 15: 817-828.
CrossRefDirect Link - Asnani, A., B. Zheng, Y. Liu, Y. Wang and H.H. Chen et al., 2018. Highly potent visnagin derivatives inhibit Cyp1 and prevent doxorubicin cardiotoxicity. JCI Insight.
CrossRefDirect Link - Yang, R.L., Y.H. Shi, G. Hao, W. Li and G.W. Le, 2008. Increasing oxidative stress with progressive hyperlipidemia in human: Relation between malondialdehyde and atherogenic index. J. Clin. Biochem. Nutr., 43: 154-158.
CrossRefDirect Link - Smathers, R.L., J.J. Galligan, B.J. Stewart and D.R. Petersen, 2011. Overview of lipid peroxidation products and hepatic protein modification in alcoholic liver disease. Chem. Biol. Interact., 192: 107-112.
CrossRefDirect Link