
Abdulaziz S. Saeedan
Department of Pharmacology, College of Pharmacy, Prince Sattam Bin Abdulaziz University, P.O. Box 173, Al-Kharj, 11942, Saudi Arabia
Majid Ahmad Ganaie
Department of Pharmacology, College of Pharmacy, Prince Sattam Bin Abdulaziz University, P.O. Box 173, Al-Kharj, 11942, Saudi Arabia
LiveDNA: 91.4319
Basit Latief Jan
Department of Clinical Pharmacy, College of Pharmacy, King Saud University, Riyadh, Saudi Arabia
Hassan Madhkali
Department of Pharmacology, College of Pharmacy, Prince Sattam Bin Abdulaziz University, P.O. Box 173, Al-Kharj, 11942, Saudi Arabia
Mohd Nazam Ansari
Department of Pharmacology, College of Pharmacy, Prince Sattam Bin Abdulaziz University, P.O. Box 173, Al-Kharj, 11942, Saudi Arabia
Najeeb U. Rehman
Department of Pharmacology, College of Pharmacy, Prince Sattam Bin Abdulaziz University, P.O. Box 173, Al-Kharj, 11942, Saudi Arabia
Summya Rashid
Department of Pharmacology, College of Pharmacy, Prince Sattam Bin Abdulaziz University, P.O. Box 173, Al-Kharj, 11942, Saudi Arabia
Muneeb U. Rehman
Department of Clinical Pharmacy, College of Pharmacy, King Saud University, Riyadh, Saudi Arabia
International Journal of Pharmacology
Year: 2020 | Volume: 16 | Issue: 4 | Page No.: 319-329







ABSTRACT
Background and Objectives: Brucine is one of the abundantly found alkaloid constituent of semen strychnine, it has amazing pharmacological profile and is known to be helpful in preventing many health ailments. Our study is designed to demonstrate the beneficial role of Brucine in experimental colon carcinogenesis and exploring the possible mechanisms involved e.g., Nrf-2 cascade and downstream inflammatory pathways involved. Materials and Methods: For experimental set up animals were allocated to four groups with six animals in each group (Group I-IV). Control group (group I) was given the vehicle orally. Group II, III and IV were given 1,2, dimethylhydrazine at the dose rate of 20 mg kg–1 b.wt. Group III and IV were treated with Brucine at a dose rate of 2 and 4 mg kg–1 b.wt., respectively continuous for first five weeks and animals were euthanized after 16 weeks. Results: It observed an increase in the level of MDA and ROS, activity of cytochrome P450-2E1 and serum marker enzyme carcinoembryonic antigen (CEA) were also elevated, inflammatory and proliferative proteins showed increased expression. Nrf-2 and NF-κB were downregulated by DMH treatment. Brucine treatment resulted in restoration of activity of the CEA and cytochrome P450-2E1. Expression of inflammatory (Cox-2, i-NoS, IL-6, TNF-α) and proliferative markers (PCNA, Ki-67) were also suppressed by Brucine. Brucine showed great potential in preventing mucosal damage. Conclusion: Considering all results from our study Brucine could be considered as an excellent chemo-preventive in chemically induced colon malignancies. More in depth studies are required to investigate the role of Brucine in modulating other pathways involved in colon malignancies.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Abdulaziz S. Saeedan, Majid Ahmad Ganaie, Basit Latief Jan, Hassan Madhkali, Mohd Nazam Ansari, Najeeb U. Rehman, Summya Rashid and Muneeb U. Rehman, 2020. Brucine Prevents DMH Induced Colon Carcinogenesis in Wistar Rats. International Journal of Pharmacology, 16: 319-329.
DOI: 10.3923/ijp.2020.319.329
URL: https://scialert.net/abstract/?doi=ijp.2020.319.329
DOI: 10.3923/ijp.2020.319.329
URL: https://scialert.net/abstract/?doi=ijp.2020.319.329
INTRODUCTION
Colon cancer is reported to be one of the leading causes of death in the US and second largest in the western world. The frequency of development of colon cancer is highest of all the forms of gastrointestinal malignancies1. Colorectal cancer has been marked as “silent disease” as it is asymptomatic till the time it becomes incurable and resulting in increased deaths. Morphologically colon cancers are represented as an outgrowth of epithelial lining of colon or rectum called as polyps, which may be either adenomatous mostly cancerous or hyperplastic and inflammatory polyps which are usually benign2.
Various factors are involved in the development of colon cancer. One of the factors being oxidative stress, wherein there is a decrease in endogenous antioxidant levels of body and increased production of free radicals. This stress is responsible for inducing cell injury and apoptotic death of cells because of increased oxidation of lipids, proteins and DNA damage. Studies found that there is upregulation of oxidative stress markers like Reactive Oxygen Species (ROS), glutathione peroxidase (GPx), catalase (CAT), nitric oxide (NO), lipid peroxidation (LPO), oxidation of DNA in colon cancer cells and downregulation of cytosine methylation in DNA3. The ROS along with inflammation have been found to be active players in the growth and expansion of colon cancers4. Inflammatory mediators such as; cytokines act as driving force for the production of ROS via interfering with the electron transport chain and resulting in an increased radical outflow from the respiratory chain, these free radicals results in mutations in DNA and formation of cancerous tissues5. The increased production of ROS results in the instigation of transcription machinery which include redox sensitive transcription factors which cause induction of various molecules involved in tumor promotion and inflammation as a result of which variety of inflammation markers which include chemokines (MCP-1 and cxcl12 cytokines and tumor necrosis factor-α and adhesion proteins like e-cadherin and p-cadherin are activated6. The activation of the inflammatory factors results in triggering of downstream signaling pathways which include NFKB, AP-1. Henceforth downstream signaling upregulates proliferative cell signaling, resisting cell death mechanisms, invasion and migration. Thereby, resulting in the formation of tumor and further metastasizing it7.
Previous epidemiological based research has established the importance of continuing inflammation in various types of cancer. Also, the number of deaths resulting from several types of tumors has been reported to get reduced due to use of non-steroidal anti-inflammatory drugs (NSAID’s), which strongly is an evidence for the contribution of inflammation in developing cancer. In recent past evidences from different studies reveal the integral role of inflammatory mediators like; cytokines, interleukins in the development of cancer4,8,9.
