
Gamal A. Gabr
Department of Pharmacology and Toxicology, College of Pharmacy, Prince Sattam bin Abdulaziz University, Al-Kharj, 11942, Saudi Arabia
LiveDNA: 20.32404
Salwa M. El-Sayed
Department of Biochemistry, Faculty of Agriculture, Ain Shams University, Cairo, Egypt
Hossam M. Abd El-Hameed
Food Technology Research Institute, Agricultural Research Center, Giza, Egypt
International Journal of Pharmacology
Year: 2020 | Volume: 16 | Issue: 4 | Page No.: 282-290







ABSTRACT
Background and Objective: Celiac disease in genetically susceptible in individuals. It is resulted from T cell intolerance to wheat gluten that is rich with proline residue and are highly resistant to degradation by the proteolytic enzymes secreted by the gastrointestinal tract. The aim of the current study was to purify and identify new prolyl endopeptidase from natural and economical source such as larva midgut of red palm weevil (Rhynchophorus ferrugineus) able to degrade gluten at low gastric pH. Materials and Methods: The prolyl endopeptidase was extracted from larva midgut of red palm weevil (Rhynchophorus ferrugineus) and purified using ammonium sulphate, acetone and trichloroacetic acid. The enzyme activity was tested of hydrolysis efficacy by gluten, gliadin and peptide peptidyl-pNA. Results: The results showed that 80% acetone give high enzyme activity, high Vmax value and low Km value. At 2 mg mL–1, the efficacy to hydrolyze both gluten and gliadin were found higher than digestine, a positive control. The optimum enzyme activity was at pH 5 and 30°C, the stability of enzyme activity was remaining at temperature reached to 50°C and pH 4.0. Conclusion: The results reflect the potential use of the prolyl endopeptidase in the medical and processing of gluten free food applications.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Gamal A. Gabr, Salwa M. El-Sayed and Hossam M. Abd El-Hameed, 2020. Novel Prolyl-endopeptidase from Rhynchophorus ferrugineus of Gluten-degrading: Potential Use to Reduce Gluten Immunogenic Peptides in Celiac Disease. International Journal of Pharmacology, 16: 282-290.
DOI: 10.3923/ijp.2020.282.290
URL: https://scialert.net/abstract/?doi=ijp.2020.282.290
DOI: 10.3923/ijp.2020.282.290
URL: https://scialert.net/abstract/?doi=ijp.2020.282.290
INTRODUCTION
Celiac Disease (CD) is the most common form of gluten intolerance1. It is an autoimmune syndrome, the immune system attacks the gluten and gut lining causing gut wall damage, severe anemia and nutrient deficiencies2,3. The symptoms of CD are including difficulty in digestion, damaging in the tissue of small intestine, diarrhea, headache, constipation, bloating, tiredness, depression, skin rashes, bad-smelling feces and weight loss4. Gluten, a protein present in all cereals mainly barley, wheat and rye are mainly composed of gliadin and glutenin5. Gliadins is the protein responsible for development of CD6. The high contents of glutamine and proline residues in gluten lead to difficulty to digest gluten by gastric hydrolases, therefore, the gluten disorders such as CD, wheat allergy and baker’s asthma are created7–9. The available therapies for the management of CD is only the avoid intake of dietary gluten, therefore, multiple studies reported many patients when they intake Gluten Free Diet (GFD) for long time lead to improves the biochemical blood parameters and other chronic pathologies such as anemia, osteoporosis, infertility, depression, mortality, growth failure and the risk of other complications related to CD10,11. Therefore, there is urgent need to find the alternative solution such as; enzymes either to gluten degrade or as non-dietary oral medicament for celiac patients12. The gluten peptides resistant to hydrolysis by proteolysis are found in the most of immunotoxic reactions in T cells of patients with CD13. Furthermore, the mammalian digestive enzymes are difficult to digest the resistant domain in gluten because of the cleavage site septicity of these enzymes. The proteolytic enzymes might be produced from a wide range of organisms such as; bacteria, insects, yeasts, molds, animals and plants14-16. The insect prolyl endopeptidases (PEPs) may be the alternative therapeutic keys for treatment of CD. These enzymes are effective in the hydrolysis of specific peptide bonds on the carboxyl group side of internal proline residues of the gluten oligopeptides17. The PEPs is one member of a serine protease family group, which are hydrolyze the peptides smaller than 30 residues18. The Red Palm Weevil (RPW), Rhynchophorus ferrugineus is a noxious internal feeder attacking many palm species including coconut, date palm and Canary Islands palm19,20. The RPW possess serine-type proteinases in their digestive system for protein digestion21,22. Therefore, this study was planned to find out natural, economical and alternative source of proteolytic enzymes that has capability to hydrolyze gluten and gliadin and to find out alternative medicine for celiac patients.
