
Hui-Fang Chiu
Department of Chinese Medicine, Taichung Hospital Ministry of Health and Well-being, Taichung, Taiwan, Republic of China
Shine-Ling Huang
School of Nutrition, Chung Shan Medical University, 110, Sec. 1, Jianguo North Road, Taichung City, Taiwan, Republic of China
You-Cheng Shen
School of Health Diet and Industry Management, Chung Shan Medical University, 110, Sec. 1, Jianguo North Road, Taichung City, Taiwan, Republic of China
Yi-Chun Han
School of Nutrition, Chung Shan Medical University, 110, Sec. 1, Jianguo North Road, Taichung City, Taiwan, Republic of China
Kamesh Venkatakrishnan
School of Nutrition, Chung Shan Medical University, 110, Sec. 1, Jianguo North Road, Taichung City, Taiwan, Republic of China
LiveDNA: 91.20696
Chin-Kun Wang
School of Nutrition, Chung Shan Medical University, 110, Sec. 1, Jianguo North Road, Taichung City, Taiwan, Republic of China
LiveDNA: 886.26538
International Journal of Pharmacology
Year: 2019 | Volume: 15 | Issue: 5 | Page No.: 533-541







ABSTRACT
Background and Objective: The grape seed polyphenol (GSP) is rich in flavonoids and tannins which contributes to diverse pharmacological activities. The current study was undertaken to assess the anti-melanogenesis effect of GSPE on UVA-exposed B16F1 melanoma cells by checking morphological and biochemical changes. Materials and Methods: The present study was sub-divided into three phases. In phase I, the anti-melanogenesis effect of crude grape seed polyphenol extract (GSPE) was checked by comparing with three commercial grape products (A, B and C) in non-UVA exposed B16F1 cells. During phase II, the anti-melanogenesis effect of GSPE was determined on UV-exposed B16F1 cells to reveal the effective dosage. Finally, in phase III the anti-melanogenesis effect of different experimental samples (GSPE; 50 μg, Vit C; 1 μg and combo of GSPE- Vit C; 50:1μg) were examined on UV-exposed B16F1 cells. Results: The crude GSPE at the dose of 50 μg exhibited better (p<0.05) melanin inhibitory activity (suppressed melanin synthesis) than commercial grape products (phase I) as well as successfully attenuated (p<0.05) UV-induced photo-oxidative stress by its free radical scavenging activity in phase II. All the experimental samples in phase III presented anti-melanogenesis activity, but crude GSPE (50 μg) displayed the greatest (p<0.05) melanin inhibitory activity by down regulating tyrosinase activity, thus proving its anti-melanogenesis activity. Conclusion: In this study, crude GSPE (50 μg) showed the maximum melanin inhibitory activity as it successfully down regulated the tyrosinase activity.
PDF Abstract XML References Citation
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Hui-Fang Chiu, Shine-Ling Huang, You-Cheng Shen, Yi-Chun Han, Kamesh Venkatakrishnan and Chin-Kun Wang, 2019. Inhibitory Effect of Grape Seed Polyphenol Extract and Vitamin C on Melanogenesis in Cultured B16-F1 Melanoma Cells. International Journal of Pharmacology, 15: 533-541.
DOI: 10.3923/ijp.2019.533.541
URL: https://scialert.net/abstract/?doi=ijp.2019.533.541
DOI: 10.3923/ijp.2019.533.541
URL: https://scialert.net/abstract/?doi=ijp.2019.533.541
INTRODUCTION
Melanin is a dark biological pigment secreted by melanocyte in the basal layer of human skin during chronic sunlight exposure or in high oxidative stress condition or during other hyperpigmentation diseases such as melanoma, melasma and leukomelanosis1. During normal condition, melanin provides photoprotection to the skin against ultraviolet (UV) radiation including UVA and UVB, but excessive level trigger various deleterious events2. Tyrosinase (tyrosinase-related protein; TRP-1 and 2) is a copper-containing enzyme that catalyzes the conversion of L-tyrosine into 3, 4-dihydroxyphenylalanine (DOPA) and DOPA to DOP Aquinone. Tyrosinase is a rate-limiting enzyme for the melanogenesis as it catalyzes the first two steps of melanin synthesis3. Hence, it’s coherent to look up for natural tyrosinase inhibitors (the whitening agent) for treating dermal disorders related to hyperpigmentation as most of the synthetic whitening agents at present are linked up with several adverse effects and also expensive.