Colon carcinogenesis is the uncontrolled proliferation of cells in colon, wherein there is an increase in proliferative protein signaling such as; Proliferating Cell Nuclear Antigen (PCNA) and Ki-6710, which are markers of hyper-proliferation, hence, infer the progress of the disease. PCNA is a 36 KDa co-factor of DNA polymerase- δ, downstream to MAPK/ERK1/2 signaling get activated in chronic proliferative signaling11,12, whereas, Ki-67 is a non-histone nuclear protein which is upregulated in malignant disorders like colorectal cancer13,14.
The serum carcinoembryonic antigen (CEA), a glycoprotein found in fetal and colon adenocarcinoma, acts as ligand in metastasis of cancer15. It is one of the most commonly used biomarkers for detection of incidence of liver cancer, treatment response, survival and recurrence in asymptomatic patients with colorectal malignancies. The level of CEA is several folds increased in tumor cells than normal cells16.
The treatment of colorectal cancer via chemotherapy increases the risk of toxic effects and economic burden for the patients. Thus, natural compounds are used in combination therapy for chemoprevention of cancers, involving co-administration of either conventional chemotherapeutic agent and a natural product or combination of different natural compounds together or as individual compound. Brucine is a natural compound, chemically belonging to indole alkaloids obtained from seeds of Strychnos nux-vomica L. (Loganiaceae). The dried seeds of nux vomica are used in traditional medicinal system in China for its anti-inflammatory, anti-rheumatic and anti-cancer properties17. The compound possesses various pharmacological activities such as; anti-mutagenic and pro-apoptotic on human hepatoma cells (HepG2) and SMMC-722118,19, anti-inflammatory potential against formaldehyde induced rat paw edema, protective effect in liver injury by CCl4 in rats20, antioxidant and antibacterial21. The current study was done to explore the role of Brucine keeping in view the cytotoxic, anti-proliferative and hepato-protective effect in various colorectal cancers by using dimethylhydrazine induced colon cancer in Wistar rats.
MATERIALS AND METHODS
The study was carried out at Pharmacology Research Laboratory, College of Pharmacy, Prince Sattam Bin Abdulaziz University, Al-Kharj, Saudi Arabia. This research project was conducted from 2017-2019.
Chemicals: Bovine Serum Albumin (BSA), EDTA, poly-L-lysine, tris-base, tris-HCl, NADPH, Mayer’s hematoxylin, Alcian blue, toluidine blue, propidium iodide, methylene blue 1,2-dimethylhydrazine (DMH), Brucine were obtained from Sigma (Sigma Chemical Co., St Louis, MO). Poly-HRP plus ONE detection system (Thermo Scientific). All other chemicals used were of highest purity grade available.
Animal study: Four to six-weeks-old, male albino rats (120–150 g) of Wistar strain were obtained from Experimental Animal Care Center, College of Pharmacy, Prince Sattam Bin Abdulaziz University, Alkharj, Saudi Arabia. Animals were maintained under standard laboratory conditions of relative humidity (45-55%), temperature (23-25°C) and light (12 h light/12 h dark), with free access to standard pellet diet and tap water throughout the experimental period. All experimental protocols described in the manuscript were duly approved by the Institutional Animal Care were conducted in accordance with NIH guidelines.
Preparation of carcinogen: The DMH was dissolved in distilled water containing 1 mM EDTA to ensure the stability of the chemical just prior to use and the pH adjusted to 6.5 with 1 M NaOH solution.
Experimental design of the medium term study
Group I (vehicle treated group): In this group, rats will receive basal diet and water ad libitum along with distilled water orally and EDTA solution (vehicle of DMH) subcutaneously.
Group II (1,2-dimethylhydrazine (DMH) treated group): In this group, rats will be treated with DMH (dissolved in 1 mM EDTA solution pH adjusted to 6.5 with 1 mM NaOH) once a week for 5 weeks at the dose of 20 mg kg–1 b.wt., subcutaneously in the groin.
Group III (Brucine and 1,2-dimethylhydrazine (DMH) treated group (prevention group I)): In this group, rats will be first treated with Brucine (2 mg kg–1 b.wt.,) 2 weeks before the DMH treatment (as in group II) and Brucine will be continued till the termination of the experiment.
Group IV (Brucine and 1,2-dimethylhydrazine (DMH) treated group (prevention group II)): In this group, rats will be first treated with Brucine (4 mg kg–1 b.wt.,) 2 weeks before the DMH treatment (as in group II) and Brucine will be continued till the termination of the experiment.
All the rats were sacrificed after 16 weeks and were evaluated for the colon toxicity and early markers of colon carcinogenesis i.e., ROS measurement, CEA, inflammatory and proliferative markers were studied.
Measurement of ROS: The ROS was measured based on the oxidation of 2’7’-dichlorodihydrofluorescein diacetate to 2’7’-dichloro-fluorescein as described by Liu et al.22.
Assay for cytochrome P4502E1: Cytochrome P4502E1 activity was measured by the method of Watt et al.23. The assay mixture contained 40 mM p-nitrophenol, 100 μg microsomal protein and 0.1 M phosphate buffer. The Addition of 10 mM of NADPH helped in the initiation of reaction, the reaction mixture was then incubated at 37°C for 60 min 20% TCA was added to stop the reaction, it was then centrifuged at 1000 rpm for 5 min, 10 mM NaOH was added to the supernatant and the absorbance was measured at 450 nm. The values are expressed as mmoles of p-nitrocatechol liberated/min/mg protein.
Measurement of MDA: The reaction mixture consisted of 0.58 mL phosphate buffer (0.1 M, pH 7.4), 0.2 mL PMS, 0.2 mL ascorbic acid (100 mM) and 0.02 mL ferric chloride (100 mM), in a total volume of 1 mL. This reaction mixture was then incubated at 37°C, in a shaking water bath for 1 h. The reaction was stopped by the addition of 1 mL trichloroacetic acid (10%). Following the addition of 1.0 mL thiobarbituric acid (TBA) (0.67%), all the tubes were placed in a boiling water bath for a period of 20 min. The tubes were shifted to an ice bath and then centrifuged at 2500 g for 10 min. The amount of malondialdehyde (MDA) formed in each of the samples was assessed by measuring the optical density of the supernatant at 535 nm. The results were expressed as nmol TBA formed/h g–1 tissue at 37°C by using a molar extinction coefficient of 1.56×105/M cm–1.