MATERIALS AND METHODS
The present investigation was carried out at the College of Pharmacy, Prince Sattam bin Abdulaziz University, Al-Kharj, Saudi Arabia and the Food Technology Research Institute, Agricultural Research Center, Giza, Egypt between February, 2018 and December, 2019.
Materials: Larvae of red palm weevil (Rhynchophorus ferrugineus) were collected from infected palm trees at the Agriculture Research Center, Cairo, Egypt. All chemicals and reagents used throughout this study were of analytical grade. The synthetic fluorogenic peptides Suc-Ala-Pro-pNA was obtained from Bachem, Torrance, CA.
Extraction of protease from larvae of red palm weevil: The larvae of R. ferrugineus were anesthetized by exposure to ether for seconds, the midguts were separated in washing buffer containing 20 mM Tris-HCl, pH 7.4, then washed by the same buffer several times. The fatty tissues were removed and a total of 30 cleaned midguts were homogenized in 20 mL lysis buffer containing 20 mM Tris-HCl pH 7.4, 2 mM β-mercaptoethanol, 5 mM EDTA. The homogenized solution was filtrated through double layer of cheese cloth and centrifuged at 6,000 rpm for 20 min23. The clear supernatant containing enzymes were lyophilized and stored in a refrigerator for further purification.
Extraction of gliadin: Gliadin was extracted by treatment of gluten with 2.5 mL of 70% ethanol (v/v) and stirred for 45 min and then centrifuged at 6000 rpm for 10 min24.
Endopeptidase purification: All purification steps were performed at 4°C, except when mentioned. After each step, fractions were assayed for protease activity and the active fractions were pooled. Three purification methods were used in this study, the first was the partial purification with ammonium sulfate. The protein extracts were subjected to 40, 60 and 80% of ammonium sulfate. The precipitate was dissolved in small amount of cold deionized water after centrifugation at 15,000 rpm for 20 min25. The second method was the using of acetone as organic solvent for precipitation of protein. The alkaline protease was collected from the crude extract by precipitation with 40-80% of acetone concentrations. The different amounts of each concentration of acetone were added slowly to the protein extract with stirring for 30 min and then allowed to settle down for 12 h at 4°C. The precipitate was dissolved in 0.1 M phosphate buffer, pH 7 after centrifugation at 15,000 rpm for 20 min26. While, the third method was precipitation by trichloroacetic acid (TCA). The protein extract was dissolved with 0.2 M TCA, centrifuged at 15,000 rpm for 20 min. The precipitate was dissolved in chilled acetone and again centrifuged at 14000 rpm for 5 min, this procedure was repeated twice to remove the remaining of TCA. The obtained pellet was dissolved in 0.1M of phosphate buffer, pH 7 and it used for protein and enzyme assay27.
Assay of protease activity: The activity of protease enzyme was determined by casein digestion assay as described by El-Beltagy et al.28. One unit of the enzyme activity is defined as the amount of enzyme required to liberates sufficient amounts of trichloroacetic acid, i.e., the absorbance at 280 nm increased by 1.00 in 1 min. Tyrosine was used as a standard.
Hydrolysis of peptidyl-pNA: The rate of enzymatic hydrolysis of the peptidyl-pNA substrate was measured in buffer containing 0.1 M Tris-HCl, pH 7.5 at 25°C by spectrophotometer at 410 nm29. The unit of the enzyme activity was defined as the enzyme quantity required to release 1 μmoL of p-nitroaniline per minute under the specific reaction conditions.
Hydrolysis of gluten and gliadin: Two different concentrations including 1 and 2 mg of the enzyme purified with 80% acetone was added to 100 mL of 2% gluten or gliadin in phosphate-citrate buffer pH 6.9, the reaction mixture incubated at 30°C for 2 h with shaking. The enzyme was subjected to heating at 100°C for 30 sec to inactivate its activity. The obtained hydrolyzed solution was centrifuged at 10000 rpm for 15 min and the percentage of hydrolysis was determined24.
Biochemical characterization of enzyme
Effect of pH and temperature on PEP activity and stability: In order to find the optimum pH, the enzyme was incubated with Suc-Ala-Pro-pNA substrate in buffers spanning the pH range of 2-12 and the activity was determined at the standard assay condition. Also, the optimal temperature of enzyme activity was measured by incubating the reaction mixture including enzyme, substrate and buffer at different temperature14 ranged from 20-60°C.
Effects of activators and inhibitors on PEP activity: Numbers of protease activators and inhibitors including MgCl2, CaCl2 and EDTA, DL-Dithiothreitol (DTT), phenylmethylsulfonyl fluoride (PMSF) and Soybean Trypsin Inhibitor (STI) were used for determination the enzyme activity. The total enzyme activity was determined in 0.1 M Tris-HCl buffer, pH 8.030. Each of the protease activators or inhibitors was used at three different concentration including 0, 5 and 10 mM, the Suc-Ala-Pro-pNA substrate was added. The reaction mixture was incubated at 30°C for l5 min, the protease activity was determined, this reaction was triplicates repeated for each treatment.