A number of studies have been reported that exposure to UVA may promote melanogenesis via excessive generation of free radicals, especially reactive oxygen species (ROS-H2O2), thus leads to oxidative stress4,5. Hence its logic to use polyphenol with potential antioxidant, anti-inflammatory and immunomodulatory properties that might influence an effective melanin inhibitory effect by lowering oxidative stress and its subsequent biochemical events for hyperpigmentation. Oligomeric proanthocyanidins (OPCs; condensed tannin) a sub-class of flavonoids have been reported to inhibit melanogenesis in melanoma cells6. The grape seed polyphenol (GSP) comprise of various flavonoids (gallic acid, flavan-3-ols catechin, epicatechin 3-O-gallate and procyanidin and more highly polymerized procyanidins) and tannins. However, among these flavonoids, oligomeric proanthocyanidins (OPCs) is the most abundant7. The OPCs are basically composed of flavan-3-ols units. The OPCs are potent antioxidants that also exhibit diverse pharmacological activities such as anti-inflammatory, anti-carcinogenic, anti-microbial, immunomodulatory hepatoprotective and hypotensive activities8,9.
Even though numerous reports showed the anti-melanogenesis effect of grape seed10,11 and Vit C5,12 but till now no combinational and comparative anti-melanogenesis studies were conducted using different experimental samples (GSPE, Vit C and GSPE-Vit C complex) on UV-induced melanoma cell line. Therefore, the present study was designed to examine the comparative anti-melanogenesis effect of GSPE, Vit C and GSPE-Vit C complex (combination) on melanogenesis of B16F1 melanoma cells. The anti-melanogenesis activity was assessed with respect to morphological and biochemical changes in terms of cell viability (cytotoxicity), protein contents, oxidative status, melanin contents and tyrosinase activity.
MATERIALS AND METHODS
Preparation of grape seed polyphenol extract (GSPE): The grape seeds (Vitis vinifera L.) were provided by Taiwan Tobacco and Liquor Corporation, Taiwan. The seeds were separated using vibrating sieves and further cleaned manually and dried at 60°C stored in dark under vacuum at room temperature. Dried seeds were milled into fine powder followed by extraction with 80% aqueous acetone (w:v = 1:5) and incubated at room temperature for 150 min and the resulting extract was filtered by using Whatman No. 4 filter paper. Repeat this filtering procedure for twice. The combined filtrates were freeze, dried by a vacuum pump and filled with nitrogen and then stored at -20°C until use. This in vitro study was carried out at Chung Shan Medical University, Taiwan, from 20th March-10th April, 2017.
Commercial grape seed extract: Three commercial grape products (A, B and C) were purchased from Ferco Development, France. Product A: Grape seed with, 95% OPCs (insoluble in water), product B: Grape seed with, 95% OPCs (soluble in water), product C: Grape skin 30% OPCs. L-ascorbic acid (Vit C) were purchased from Sigma, St. Louis, MO, USA. The crude GSPE, Vit C and commercial grape products (A, B and C) were dissolved in culture medium containing dimethyl sulfoxide (DMSO) and diluted to appropriate concentrations.
Cell culture studies: B16F1 murine melanoma cells were bought from the American Type Culture Collection Center (MD, USA). Cells were cultured in Dulbecco's Modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS), 1% penicillin and streptomycin (Gibco, NY, USA) in 5% CO2 at 37°C.
UVA irradiation: The method of UVA-promoted melanin formation was employed as described by Neitmann et al.13 with some modification. Briefly, melanoma cells B16F1 were irradiated by UVA filter (320-400 nm) at room temperature using a Vilber Lourmat UV lamp (with a light intensity of 0.72 mJ cm–2 at a distance of 5 cm, which was measured by Vilber Lourmat radiometer). Prior to UVA irradiation, the culture medium must be removed and B16F1 cells were treated with different experimental samples and washed with phosphate buffered saline (PBS). Cellular melanin contents and tyrosinase activities were measured immediately.
Morphological analysis: For morphological analysis, B16F1 melanoma cells with or without UVA exposure and GSPE treated cells were observed under a normal phase contrast microscope (Nikon Eclipse T 100, Tokyo, Japan) and photographed using a digital camera (Scopetek, Inc., Hangzhou, China).
Biochemical analysis
Cell viability assay: The viability of B16F1 melanoma cells was studied using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) method. Cells (200 μL; 1×105/mL) were seeded in each well of 96-well plates and allowed to adhere completely to the plate overnight. Next day media was discarded and substituted with new media and mixed with increasing concentrations of GSPE, Vit C at 37°C for 24 h. After samples treatment, MTT was added (100 μL) to each well. Cells were incubated at 37°C for 30 min and DMSO was added to dissolve the formazan crystals. The absorbance was measured at 560 nm with a spectrophotometer.