Immunohistochemical staining NF-κB-p65, Nrf-2, Ki-67, PCNA: The processed colon tissues were obtained and preserved in 10% paraformaldehyde overnight followed by dehydration in 30, 20 and 10% sucrose solution successively up to 3 days and were then fixed in formaldehyde fixative until immunochemical staining. Then, 5-15 μm-thick sections of paraffin embedded tissues were cut by using microtome and boiled in 0.1 M-citrate buffer (pH 6.0) for 5 min for the antigen retrieval process and then incubated in 0.3% H2O2 in methanol followed by incubation in blocking buffer containing 0.1 M-PBS, 0.04% Triton X-100 and 10% normal goat serum. Tissue sections were stained with antibodies against anti-rat NF-κB-p65 rabbit antibody (dilution 1:100), anti-rat Nrf-2 rabbit antibody (dilution 1:100), anti-rat Ki-67 polyclonal antibody (dilution 1:200) and anti-rat PCNA polyclonal antibody (dilution 1:200) overnight at 48°C. After rinsing in the buffer, the sections were processed using a three-layer peroxidase staining kit. The peroxide complex was visualised with 3,3-diaminobenzidine. Lastly, the slides were counterstained with haematoxylin for 5 sec. The slides were then cleaned in sterile HPLC-grade water, gradually dehydrated with ethanol, cover slipped in mounting medium and photographed under microscope.
Assay for carcinoembryonic antigen (CEA): The activity of carcinoembryonic antigen (CEA) enzyme levels in serum was measured by ARCHITECT CEA enzyme chemiluminescent microparticle immunoassay test kit (Abbott, Ireland Diagnostic Division, Sligo, Ireland) as per the manufacturer protocol.
Assay for tumor necrosis factor-alpha (TNF-α): TNF-α levels were determined by rat TNF-α kit (eBioscience, Inc., San Diego., USA). The method is based on enzyme-linked immunosorbent assay (ELISA). We have performed measurement of TNF-α in the colonic tissue by ELISA. Samples were prepared in Phosphate Buffered Saline (PBS) containing protease inhibitor cocktail. Analysis was performed by ELISA Plate Reader (Multiskan EX, Thermo) according to the manufacturer’s instruction.
Assay for interleukin 6 (IL-6): IL-6 was assayed by rat IL-6 kit (eBioscience, Inc., San Diego., USA). The method is based on enzyme-linked immunosorbent assay (ELISA). We have performed measurement of IL-6 in the colonic tissue by ELISA. Samples were prepared in Phosphate Buffered Saline (PBS) containing protease inhibitor cocktail. Analysis was performed by Elisa Plate Reader (Multiskan EX, Thermo) according to the manufacturer’s instruction.
Assay for cyclooxygenase-2 (Cox-2): Cox-2 levels were determined by rat Cox-2 kit (eBioscience, Inc., San Diego., USA). The method is based on enzyme-linked immuno-sorbent assay (ELISA). We have performed measurement of Cox-2 in the colonic tissue by ELISA. Samples were prepared in Phosphate Buffered Saline (PBS) containing protease inhibitor cocktail. Analysis was performed by ELISA Plate Reader (Multiskan EX, Thermo) according to the manufacturer’s instruction.
Assay for inducible-nitrogen oxide synthesis (i-NOS): The i-NOS levels were determined by rat Cox-2 kit (eBioscience, Inc., San Diego., USA). The method is based on enzyme-linked immuno-sorbent assay (ELISA). We have performed measurement of i-NOS in the colonic tissue by ELISA. Samples were prepared in Phosphate Buffered Saline (PBS) containing protease inhibitor cocktail. Analysis was performed by ELISA Plate Reader (Multiskan EX, Thermo) according to the manufacturer’s instruction.
Statistical analysis: The data from individual groups were presented as the mean±Standard Error of the Mean (SEM). Differences between groups were analyzed by using analysis of variance (ANOVA) followed by Tukey-Kramer multiple comparisons test and minimum criterion for statistical significance was set at p<0.05 for all comparisons.
RESULTS
Effect of Brucine on MDA levels: Figure 1 shows the data on the effect of Brucine on the levels of lipid peroxidation of control and experimental rats. In DMH alone treated rats (group II) the levels of were significantly (***p<0.001) increased in the colon as compared to the control (group I). On the other hand, Brucine supplementation (group III and IV) to DMH treated rats during the experimental period of 16 weeks restored (##p<0.01, ###p<0.001) the levels of lipid peroxidation to near those of control (Fig. 1).
Effect of DMH and Brucine on phase-I xenobiotic metabolizing enzymes (microsomes): The activities of phase-I enzymes were significantly increased (***p<0.001) on DMH treatment.
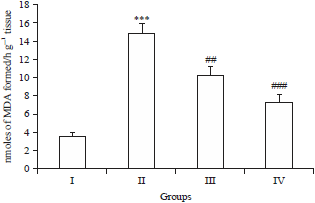 | |
| Fig. 1: | Effects of Brucine and DMH on MDA levels |
In DMH treated group, the MDA level was increased significantly (***p<0.001) as compared to control group I, Brucine administration significantly attenuated the MDA level in group III (##p<0.01) and group IV (###p<0.001) as compared to group II | |
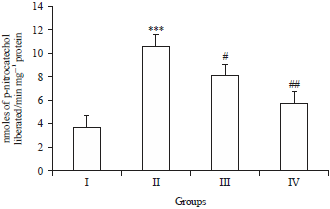 | |
| Fig. 2: | Effects of Brucine on CYP450 2E1 levels in DMH induced colon carcinogenesis |
In DMH-treated group, the CYP450 2E1 level was increased significantly (***p<0.001) as compared to control group I, while, treatment with Brucine significantly attenuated the CYP450 2E1 level in group III (#p<0.05) and group IV (##p<0.01) as compared to group II | |
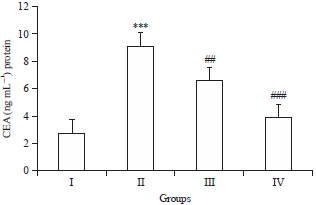 | |
| Fig. 3: | Effects of Brucine on CEA levels in DMH induced colon carcinogenesis |
In DMH-treated group, the CEA level was increased significantly (***p<0.001) as compared to control group I, treatment with Brucine significantly attenuated the CEA level in group III (##p<0.01) and group IV (###p<0.001) as compared to group II | |
Brucine regulates the activity of xenobiotic metabolizing enzymes of phase I, cytochrome P4502E1 in the colon mucosa of experimental and control animals. Supplementation with different doses of Brucine significantly (#p<0.05, ##p<0.01) diminished the activity of cytochrome P4502E1. A more pronounced effect being observed in the rats supplemented with Brucine at 2 and 4 mg kg–1 b.wt., (Fig. 2).