Effect of substrate concentration on PEP activity and kinetic constants: Enzyme kinetics and Michaelis constants (Km and Vmax) were determined to estimate the substrate concentration effect on the purified enzyme activity. This was carried out with using of Bovine Serum Albumin (BSA) as enzyme substrate. The Km and Vmax of the purified enzyme were determined from the Lineweaver-Burk plot27.
Free amino acids determination: The free amino acids were determined calorimetrically by using Ninhydrin reagent according to Morales et al.31 as follows, 3 mL of 5% trichloroacetic acid and 1 mL of phosphate buffer, pH 8 were added to 1 mL of the hydrolysate mixture incubated at room temperature for 15 min, the precipitate was removed by centrifugation at 6000 rpm for 15 min, then the supernatant was used for measuring of the amount of free amino acids by using of ninhydrin reagent, glycine was used as a standard.
Statistical analysis: In the current study, the obtained data were expressed as Means±SD from three replicates. The data were subjected to one-way ANOVA. The means values of the different treatments were compared using Duncan’s multiple range test at p<0.05. The statistical analyses were carried out by using SPSS statistical software (IBM SPSS Statistics, version 20)32.
RESULTS AND DISCUSSION
Fractional precipitation of protease enzyme extracts: The precipitation of the enzyme extract with different concentrations of ammonium sulphate was described in Fig. 1. The crude enzyme extract from RPW of 6 week old was contained 0.34 mg protein. The 80% of ammonium sulphate precipitated fraction showed high specific activity accounted by 5333 U mg–1 protein and purification fold 1.31 followed by 4917 U mg–1 protein at 60% precipitation. Moreover, the high specific activity of protease was obtained in the 80% fraction 7000 U mg–1 protein which represents 1.76 of purification fold followed by precipitation with 60% which exhibits 5667 U mg–1 protein for protease extracted from RPW of 12 weeks old.
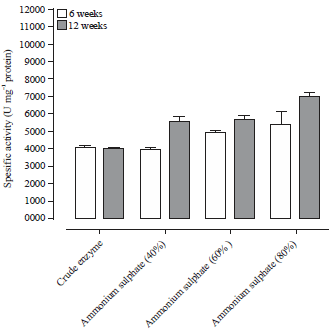 | |
| Fig. 1: | Purification of PEP extracted from RPW with ammonium sulfate |
PEP: Prolyl endopeptidase, RPW: Red palm weevil, data represent the Means±SE calculated from three replicates | |
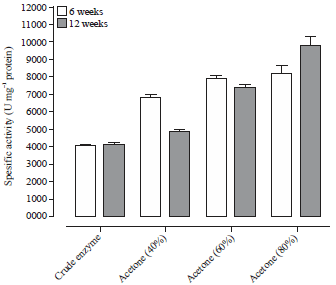 | |
| Fig. 2: | Purification of PEP extracted from RPW with acetone |
PEP: Prolyl endopeptidase, RPW: Red palm weevil, Data represent the Means±SE calculated from three replicates | |
The precipitation of protease extracted from RPW aged 6 and 12 weeks with acetone was showed in Fig. 2. The specific activity of the enzyme extracted from larvae of red palm weevil aged 6 weeks was 4059 U mg–1 protein. By partial fractionation with acetone at 40, 60 and 80%, the enzyme specific activity was increased by 6800, 7875 and 8200 U mg–1 protein, respectively.
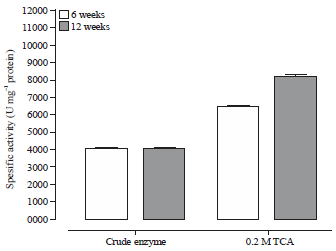 | |
| Fig. 3: | Purification of PEP extracted from RPW with 0.2 M TCA |
| PEP: Prolyl endopeptidase, RPW: Red palm weevil, data represent the Means±SE calculated from three replicates | |
On the other hand, the highest specific activity of protease enzymes at aged 12 weeks was recorded as 9800 U mg–1 protein which represents 2.40 purification fold at 80% acetone fractionation. Whereas, the enzyme specific activity was high in fraction of 80% acetone for both age of RPW and amounted as 9800 and 8200 U mg–1, respectively.
The purification of PEP extracted from RPW aged 6 and 12 weeks by 0.2 M TCA illustrated in Fig. 3. The specific activity of the purified protease with TCA at age 12 weeks was recorded as 8200 U mg–1 protein and it is higher than that of 6 week age which amounted as 6500 U mg–1 protein. Previous study reported the protease enzymes extracted with 0.1 M phosphate buffer at pH 7 and precipitated with 0.2 M TCA has high enzyme recovery due to the negative effect of TCA on the enzyme denaturation17.