Determination of free radical generation (ROS): The intracellular ROS generation was assayed using 2',7'-dichlorofluorescein diacetate (DCFH-DA) by the method of Bai and Cederbaum14. Briefly, B16F1 cells were co-cultured with different concentrations of experimental samples for 1 h and followed by DCFH-DA (5 μM) in dark condition for 1 h (both with and with UVA-irradiated cells). Then the melanoma cells were mixed with PBS solution and the absorbance (OD) was read immediately using a fluorescence spectrophotometer (λex = 488 nm and λem = 525 nm). Results were expressed as arbitrary units of fluorescence per 106 cells.
Melanin contents: The melanin content of the cultured B16F1 melanoma cells was measured by the method of Rosenthal et al.15 with minor alterations. Melanoma cells were rinsed with PBS and treated with the mixture of ethyl alcohol and diethyl ether (3:1) with different concentration of samples (GSPE, Commercial product, Vit C and GSPE-Vit C complex) and centrifuged at 10,000 g. The resulting cell pellet was resuspended in 1 mL of 1 N NaOH or KOH. The optical density of each cultured wells was measured using an ELISA reader at 400 nm. The amount of melanin (cellular) was corrected according to the protein content of the samples. The protein content was determined by Bio-rad Protein Assay Kit (Bio-Rad, CA, USA) with BSA as the standard.
Assay of cell-free tyrosinase inhibition assay (cell-free): A cell-free tyrosinase activity was assayed as DOPA oxidase activity by the method of Yagi et al.16 with a slight alteration. The reaction mixture contained 70 μL of 0.1 M phosphate buffered saline (PBS, pH 6.8) with different concentrations of experimental samples and 20 μL of 10 μg mL–1 mushroom tyrosinase in 96-well plate. The 10 μL of 10 mM L-DOPA was also added to this mixture. The amount of dopachrome produced in the reaction mixture was determined against a blank and the absorbance was measured at 475 nm using a microplate reader. The inhibition of tyrosinase was calculated in percentage by using the equation:
![]()
| A | = | Absorbance at 475 nm without test sample |
| B | = | Absorbance at 475 nm with the test sample |
Assay of cellular tyrosinase activity: Tyrosinase activity was measured as described by Kojima et al.17. Briefly, B16F1 melanoma cells were cultured with or without samples in a Petri dish for 24 h followed by washing with potassium phosphate buffered saline (PBS, pH 7.4) and disrupted in PBS containing 1% Triton X-100. Then, cells were lysed by a sonicator and the lysates were centrifuged at 10,000×g for 10 min. The supernatant was used for assaying tyrosinase activity. Then 0.4-0.5 mL of supernatant was added in each well of a 96-well plate which contained approximately 40 μg protein, 2.5 mM L-DOPA and 0.1 M PBS. After incubation at 37°C for 1 h, the absorbance (as optical density (OD)) was measured at 475 nm in an ELISA reader for every 5 min. The specific activity of tyrosinase was expressed as units per liter (U L–1) from ΔE/min at 475 nm.
Statistical analysis: Results were explicated as the mean±standard deviation (SD). Each experiment was tested in triplicate. The difference in the results of this study was explored by using one-way ANOVA for comparison between each group and Student t-test for comparison within the group using SPSS software (21.0; SPSS Inc.). The p<0.05 was deemed as statistically significant.
RESULTS
Effect of GSPE on morphological changes and protein quantification: The phase contrast microscope revealed no notable changes in the morphology of melanoma cells upon treatment with different dosage and duration (24, 48 and 72 h) of GSPE in UVA unexposed (Fig. 1).
 | |
| Fig. 1(a-l): | Microscopic examination of normal B16F1 cells without exposure of UVA light for (a-d) 24 h, (e-h) 48 h and (i-l) 72 h portrait no visible change or pigmentation in melanoma cells (400x) after treatment with different doses of GSPE |
Whereas, B16F1 melanoma cells exposed to UVA light and treated with 50 μg of GSPE did not show any hyperpigmentation or any other morphological changes in melanoma cells (Fig. 2). Whereas the protein content of UVA unexposed (Fig. 3a) and exposed (Fig. 4a) B16F1 cells did not show any marked alteration on treatment with GSPE.