Brucine regulates carcinoembryonic antigen (CEA): There was marked rise in level of CEA in DMH-treated group II (***p<0.001) compared to control. Following DMH induction, Brucine treatment showed significant (##p<0.01, ###p<0.001) decline in CEA levels as compared to control group I (Fig. 3).
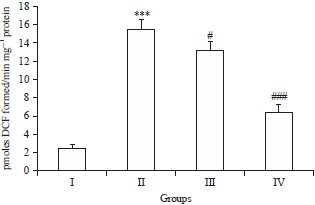 | |
| Fig. 4: | Effects of Brucine and DMH on ROS levels |
In DMH treated group, the ROS level was increased significantly (***p<0.001) as compared to control group I, treatment with Brucine significantly attenuated the ROS level in group III (#p<0.05) and group IV (###p<0.001) as compared to group II | |
Effect of Brucine on ROS levels in colonic tissue: The ROS level significantly increased (***p<0.001) in DMH treated group (group II) as compared with vehicle control (group I). The data shows that dimethyl hydrazine treatment causes oxidative stress by enhancing reactive oxygen species production in the rat’s colon. Brucine can lower the ROS levels in the colon of both group III (#p<0.05) and group IV (###p<0.001) (Fig. 4).
Effect of Brucine on inflammatory signaling: Immuno-histochemistry patterns of NF-κB and Nrf-2 protein expression in the tissues of colon showed intense immunoreactivity in DMH treated rats (group II) than in control (group I) (Fig. 5, 6). Visible reduction in the expression of all these redox sensitive inflammatory markers NF-κB and Nrf-2 were observed in the groups treated with Brucine (group III and IV) compared to DMH only treated group (Fig. 5, 6). There was a steep increase in level of Cox-2 and i-NOS in DMH treated group II. Also, TNF-α and IL-6 levels were found to be significantly (***p<0.001) higher as compared to group I control. Treatment with Brucine at both the doses markedly reduced the level of all inflammatory markers (Table 1). Hence, it proposed that Brucine can inhibit inflammatory signaling by regulating the above proteins.
Effect of Brucine and DMH on the expression of Ki-67 and PCNA: The Ki-67 and PCNA are well known and reliable markers of hyperproliferation. It observed that DMH treated group (group II) have more Ki-67, PCNA, immunopositive staining as indicated by brown colour as compared to control group (group I). Treatment of Brucine reduced the immunostaining of both hyperproliferative markers studied PCNA and Ki-67, in group III and IV as compared to group II.
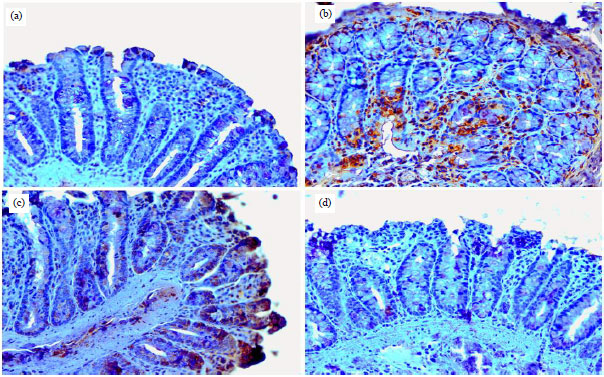 | |
| Fig. 5(a-d): | Photomicrographs depicting immunohistochemical staining of NF-κB. For immunohistochemical analyses, brown colour indicates specific immunostaining of NF-κB and light blue colour indicates haematoxylin staining. The colonic section of DMH-treated group (group II) has more NF-κB, immunopositive staining as indicated by brown colour as compared to control group (group I) while, treatment with Brucine in group III and group IV reduced NF-κB, immunostaining as compared to group II |
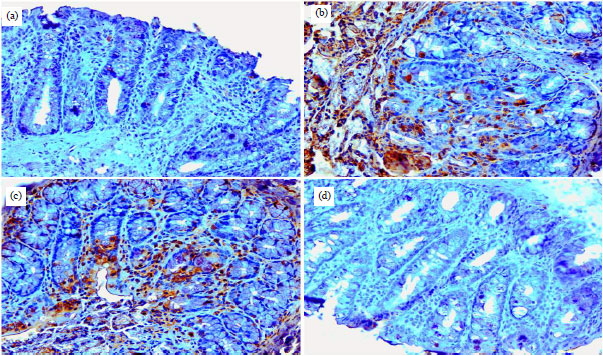 | |
| Fig. 6(a-d): | Photomicrographs depicting immunohistochemical staining of Nrf-2. For immunohistochemical analyses, brown colour indicates specific immunostaining of Nrf2 and light blue colour indicates nuclear haematoxylin staining. The colonic section of DMH treated group (group II) has less Nrf2 immunopositive staining as indicated by no or very less blue colour as compared to control group (group I) while treatment with Brucine in group III and IV increased Nrf2 immunostaining as compared to group II |
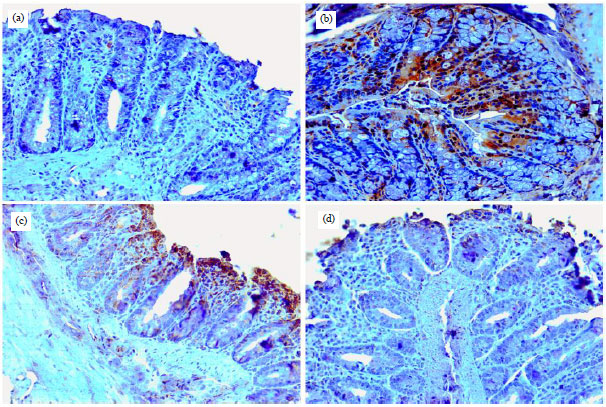 | |
| Fig. 7(a-d): | Photomicrographs depicting immunohistochemical staining of PCNA. For immunohistochemical analyses, brown colour indicates specific immunostaining of PCNA and light blue colour indicates nuclear haematoxylin staining. The colonic section of DMH-treated group (group II) has more PCNA immunopositive staining as indicated by brown colour as compared to control group (group I) while treatment with Brucine in group III and IV reduced PCNA immunostaining as compared to group II |
| Table 1: | Effect of Brucine on serum levels of inflammatory cytokines (IL-6, TNF-α, i-NOS and Cox-2) |
 | |
Results obtained are significantly different from group I (***p<0.001), results obtained are significantly different from group II (#p<0.05, ##p<0.01 and ###p<0.001), IL-6: Interleukin 6, TNF-α: Tumor necrosis factor-alpha, i-NOS: Inducible nitrogen oxide synthesis, Cox-2: Cyclooxygenase-2 | |
For immunohistochemical analyses, brown colour indicates specific immunostaining of PCNA and Ki-67 and light blue color indicates hematoxylin staining (Fig. 7, 8).