Results concluded that, the best agents to purify PEP extracted from RPW at age 6 and 12 weeks were in the following order 80% acetone >80% ammonium sulfate>0.2 M TCA. Also, the purification of PEP extracted from RPW aged 12 weeks by 80% acetone give higher specific activity (9800 U mg–1 protein) which represents 2.40 of purification fold compared to all other treatments. This may be attributed to increasing of hydrolytic enzyme activity with increasing larvae growth before entering into pupa phase as reported by Mahmud et al.33. On other hands, multiple studies reported that, 80% acetone precipitation is the best agent to purify serine proteases extracted from RPW34,35.
Biochemical characterization of enzyme
Effect of pH and temperature on PEP activity and stability: The optimum activity of the purified enzyme with 80% acetone was determined at different range of pH 2.0, 4.0, 6.0, 8.0 and 10.0. As shown in Fig. 4, the optimum pH was observed at pH 5.0.
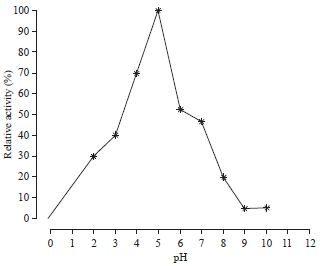 | |
| Fig. 4: | Effect of different pH on PEP activity |
| PEP: Prolyl endopeptidase | |
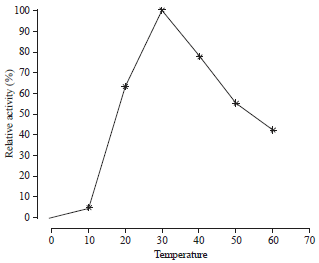 | |
| Fig. 5: | Effect of different temperature on PEP activity |
| PEP: Prolyl endopeptidase | |
This pH value was in agreement with that obtained by Peterson et al.36 who found the optimum pH was 10.0 and 10.5 for the trypsin activity extracted from midgut of S. littoralis and Manduca sexta L., respectively. The results also indicated that the PEP activity was maximized at 30°C, then the activity was gradually decreased by increasing the incubation temperature (Fig. 5).
Effects of activators and inhibitors on PEP: Different compounds (MgCl2, CaCl2, DTT, PMSF and STI) were used to estimate the protease activity at three concentrations (0, 5 and 10 mM) (Fig. 6). Under optimal pH, the effects of protease activators and inhibitors were determined on the midgut protease activity of Rhynchophorus ferrugineus aged 12 weeks and purified with 80% acetone. Different compounds (MgCl2, CaCl2, DTT, PMSF and STI) were used to estimate the protease activity at 3 concentrations 0, 5 and 10 M (Fig. 6).
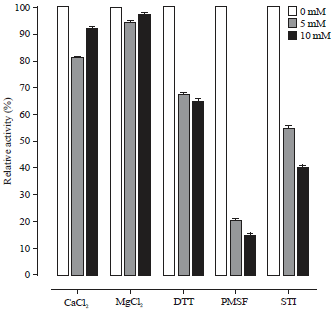 | |
| Fig. 6: | Effect of different activators and inhibitors on PEP activity |
| PEP: Prolyl endopeptidase | |
The relative activity of PEP was decreased by 20 and 8% at incubation with 5 and 10 mM of CaCl2, while, incubation with MgCl2 was almost keep the relative activity as same as control. On the other hand, DTT was inhibited the relative activity to 67 and 65% at the two concentrations. Although, PMSF was completely deactivated the enzyme activity, while, 5 and 10 mM of STI were reduced the activity to 55 and 40%, respectively.
Effect of substrate concentration on protease activity and kinetic constants: Enzyme kinetics and Michaelis constants (Km and Vmax) determined the effect of substrate concentration on the enzyme activity. Low Km means only small amount of substrate is needed to saturate the enzyme and high affinity between enzyme and substrate37. The effect of substrate (bovine serum albumin, BSA) concentration on the activity of partial purified PEP extracted from RPW was described in Fig. 7. The extracted PEP at age 12 weeks and purified by 80% acetone exhibited high value of Vmax (161.29 mg min–1), low value of Km (0.129 mg mL–1) and high specific activity.
Gluten and gliadin hydrolysis by PEP: The current study aimed to reach the maximum percentage of gluten and gliadin hydrolysis. Data presented in Table 1 illustrated the extracted PEP from RPW aged 12 weeks and purified by 80% acetone. It yielded the same percentage of gluten hydrolysis, 32% at 1 and 2 mg mL–1 of enzyme concentration and almost closed to the percentage of hydrolysis at 1 mg mL–1 of Digestin.