Effect of GSPE on cell viability: The MTT assays are indicating the cell viability or cytotoxicity on B16F1 cells treated with GSPE with or without UV exposure. There were no significant changes were noted in the case of viability in both the case, even at the higher concentration or UVA irradiation. Almost, 93% of melanoma cells were viable in the GSPE alone treatment group without UV exposure (Fig. 3b), whereas 91% of melanoma cells are surviving in the GSPE alone treated group with UV exposure (Fig. 4b) when compared with control cells.
Effect of GSPE on oxidative stress (ROS): The levels of intracellular ROS have concomitantly elevated in UVA-exposed cells as compared with the control group (Fig. 5). Treatment with increased concentration of GSPE exhibited a marked reduction in ROS level. The higher dose of GSPE (50 μg) showed maximum inhibitory levels of ROS on equivalence with other doses.
Effect of various experimental samples on melanin: A significant reduction in the levels of melanin content was observed in GSPE (50 μg) than commercial grape products (A, B and C) in UVA unexposed B16F1 cells (Fig. 6).
 | |
| Fig. 2(a-b): | Microscopic examination of normal B16F1 cells with exposure of UVA light for 72 h, (a) Control and (b) Treated with 50 μg of GSPE portrait no marked alteration or hyper-pigmentation in melanoma cells |
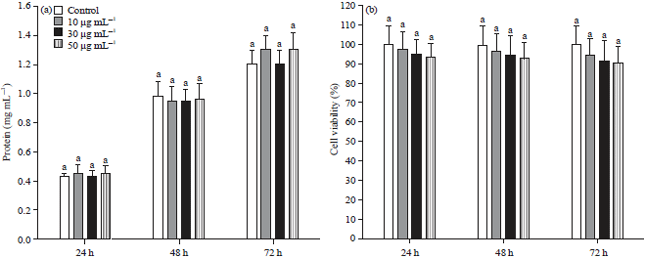 | |
| Fig. 3(a-b): | Effect of GSPE on the (a) Protein content and (b) Cell viability in B16F1 cells without UVA exposure |
Values were expressed as mean±SD. Data bearing different letters were significantly different (p<0.05) | |
Similarly, GSPE (50 μg) treatment concomitantly suppressed the melanin content in UV exposed B16F1 cells (Fig. 7). While, the melanin content was significantly suppressed on the addition of GSPE and combination of GSPE-Vit C in UV exposed B16F1 cells (Fig. 8), without any change in Vit-C alone group. Nevertheless, GSPE alone group showed lowered melanin content in UV exposed B16F1 cells than other experimental samples.
Effect of various experimental samples on tyrosine activity (cell-free and B16F1 cell model): All the experimental groups (GSPE, Vit C and GSPE-Vit C) showed greater inhibition on mushroom tyrosinase (cell-free model) as well as in B16F1 cellular model. However, the GSPE alone group showed the highest inhibition on mushroom tyrosinase than Vit C or GSPE-Vit C group (Fig. 9). The GSPE showed 4.68-fold of inhibition, but Vit C and GSPE-Vit C group also showed 2.14 and 3.70 fold of inhibition, respectively. Whereas, overall, GSPE alone group showed the highest inhibition on tyrosinase activity than other experimental groups (Fig. 10).
DISCUSSION
The present in vitro study was segregated into three phases (I, II and III). During phase I, no considerable changes in morphology or biochemical events, especially cell viability and protein content were noted in GSPE or commercial product of grapes indicated that natural materials from grape did not activate apoptosis or necrosis in a melanoma cell line and demonstrated its cytoprotective nature.
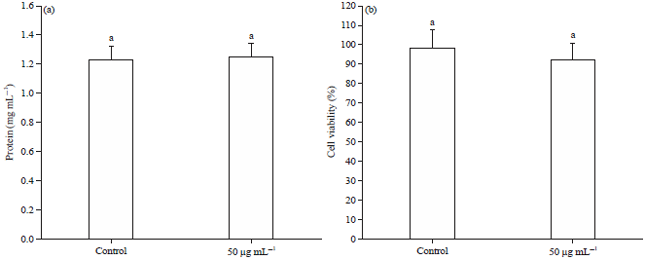 | |
| Fig. 4(a-b): | Effect of GSPE on the (a) Protein content and (b) Cell viability in UVA exposed B16F1 cells |
Values were expressed as mean±SD. Data bearing different letters were significantly different (p<0.05) | |
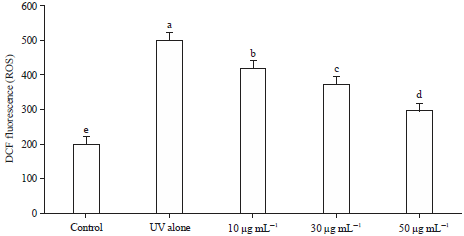 | |
| Fig. 5: | Effect of GSPE on oxidative stress (ROS) in UVA exposed B16F1 cells |
Values were expressed as mean±SD. Data bearing different letters were significantly different (p<0.05) | |
Hence, further studies are focused on the melanogenesis pathway instead of apoptosis or necrosis. GSPE (50 μg) showed a good melanin inhibitory effect on B16F1 cell as compared to commercial products. The outcome of phase I clearly indicated that melanogenesis was slightly triggered in B16F1 cells by secreting α-MSH owing to overproduction of free radical (oxidative stress)17.