DISCUSSION
In present manuscript, it analyzed potential of Brucine on the inflammatory and proliferative pathway in the MHD induced colon cancer. The results from present investigation demonstrate significant changes in inflammatory and proliferative markers in DMH induced colon carcinogenesis in Wistar rats and administration of Brucine showed significant suppression of inflammatory and proliferative markers expression. As per literature search, our study is the first report showed evidences of colon cancer prevention by Brucine via effecting inflammation and proliferation in Wistar rats. Epidemiological reports reveal an opposite relationship between the usage of naturally occurring phytochemicals and colon cancer risk4,24-26. Phytochemicals from plants have huge potential to modulate cellular defense systems including anti-inflammatory and anti-proliferative response. Such defensive mechanism systems of plant compounds are further explored to target vital markers of inflammatory and proliferative cascade27,28. These cellular defensive mechanisms by phytochemicals are being elicited by targeting different transcription factors and other downstream regulatory molecules of inflammatory and proliferative cascade29,30.
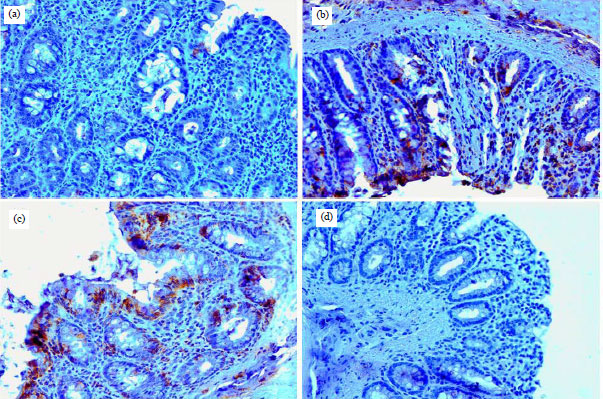 | |
| Fig. 8(a-d): | Photomicrographs depicting immunohistochemical staining of Ki-67: For immunohistochemical analyses, brown colour indicates specific immunostaining of Ki-67 and light blue colour indicates haematoxylin staining. The colonic section of DMH-treated group (group II) has more Ki-67, immunopositive staining as indicated by brown colour as compared to control group (group I) while treatment with Brucine in group III and IV reduced i- Ki-67, immunostaining as compared to group II |
Brucine obtained mainly from seeds Strychnos nuxvomica, is a naturally occurring alkaloid type (dihydroindole-e) have been reported to exhibit many important pharmacological properties e.g., anti-oxidant, anti-inflammatory and anti-mutagenic potential etc31-33.
In our recent report, it found oxidative stress plays main role in DMH induced colon toxicity and carcinogenesis8. The DMH is a pro-carcinogen, gets converted to a methyl free radical, results in the formation of hydrogen peroxide or hydroxyl radical with co-occurrence of metal ions, which might be a contributing factor to peroxidation of lipids and cancer development. Increased levels of malonyl aldehyde, direct indicator of lipid peroxidation have been studied as one of the most significant markers of oxidative injury. The DMH induced elevation of MDA is widely reported4, similar results were obtained in the present study as well. Brucine administration significantly brought back the level of MDAs to normal (Fig. 1) which corresponds to the previous reports of Saraswati et al.31 and Saminathan et al.34.
Sohn et al.35 for the first time reported role of methylazoxymethanol (MAM), which is a major metabolite of DMH and a very strong carcinogen to be activated metabolically by CYP2E1 under in vitro as well as in vivo condition. It observed that Brucine restored the normal activity of CYP2E1 (a phase 1 enzyme) which showed enhanced activity following DMH treatment (Fig. 2) as reported previously36. Bio-activation of DMH occurs by either cytochrome P450 dependent oxidation or by glutathione (GSH) conjugation leads to toxicity. The GSH is a cellular antioxidant, low-molecular-weight tripeptide and is found to be depleted following DMH treatment37,38. Reports suggested that hallmarks of oxidative stress response get malformed after DMH treatment which include decreased reduced glutathione (GSH) and associated redox cycle enzymes including catalase, SOD, GPX and GR38. Brucine is anti-oxidative as per reports31,34,38,39.
Carcinoembryonic antigen (CEA) an adhesion molecule is a typical tumor marker for colon cancer being associated with advanced and recurrent diseases40,41. A member of super-immunoglobulin gene family, which encodes for adhesion proteins, CEA is found in increased levels or upregulated in colon cancers16. Therefore, regulating the level of expression of CEA in colon cancer is a smart way to determine the effectiveness of therapy. The DMH causes immense variations in CEA levels via enhancing ROS induced injury to colon cells16. In the present study, there was an elevation of CEA in DMH induced rats as compared to control group animals and treatment with Brucine ameliorated CEA levels (Fig. 3).