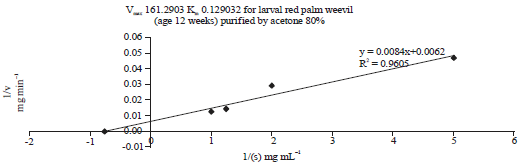 | |
| Fig. 7: | Lineweaver-burk plot show kinetic constants (Km and Vmax) of PEP extracted from larval red palm weevil (age 12 weeks) and purified by 80% acetone |
| PEP: Prolyl endopeptidase | |
| Table 1: | Gluten hydrolysis with different protease concentration |
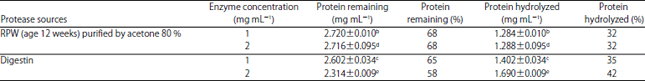 | |
*Data represent the Means±SE calculated from three replicates, Different letters refer to significant differences at p<0.05, Gluten concentration before hydrolysis was 4.004 mg mL–1, Digestin drug is composed of Papain and Sanzyme | |
| Table 2: | Gliadin hydrolysis with different protease concentration |
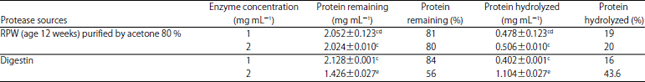 | |
| *Data represent the Means±SE calculated from three replicates, different letters refer to significant differences at p<0.05, Gliadin concentration before hydrolysis was 2.53 mg mL–1 | |
While Digestin give higher percentage of gluten hydrolysis by amount 35 and 42% at 2 concentration compared to the purified PEP, because of Digestin drug is composed of Papain and Sanzyme, while, the PEP is composed of only single enzyme.
Gliadin hydrolysis with PEP: The extracted PEP was also examined to hydrolyze gliadin which extracted from gluten. Gliadin considered the primary reason for celiac disease. Gliadin is a polypeptide consists of 33 amino acids with known sequence rich with proline and glutamine. Proline gives the peptide increased resistance to gastrointestinal proteolysis causing risk for CD38. In healthy peoples this polypeptide does not permeate into their intact intestine and excrete in an undigested form. Data presented in Table 2 demonstrated that, the purified protease give higher percentage of gliadin hydrolysis 19% at 1 mg mL–1 of enzyme concentration than the gliadin hydrolysis by Digestin which give only 16%. While, the percentages of hydrolyzed gliadin by Digestin at 2 mg mL–1 was equal to 43.6% and higher than the purified PEP that give 20% of hydrolysis.
Free amino acids concentration after gluten and gliadin hydrolysis: The free amino acids produced from hydrolysis of gluten were 0.52 and 0.54% at enzyme concentrations of 1 and 2 mg mL–1 while 0.56 and 0.65% with using Digestin at 1 and 2 mg mL–1, respectively (Table 3).
The results showed that the free amino acids liberated from gliadin hydrolysis by Digestin at enzyme concentrations 1 and 2 mg mL–1 were recorded as 1.01 and 1.25%, respectively and these were higher than that of percentage of hydrolysis by the purified PEP at the optimum incubation reaction (Table 4).
The using of gluten free diet is the only treatment of celiac disease. The treatment diet is expensive and difficult to getting it, as well as, it is not completely free gluten and this will directly effect on celiac patients. The recent alternative strategies are focus on the using of new therapeutic treatment for decreasing the celiac effects of wheat gliadins35, such as orally supplementation with proline digesting enzymes that could be appreciate method of elimination of the proline–rich T cell stimulatory epitopes from gluten.
| Table 3: | Free amino acids concentration from hydrolyzed gluten |
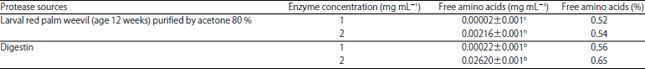 | |
| *Data represent the Means±SE calculated from three replicates, Different letters refer to significant differences at p<0.05, Gluten concentration before hydrolysis was 4.004 mg mL–1 | |
| Table 4: | Free amino acid concentration in hydrolyzed gliadin |
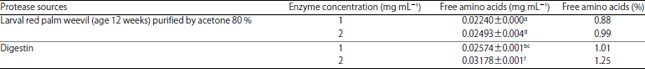 | |
| *Data represent the Means±SE calculated from three replicates, Different letters refer to significant differences at p<0.05, Gliadin concentration before hydrolysis was 2.53 mg mL–1 | |
These will be performed by transglutaminase inhibition37 and HLA-DQ2 blocking to prevent of immunogenic peptides binding39 or by using more recent promising alternative therapeutic approach as the enzymatic cleavage of immunogenic gliadins, the purified prolyl-endopeptidase obtained from the present study might be used for this purpose.