In correlation with the results of phase I, GSPE showed no significant change in the levels of protein content, cell viability and morphological changes even after the exposure to UVA light (phase II), which indicated that GSPE was more cytoprotective by effectively scavenging the free radicals generated by UVA. The levels of ROS (DCFH) were substantially increased in UV-irradiated melanoma cells owing to increased ROS generation, but on treatment with an increased concentration of GSPE substantially lowered the levels of ROS in a dose-dependent fashion, due to free radical scavenging activity8,10. Total antioxidant status in UV exposed melanoma cell line also showed decreased antioxidant activity in DMSO (control) group, but when treated with different concentrations of GSPE, the total antioxidant capacity was enormously improved. Melanoma cell exposed to UV light showed increased oxidative stress via triggering α-MSH secretion, which in turn activates MITF (microphthalmia-associated transcription factor) and thereby positively modulates the transcription of tyrosinase and subsequent melanin synthesis2,18. Therefore, by lowering the oxidative stress, the activation of MITF can be downregulated and thus inhibit tyrosinase activity and melanogenesis19.
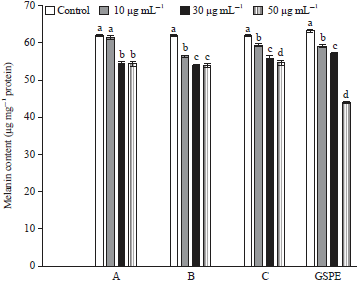 | |
| Fig. 6: | Effect of GSPE, commercial compounds (A, B and C) on Melanin content in B16F1 cells without UVA exposure |
Values were expressed as mean±SD. Data bearing different letters were significantly different (p<0.05) | |
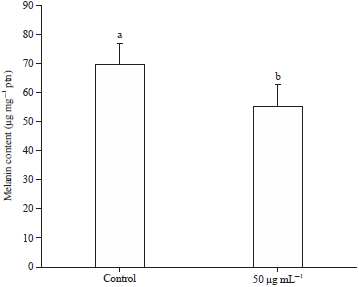 | |
| Fig. 7: | Effect of GSPE (50 μg) on Melanin content in UVA exposed B16F1 cells |
Values were expressed as mean±SD. Data bearing different letters were significantly different (p<0.05) | |
B16F1 cells treated with GSPE can effectively suppress free radical generation (ROS), which in turn inhibit the activation of α-MSH, MITF and TRP-1/2 and hence inhibited melanin synthesis. Previously, grape seed polyphenol is reported to enhance antioxidative status by positively regulating the Nrf2/HO-1 pathway20.
The inhibitory activity of mushroom tyrosinase by GSPE, were significantly higher in GSPE alone group than Vit C or GSPE-Vit C group indicating its direct action on TRP 1 and 2. However, the mechanism is yet to prove, but it has a positive regulation over melanin inhibition.