The ROS generation is the hallmark of DMH toxicity, which included singlet oxygen, hydrogen peroxide, superoxide and hydroperoxides4,25. In the present communication, DMH exposure prompted the massive production of ROS, which causes oxidative damage in the colon. Many studies have previously showed that the antioxidant potential or ROS scavenging potential of natural product is one of the mechanisms for its anti-carcinogenic potential in the present colon carcinogenesis rat model. As previously shown supplementation with Naringenin, Zingerone etc., have significantly re-establish the normal levels of ROS in DMH induced colon carconigenesis4,8,9. In accordance with mentioned reports Brucine markedly reversed colon toxicity induced by DMH via decreasing the oxidative stress in rat colon, due to regulation of free radicals (Fig. 4).
Oxidative stress and inflammation are intricately connected via a series of pathways in proliferative disorders25,42. Mast cells are one of the primary cell types authenticated to have a role in inflammation driven proliferation and tumor promotion43,44. Excess infiltration of mast cells is characteristic of gastrointestinal disorders, it is also associated with increased polyp number and promotion of colon carconogensis4,44. Brucine has been documented to have good anti-inflammatory activity31 and in line with this, it reduced permeation of mast cells in experimental models of colon carcinogenesis.
Nuclear factor kappa B is a redox sensitive transcription factor, regulates various biochemical activities like; inflammation differentiation and cellular proliferation. Under normal conditions, NF-κB is attached to cytosolic repressor protein, IκB. External stimuli (oxidative stress, radiations and carcinogens) leads to the degradation and phosphorylation of IκB by cytoplasmic enzyme IκB kinase (IKK), resulting in nuclear translocation of NF-κB where regulation of the transcription of several associated genes and several downstream inflammatory mediators occurs e.g., Cox-2, i-NOS, interleukins and cadherins etc45,46. Carcinogens and chronic inflammation activate NF-κB signaling47-49, which trans-activate downstream pathways including TNF-α, Cox-2, i-NOS and IL-6 etc. NF-κB being a sensitive transcription factor has been reported to get activated on exposure to oxidants generated by DMH4,9. Inconsistent with previous papers strong positive signal of NF-κB in colon tissue was observed after
DMH treatment (Fig. 5). It observed Brucine inhibited NF-κB expression and downstream molecules (Table 1) in the colon, which may be a plausible mechanism of inhibition of colon carcinogenesis.
Cellular proliferation acts an important player in cancer progression and has been documented to have a direct role in the pathogenesis of colon carcinogenesis, it is also consistent indicators in the gauging kinetics of cell proliferation in tumor50,51. One of the most efficient markers of cellular proliferation is proliferating cell nuclear antigen (PCNA), produced between late G1 and S phase and is an ancillary protein of DNA polymerase δ52. Ki-67 is another known protein marker in a nucleus which is upregulated in cells having proliferative signaling state4,53,54 and a few others reported down regulation of PCNA and Ki-67 by the use of natural phytochemicals in colon carcinogenesis induced by DMH8,9. Brucine has also been shown to suppress the hyperproliferative response in different experimental models31,55. In agreement with previous reports, in the current study there was decrease in expression of PCNA and Ki-67 after Brucine treatment (Fig. 7 and 8).
CONCLUSION
The inflammatory signaling controlled by NF-κB and Nrf-2 are the major targets of Brucine, further proliferation regulated by PCNA and Ki-67 is also targeted by Brucine, thus, accounting for its anti-cancer property in animal model of colon cancer. The results are validated by the studies which are going on or have been done previously which support the claims about exceptional property of Brucine. The study highlights the effect and influence of Brucine on some molecular targets in colon cancer induced by DMH. Further research needs to be done to explore their mechanistic insight.
SIGNIFICANCE STATEMENT
Brucine treatment resulted in restoration of activity of the carcinoembryonic antigen and cytochrome P450-2E1. Also, suppression of expression of inflammatory (Cox-2, i-NoS, IL-6 and TNF-α) and proliferative markers (PCNA, Ki-67). Brucine showed promising potential in preventing mucosal damage. Considering all results from our study brucine could be considered as an excellent chemo-preventive agent in chemically induced colon malignancies. More in depth studies are required to investigate role of Brucine in modulating other pathways involved in colon malignancies.
ACKNOWLEDGMENT
This project was supported by the Deanship of Scientific Research at Prince Sattam Bin Abdulaziz University under the research project number 2016/03/6564. The funding source had no role in designing, writing and in the decision to submit the article for publication.
REFERENCES
- Siegel, R., D. Naishadham and A. Jemal, 2012. Cancer statistics, 2012. CA: Cancer J. Clinicians, 62: 10-29.
CrossRefPubMedDirect Link - Dubois R.N., 2014. Role of inflammation and inflammatory mediators in Colorectal Cancer. Trans. Am. Clin. Climatol. Assoc., 125: 358-373.
Direct Link - Sreevalsan S., S. Safe, 2013. Reactive Oxygen Species and Colorectal Cancer. Curr. Colorectal Cancer Rep., 9: 350-357.
CrossRefDirect Link - Rehman, M.U., M.U. Rahman Mir, A. Farooq, S.M. Rashid and B. Ahmad et al., 2018. Naringenin (4,5,7-trihydroxyflavanone) suppresses the development of precancerous lesions via controlling hyperproliferation and inflammation in the colon of Wistar rats. Environ. Toxicol., 33: 422-435.
CrossRefDirect Link - Dalle-Donne, I., R. Rossi, R. Colombo, D. Giustarini and A. Milzani, 2006. Biomarkers of oxidative damage in human disease. Clin. Chem., 52: 601-623.
CrossRefDirect Link - Wongchana W., T. Palaga 2011. Direct regulation of interleukin-6 expression by Notch signaling in macrophages. Cell Mol. Immunol., 9: 155-162.
CrossRefDirect Link - Zheng, W., M. Umitsu, I. Jagan, C.W. Tran and N. Ishiyama et al., 2016. An interaction between Scribble and the NADPH oxidase complex controls M1 macrophage polarization and function. Nat. Cell Biol., 18: 1244-1252.