Earlier studies reported that, the prolyl oligopeptidases are not stable at the low pH of gastric juice39, possibility to digestion with pepsin40 and not enough active at high amount of gluten41. As well as, the prolyl oligopeptidases protection from gastric juice by encapsulation is not effective because of the gluten degradation will not carried out before it reaches to the proximal part of the duodenum, whereas the gluten induces inflammatory T cell responses42. Furthermore, AN-PEP enzyme was extracted from Aspergillus niger was active at pH ranged from 2-8, with an optimum activity at pH 4-5, therefore, it is effective under stomach conditions and remains fully functional43. Another advantage of this enzyme is its capability of whole gluten and gluten peptides into non-immunogenic residues degredation44. The prolyl endopeptidases have pharmaceutical and medicinal applications, as well as, play an important role in the various foodstuffs production due to its need for an efficient of gluten degradation in raw material or during food processing. The prolyl endopeptidases belong to serine proteases that can be peptide bonds cleavage at the carboxy-terminal end of proline residues. The PEP under investigation was extracted from Rhynchophorus ferrugineus aged 12 weeks and purified by 80% acetone. According to the pH profile, the enzyme showed the highest activity at pH 5.0 and between 40-70% of the activity was retained at pH 3.0 and 5.0. The PEP was stable at up to 50°C, with a relative activity of approximately 80% incubating at pH 5.0 for 1 h. The optimum temperature for the PEP was established at 30°C, whereas, a decrease in activity was observed when the enzyme was heated at temperature above 50°C. The relative protease activity remained similar when incubated with MgCl2, CaCl2 and DTT. While, PMSF and STI are inhibited the activity to 20 and 15% at 5 and 10 mM of PMSF and 60; 40% for STI, when incubated with the peptide Suc-Ala-Pro-pNA.
It is demonstrated that the prolyl endopeptidase from Rhynchophorus ferrugineus can act under conditions similar to those found in the gastrointestinal tract and is capable of degrading intact gluten molecules and T cell stimulatory epitopes from gluten into harmless fragments, therefore, the purified enzyme can be used as alternative treatment of celiac disease.
CONCLUSION
The prolyl oligopeptidases extracted from larvae of palm weevil at age 12 weeks and purified with 80% acetone is the potential alternative recent drug for CD treatment due to its capability to resist the low gastric pH and efficiency to digest gluten and gliadin at the same condition of stomach digestion.
SIGNIFICANCE STATEMENT
This study extracted and purified of a new specific endopeptidases of insect origin for serving as a potential therapeutic tool for degradation of wheat gluten and gliadins, the main causes of celiac disease.
ACKNOWLEDGMENT
The authors would like to thank the College of Pharmacy at Prince Sattam Bin Abdulaziz University, Al-Kharj, Kingdom of Saudi Arabia for providing the necessary facilities to carry out this research work.
REFERENCES
- Caio, G., U. Volta, A. Sapone, D.A. Leffler, R. De Giorgio, C. Catassi and A. Fasano, 2019. Celiac disease: A comprehensive current review. BMC Med., Vol. 17, No. 1.
CrossRefDirect Link - Moreno, M.L., A. Cebolla, A. Muñoz-Suano, C. Carrillo-Carrion and I. Comino et al., 2017. Detection of gluten immunogenic peptides in the urine of patients with coeliac disease reveals transgressions in the gluten-free diet and incomplete mucosal healing. Gut, 66: 250-257.
CrossRefDirect Link - Ortiz, C., R. Valenzuela and A.Y. Lucero, 2017. Celiac disease, non celiac gluten sensitivity and wheat allergy: Comparison of 3 different diseases triggered by the same food. Rev. Chil. Pediatr., 88: 417-423.
CrossRefPubMedDirect Link - Bittker, S.S. and K.R. Bell, 2019. Bittker, S.S. and K.R. Bell, 2019. Potential risk factors for celiac disease in childhood: A case-control epidemiological survey. Clin. Exp. Gastroenterol., 12: 303-319.
CrossRefPubMedDirect Link - Rosell, C., F. Barro, C. Sousa and M.C. Mena, 2014. Cereals for developing gluten-free products and analytical tools for gluten detection. J. Cereal Sci., 59: 354-364.
CrossRefDirect Link - Calasso, M., R. Francavilla, F. Cristofori, M. de Angelis and M. Gobbetti, 2018. New protocol for production of reduced-gluten wheat bread and pasta and clinical effect in patients with irritable bowel syndrome: A randomised, double-blind, cross-over study. Nutrients, Vol. 10, No. 12.
CrossRefDirect Link - Deora, N.S., A. Deswal and H.N. Mishra, 2015. Functionality of alternative protein in gluten-free product development. Food Sci. Technol. Int., 21: 364-379.
CrossRefDirect Link - Moreno, M.L., A. Rodríguez-Herrera, C. Sousa and I. Comino, 2017. Biomarkers to monitor gluten-free diet compliance in celiac patients. Nutrients, Vol. 9, No. 1.
CrossRefDirect Link - Dessì, M., A. Noce, S. Vergovich, G. Noce and N. Di Daniele, 2013. Safety food in celiac disease patients: A systematic review. Food Nutr. Sci., 4: 55-74.
CrossRefDirect Link - Comino, I., A. Real, S. Vivas, M.Á. Síglez and A. Caminero et al., 2012. Monitoring of gluten-free diet compliance in celiac patients by assessment of gliadin 33-mer equivalent epitopes in feces. Am. J. Clin. Nutr., 95: 670-677.
CrossRefDirect Link - Ciacci, C., P. Ciclitira, M. Hadjivassiliou, K. Kaukinen and J.F. Ludvigsson et al., 2015. The gluten-free diet and its current application in coeliac disease and dermatitis herpetiformis. United Eur. Gastroenterol. J., 3: 121-135.
CrossRefDirect Link - Helmerhorst, E.J., M. Zamakhchari, D. Schuppan and F.G. Oppenheim, 2010. Discovery of a novel and rich source of gluten-degrading microbial enzymes in the oral cavity. PLoS One, Vol. 5, No. 10.
CrossRefDirect Link - Shan, L., T. Marti, L.M. Sollid, G.M. Gray and C. Khosla, 2004. Comparative biochemical analysis of three bacterial prolyl endopeptidases: Implications for coeliac sprue. Biochem. J., 383: 311-318.
CrossRefDirect Link - Iqbalsyah, T.M., Malahayati, Atikah and Febriani, 2019. Purification and partial characterization of a thermo-halostable protease produced by Geobacillus sp. strain PLS A isolated from undersea fumaroles. J. Taibah Univ. Sci., 13: 850-857.
CrossRefDirect Link - Mohammad, V., H. Vahid, G. Mohammad and M.S. Seyede, 2014. Plant cell wall degrading enzymes, pectinase and cellulase, in the digestive system of the red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae). Plant Protect. Sci., 50: 190-198.
CrossRefDirect Link - Hasnain, S.Z., M.A. McGuckin, R.K. Grencis and D.J. Thornton, 2012. Serine protease (s) secreted by the nematode Trichuris muris degrade the mucus barrier. PLoS Negl. Trop. Dis., Vol. 6, No. 10.
CrossRefDirect Link - Rea, D. and V. Fülöp, 2006. Structure-function properties of prolyl oligopeptidase family enzymes. Cell Biochem Biophys., 44: 349-365.
CrossRefDirect Link - Ehren, J., B. Morón, E. Martin, M.T. Bethune, G.M. Gray and C. Khosla, 2009. A food-grade enzyme preparation with modest gluten detoxification properties. PloS One, Vol. 4, No. 7.
CrossRefDirect Link - Dembilio, Ó., G.V. Tapia, M.M. Téllez and J.A. Jacas, 2012. Lower temperature thresholds for oviposition and egg hatching of the red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae), in a Mediterranean climate. Bull. Entomol. Res., 102: 97-102.
CrossRefDirect Link - Hoddle, M.S., A.H. Al-Abbad, H.A.F. El-Shafie, J.R. Faleiro, A.A. Sallam and C.D. Hoddle, 2013. Assessing the impact of areawide pheromone trapping, pesticide applications and eradication of infested date palms for Rhynchophorus ferrugineus (Coleoptera: Curculionidae) management in Al Ghowaybah, Saudi Arabia. Crop Protect., 53: 152-160.
CrossRefDirect Link - Alarcon, F.J., T.F. Martınez, P. Barranco, T. Cabello, M. Diaz and F.J. Moyano, 2002. Digestive proteases during development of larvae of red palm weevil, Rhynchophorus ferrugineus (Olivier, 1790) (Coleoptera: Curculionidae). Insect Biochem. Mol. Biol., 32: 265-274.
CrossRefDirect Link - Capocchi, A., M. Cinollo, L. Galleschi, F. Saviozzi, L. Calucci, C. Pinzino and M. Zandomeneghi, 2000. Degradation of gluten by proteases from dry and germinating wheat (Triticum durum) seeds: An in vitro approach to storage protein mobilization. J. Agric. Food Chem., 48: 6271-6279.
CrossRefPubMedDirect Link - Contesini, F.J., R.R.D. Melo and H.H. Sato, 2018. An overview of Bacillus proteases: From production to application. Crit. Rev. Biotechnol., 38: 321-334.
CrossRefDirect Link - Geethanjali, S. and A. Subash, 2013. Comparative study on precipitation techniques for protease isolation and purification from Labeo rohita Viscera. J. Aquat. Food Prod. Technol., 22: 121-128.
CrossRefDirect Link - Upadhyay, S.K., R.T. Magar and C.J. Thapa, 2013. Biochemical characterization of protease isolated from different parts of Choerospondias axillaris (Lapsi). Biochem. Anal. Biochem., Vol. 2, No. 3.