CrossRefDirect Link - Ganaie, M.A., A. Al Saeedan, H. Madhkali, B.L. Jan and T. Khatlani et al., 2019. Chemopreventive efficacy zingerone (4‐[4‐hydroxy‐3‐methylphenyl] butan‐2‐one) in experimental colon carcinogenesis in Wistar rats. Environ. Toxicol. 34: 610-625.
CrossRefDirect Link - Khan, T.H., M.A. Ganaie, K.M. Alharthy, H. Madkhali, B.L. Jan and I.A. Sheikh, 2018. Naringenin prevents doxorubicin-induced toxicity in kidney tissues by regulating the oxidative and inflammatory insult in wistar rats. Arch. Physiol. Biochem., 8: 1-8.
CrossRefDirect Link - Bostick, R.M., L. Fosdick, T.J. Lillemoe, P. Overn and J.R. Wood et al., 1997. Methodological findings and considerations in measuring colorectal epithelial cell proliferation in humans. Cancer Epidemiol. Biomark., 6: 931-942.
Direct Link - Guzińska-Ustymowicz, K., A. Pryczynicz, A. Kemona and J. Czyzewska, 2009. Correlation between proliferation markers: PCNA, Ki-67, MCM-2 and antiapoptotic protein Bcl-2 in colorectal cancer. Anticancer Res., 29: 3049-3052.
Direct Link - Kelman Z., 2002. PCNA: structure, functions and interactions. Oncogene, 14: 629-640.
CrossRefDirect Link - Lian T.L., J. Guan, C. Qian and J.N. Zheng, 2014. Ki67 is a promising molecular target in the diagnosis of cancer (Review). Mol. Med. Rep., 11: 1566-1572.
CrossRefDirect Link - Hu H.Y., H. Liu, J.W. Zhang, K. Hu and Y. Lin, 2012. Clinical Significance of Smac and Ki-67 Expression in Pancreatic Cancer. Hepatogastroenterol., 59: 2640-2643.
CrossRefDirect Link - Cho, W.K., D.H. Choi, H.C. Park, W. Park and J.I. Yu et al., 2017. Elevated CEA is associated with worse survival in recurrent rectal cancer Oncotarget 8: 105936-105941.
CrossRefDirect Link - Chan C.H.F., P. Camacho-Leal and C.P. Stanners, 2007. Colorectal Hyperplasia and Dysplasia due to Human Carcinoembryonic Antigen (CEA) family member expression in transgenic mice. PLoS ONE.
CrossRefDirect Link - Bhati, R., A. Singh, V.A. Saharan, V. Ram and A. Bhandari, 2012. Strychnos nux-vomica seeds: Pharmacognostical standardization, extraction, and antidiabetic activity. J. Ayurveda Integr. Med., 3: 80-84.
CrossRefDirect Link - Yin, W., X.K. Deng, F.Z. Yin, X.C. Zhang and B.C. Cai, 2007. The cytotoxicity induced by brucine from the seed of Strychnos nux-vomica proceeds via apoptosis and is mediated by cyclooxygenase 2 and caspase 3 in SMMC 7221 cells. Food Chem. Toxicol., 45: 1700-1708.
CrossRefDirect Link - Mitra S., V. Kumar and B.K. Ashok, 2011. A comparative anti-inflammatory activity of raw and processed Kupeelu Strychnos nux-vomica (Linn.) seeds on albino rats. Anc. Sci. Life, 31: 73-75.
Direct Link - Gopalkrishna S.V., N.M. Lakshmi and S.S. Ramachandra, 2010. Hepatoprotective activity of detoxified seeds of nux-vomica against CCl4 induced hepatic injury in albino rats. Pharmacologyonline, 3: 803-815.
Direct Link - Liu, C.M., Y.L. Zheng, J. Lu, Z.F. Zhang, S.H. Fan, D.M. Wu and J.Q. Ma, 2010. Quercetin protects rat liver against lead-induced oxidative stress and apoptosis. Environ. Toxicol. Pharmacol., 29: 158-166.
CrossRefDirect Link - Watt K.C., C.G. Plopper and A.R. Buckpitt, 2002. Measurement of cytochrome P450 2E1 activity in Rat Tracheobronchial airways using High-Performance Liquid Chromatography with electrochemical detection. Anal. Biochem., 248: 26-30.
CrossRefDirect Link - Janati, A.I., I. Karp, H. Sabri and E. Emami, 2019. Is a fusobacterium nucleatum infection in the colon a risk factor for colorectal cancer? a systematic review and meta-analysis protocol. Syst. Rev.,.
CrossRefDirect Link - Rehman, M.U., M. Tahir, A.Q. Khan, R. Khan and A. Lateef et al., 2013. Chrysin suppresses renal carcinogenesis via amelioration of hyperproliferation, oxidative stress and inflammation: plausible role of NF-κB. Toxicol. Lett., 216: 146-158.
CrossRefDirect Link - Birt D.F. and G.J. Phillips, 2013. Diet, Genes, and Microbes. Toxicol. Pathol., 42: 182-188.
CrossRefDirect Link - Trock B., E. Lanza and P. Greenwald, 2002. Dietary fiber, vegetables, and Colon Cancer: critical review and meta-analyses of the epidemiologic evidence. J. Natl. Cancer Inst., 82: 650-661.
CrossRefDirect Link - Hanahan, D. and R.A. Weinberg, 2011. Hallmarks of cancer: The next generation. Cell, 144: 646-674.
CrossRefPubMedDirect Link - Zhu, D., Y. Sun, L. Qi, R. Zhong and X. Miao, 2015. Dietary legume consumption reduces risk of colorectal cancer: evidence from a meta-analysis of cohort studies. Sci. Rep.
CrossRefDirect Link - Aravindaram, K. and N.S. Yang 2010. Anti-inflammatory plant natural products for cancer therapy. Planta Med., 76: 1103-1117.
CrossRefDirect Link - Saraswati S., A.A. Alhaider and S.S. Agrawal, 2013. Anticarcinogenic effect of brucine in diethylnitrosamine initiated and phenobarbital-promoted hepatocarcinogenesis in rats. Chem. Biol. Interact., 206: 214-221.
CrossRefDirect Link - Serasanambati, M., S.R. Chilakapati, P.K. Manikonda, J.R. Kanala and D.R. Chilakapati, 2015. Anticancer effects of brucine and gemcitabine combination in MCF-7 human breast cancer cells. Nat. Prod. Res., 29: 484-490.
CrossRefDirect Link - Guo R., T. Wang, G. Zhou, M. Xu and X. Yu et al., 2018. Botany, Phytochemistry, Pharmacology and Toxicity of Strychnos nux-vomica L.: A Review. Am. J. Chin. Med. 46: 1-23.
CrossRefDirect Link - Saminathan U., P. Pugalendhi, S. Subramaniyan and R. Jayaganesh, 2018. Biochemical studies evaluating the chemopreventive potential of brucine in chemically induced mammary carcinogenesis of rats. Toxicol. Mech. Methods, 29: 8-17.
CrossRefDirect Link - Sohn, O.S., H. Ishizaki, C.S. Yang and E.S. Fiala, 1991. Metabolism of azoxymethane, methylazoxymethanol and N-nitrosodimethylamine by cytochrome P45011El. Carcinogenesis, 12: 127-131.
CrossRefDirect Link - Chihara T., K. Shimpo, T. Kaneko, H. Beppu, A. Tomatsu and S. Sonoda, 2009. Inhibitory effects of high temperature- and pressure-treated garlic on formation of 1,2-dimethylhydrazine-induced mucin-depleted foci and O(6)-methylguanine DNA adducts in the rat colorectum. Asian Pac. J. Cancer Prev., 10: 827-831.
Direct Link - Rajeshkumar N.V. and R. Kuttan, 2003. Modulation of carcinogenic response and antioxidant enzymes of rats administered with 1,2-dimethylhydrazine by Picroliv. Cancer Letters, 191: 137-143.
CrossRefDirect Link - Sengottuvelan, M., R. Senthilkumar and N. Nalini, 2006. Modulatory influence of dietary resveratrol during different phases of 1,2-dimethylhydrazine induced mucosal lipid-peroxidation, antioxidant status and aberrant crypt foci development in rat colon carcinogenesis. Biochim. Biophys. Acta, 1760: 1175-1183.
CrossRefPubMedDirect Link - Tripathi, Y.B. and S. Chaurasia, 2000. Interaction of Strychnos nux-vomica-products and iron: With reference to lipid peroxidation. Phytomedicine, 7: 523-528.
CrossRefDirect Link - Duffy, M.J., 2001. Carcinoembryonic antigen as a marker for colorectal cancer: Is it clinically useful? Clin. Chem., 47: 624-630.
Direct Link - Das, V., J. Kalita and M. Pal, 2017. Predictive and prognostic biomarkers in colorectal cancer: A systematic review of recent advances and challenges. Biomed. Pharmacother., 87: 8-19.
CrossRefDirect Link - Bickers, D.R. and M. Athar, 2006. Oxidative stress in the pathogenesis of skin disease. J. Invest. Dermatol., 126: 2565-2575.
CrossRefPubMedDirect Link - Gounaris E., S.E. Erdman, C. Restaino, M.F. Gurish, D.S. Friend et al., 2007. Mast cells are an essential hematopoietic component for polyp development. Proc. Natl. Acad. Sci., 104: 19977-19982.
CrossRefDirect Link - Heijmans J., N.V. Büller, V. Muncan and G.R. van den Brink, 2010. Role of mast cells in colorectal cancer development, the jury is still out. Biochimica et Biophysica Acta (BBA) - Mol. Basis Dis., 1822: 9-13.
CrossRefDirect Link - Karin, M. and F.R. Greten, 2005. NF-kB: Linking inflammation and immunity to cancer development and progression. Nat. Rev. Immunol., 5: 749-759.
CrossRefDirect Link - Hatanaka N., T. Seki, J.I. Inoue, A. Tero and T. Suzuki, 2018. Critical roles of IκBα and RelA phosphorylation in transitional oscillation in NF-κB signaling module. J. Theor. Biol., 462: 479-489.
CrossRefDirect Link - Pawlowska, E., J. Szczepanska, K. Wisniewski, P. Tokarz, D. Jaskólski and J. Blasiak, 2018. NF-κB-Mediated Inflammation in the Pathogenesis of Intracranial Aneurysm and Subarachnoid Hemorrhage. Does Autophagy Play a Role? Int. J. Mol. Sci.,.
CrossRefDirect Link - Gupta S.C., A.B. Kunnumakkara, S. Aggarwal and B.B. Aggarwal, 2018. Inflammation, a Double-Edge Sword for Cancer and Other Age-Related Diseases. Front. Immunol.
CrossRefDirect Link - Dolcet X., D. Llobet, J. Pallares and X. Matias-Guiu, 2005. NF-kB in development and progression of human cancer. Virchows Arch., 446: 475-482.
CrossRefDirect Link - Xu, G., G. Ren, X. Xu, H. Yuan and Z. Wang, 2009. Combination of curcumin and green tea catechins prevents dimethylhydrazine-induced colon carcinogenesis. Food Chem. Toxicol., 48: 390-395.
CrossRefDirect Link - Mori H., K. Hata, Y. Yamada, T. Kuno, and A. Hara, 2005. Significance and role of early-lesions in experimental colorectal carcinogenesis. Chem. Biol. Interact., 155: 1-9.
CrossRefDirect Link - Takasaki Y., J.S. Deng and E.M. Tan, 2004. A nuclear antigen associated with cell proliferation and blast transformation. J. Exp. Med., 154: 1899-1909.
CrossRefDirect Link - Scholzen, T. and J. Gerdes, 2000. The Ki-67 protein: From the known and the unknown. J. Cell. Physiol., 182: 311-322.
CrossRefDirect Link - Sahin A.A., J.Y. Ro and R.W. Brown, 1994. Assessment of Ki-67 derived tumor proliferative activity in colorectal adenocarcinomas. Modern Pathol., 7: 17-22.
Direct Link - Tang M., W.J. Zhu, Z.C. Yang and C.S. He, 2019. Brucine inhibits TNF‑α‑induced HFLS‑RA cell proliferation by activating the JNK signaling pathway. Exp. Ther. Med., 18: 735-740.
CrossRefDirect Link