CrossRef - El-Beltagy, A.E., T.A. El-Adawy, E.H. Rahma and A.A. El-Bedawey, 2005. Purification and characterization of an alkaline protease from the viscera of bolti fish (Tilapia nilotica). J. Food Biochem., 29: 445-458.
CrossRefDirect Link - Erlanger, B.F., N. Kokowsky and W. Cohen, 1961. The preparation and properties of two new chromogenic substrates of trypsin. Arch. Biochem. Biophys., 95: 271-278.
CrossRefPubMedDirect Link - Zhao, A., X. Yuan, D. Hu, C. Leng, Y. Li, P. Wang and Y. Li, 2019. The effect of host plant on the development and larval midgut protease activity of Plutella xylostella (Lepidoptera: Plutellidae). Phytoparasitica, 47: 475-483.
CrossRefDirect Link - Morales, A., L. Buenabad, G. Castillo, L. Vázquez, S. Espinoza, J.K. Htoo and M. Cervantes, 2017. Dietary levels of protein and free amino acids affect pancreatic proteases activities, amino acids transporters expression and serum amino acid concentrations in starter pigs. J. Anim. Physiol. Anim. Nutr., 101: 723-732.
CrossRefDirect Link - Snedecor, G.W. and W.G. Cochran, 1980. Statistical Methods. 7th Edn., Iowa State University Press, Iowa, USA., ISBN-10: 0813815606, Pages: 507.
Direct Link - Mahmud, A.I., J. Farminhao and E.R. Viez, 2015. Red palm weevil (Rhynchophorus ferrugineus Olivier, 1790): Threat of palms. J. Biol. Sci., 15: 56-67.
CrossRefDirect Link - Al-Jab, A. and Abo-El-Saad, 2008. A putative serine protease from larval midgut of red palm weevil Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae): Partial purification and biochemical characterization. Am. J. Environ. Sci., 4: 595-601.
CrossRefDirect Link - Al-Khaldi, E.M. and A.H. Alabdalall, 2016. Effect of activators and inhibitors on extracellular thermostable alkaline protease isolated from Bacillus subtilis obtained from eastern province of Saudi Arabia. Afr. J. Biotechnol., 16: 232-237.
CrossRefDirect Link - Peterson, A.M., C.V. Barillas-Mury and M.A. Wells, 1994. Sequence of three cDNAs encoding an alkaline midgut trypsin from Manduca sexta. Insect Biochem. Mol. Biol., 24: 463-471.
CrossRefDirect Link - Siegel, M., M.T. Bethune, J. Gass, J. Ehren and J. Xia et al., 2006. Rational design of combination enzyme therapy for celiac sprue. Chem. Biol., 13: 649-658.
CrossRefDirect Link - Van den Broeck, H.C., H.C. de Jong, E.M. Salentijn, L. Dekking and D. Bosch et al., 2010. Presence of celiac disease epitopes in modern and old hexaploid wheat varieties: Wheat breeding may have contributed to increased prevalence of celiac disease. Theor. Applied Genet., 121: 1527-1539.
CrossRefDirect Link - Polgar, L., 1992. Prolyl endopeptidase catalysis. A physical rather than a chemical step is rate-limiting. Biochem. J., 283: 647-648.
CrossRefDirect Link - Matysiak-Budnik, T., C. Candalh, C. Cellier, C. Dugave and A. Namane et al., 2005. Limited efficiency of prolyl-endopeptidase in the detoxification of gliadin peptides in celiac disease. Gastroenterology, 129: 786-796.
CrossRefDirect Link - Gass, J., J. Ehren, G. Strohmeier, I. Isaacs and C. Khosla, 2005. Fermentation, purification, formulation, and pharmacological evaluation of a prolyl endopeptidase from Myxococcus Xanthus: Implications for celiac sprue therapy. Biotechnol. Bioeng., 92: 674-684.
CrossRefDirect Link - Stepniak, D., L. Spanij-Dekking, C. Mitea, M. Moester and A. de Ru et al., 2006. Highly efficient gluten degradation with a newly identified prolyl endoprotease: Implications for celiac disease. Am. J. Physiol. Gastrointest. Liver Physiol., 291: G621-G629.
CrossRefPubMedDirect Link - Mitea, C., R. Havenaar, J.W. Drijfhout, L. Edens, L. Dekking and F. Koning, 2008. Efficient degradation of gluten by a prolyl endoprotease in a gastrointestinal model: implications for coeliac disease. Gut, 57: 25-32.
CrossRefPubMedDirect Link - Edens, L., P. Dekker, R. Van Der Hoeven, F. Deen, A. de Roos and R. Floris, 2005. Extracellular prolyl endoprotease from Aspergillus niger and its use in the debittering of protein hydrolysates. J. Agric. Food Chem., 53: 7950-7957.
CrossRefDirect Link