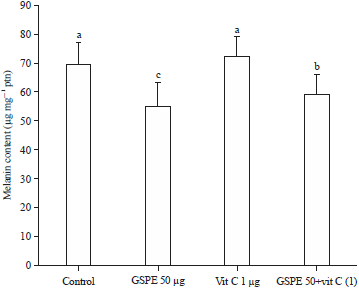 | |
| Fig. 8: | Effect of various experimental group (GSPE, Vit C and GSPE-Vit C) on melanin content in UVA exposed B16F1 cells |
Values were expressed as mean±SD. Data bearing different letters were significantly different (p<0.05) | |
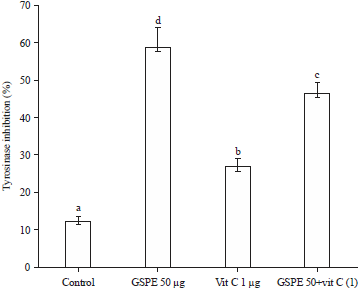 | |
| Fig. 9: | Effect of various experimental group (GSPE, Vit C and GSPE-Vit C) on tyrosinase inhibitory activity in mushroom tyrosinase in vitro model |
Values were expressed as mean±SD. Data bearing different letters were significantly different (p<0.05) | |
The GSPE alone showed the best inhibition than other groups, owing to the presence of various polyphenols (OPCs), which helps in downregulating the activation of TRP1 and 2. The GSPE could directly encounter free radical produced by melanoma cells and protect the cells from undergoing apoptosis or necrosis, which might be the reason for higher cell viability rate in GSPE group. In addition, the metal chelating activity of GSPE might change the conformational structure (3D) of copper containing tyrosinase enzymes and thereby inhibiting the activity of tyrosinase11.
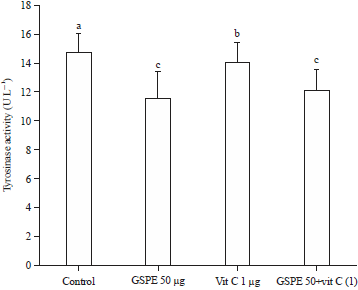 | |
| Fig. 10: | Effect of various experimental group (GSPE, Vit C and GSPE-Vit C) on tyrosinase activity in B16F1 cell model |
Values were expressed as mean±SD. Data bearing different letters were significantly different (p<0.05) | |
GSPE alone group showed superior anti-melanogenesis effect owing to the synergic or holistic effect exerted by various phyto-components (OPCs). Similarly, OPCs in GSPE had been reported to inhibit the melanin synthesis in melanocytes of the guinea pig skin via an antioxidant property10. On the basis of these findings, the crude GSPE extract might be recommended for treating hyperpigmentation disorder.
CONCLUSION
The outcome of the current study indicated that GSPE (crude) did not stimulate cytotoxicity in both UV-exposed and unexposed B16F1 cells and thus indicating its safety usage as a whitening agent. The GSPE alone group exhibited anti-melanogenesis activity by lowering oxidative stress and effectively downregulating the tyrosinase activity and thereby abolishing the melanogenesis process. However, further studies are needed to clarify the exact mechanism behind the anti-melanogenesis activity of GSPE.
SIGNIFICANCE STATEMENT
This in vitro study exhibited that crude extract of GSPE possesses potent anti-melanogenesis properties by suppressing free radical generation and tyrosinase activity. Therefore, GSPE can be a promising contender for treating hyperpigmentation disorder as well as recommended as a whitening agent in cosmetic industries.
ACKNOWLEDGMENT
We are thankful to the National Science Council, Taiwan (NSC 90-2313-B040-004) for funding this study.
REFERENCES
- Hearing, V.J., 2005. Biogenesis of pigment granules: A sensitive way to regulate melanocyte function. J. Dermatol. Sci., 37: 3-14.
CrossRefDirect Link - Wang, S.Q., R. Setlow, M. Berwick, D. Polsky, A.A. Marghoob, A.W. Kopf and R.S. Bart, 2001. Ultraviolet A and melanoma: A review. J. Am. Acad. Dermatol., 44: 837-846.
CrossRefPubMedDirect Link - Yoon, W.J., M.J. Kim, H.B. Koh, W.J. Lee, N.H. Lee and C.G. Hyun, 2010. Effect of korean red sea cucumber (Stichopus japonicus) on melanogenic protein expression in murine B16 melanoma. Int. J. Pharmacol., 6: 37-42.
CrossRefDirect Link - Nichols, J.A. and S.K. Katiyar, 2010. Skin photoprotection by natural polyphenols: Anti-inflammatory, antioxidant and DNA repair mechanisms. Arch. Dermatol. Res., 302: 71-83.
CrossRefDirect Link - Panich, U., V. Tangsupa-a-nan, T. Onkoksoong, K. Kongtaphan, K. Kasetsinsombat, P. Akarasereenont and A. Wongkajornsilp, 2011. Inhibition of UVA-mediated melanogenesis by ascorbic acid through modulation of antioxidant defense and nitric oxide system. Arch. Pharm. Res., 34: 811-820.
CrossRefDirect Link - Shoji, T., S. Masumoto, N. Moriichi, M. Kobori, T. Kanda, H. Shinmoto and T. Tsushida, 2005. Procyanidin trimers to pentamers fractionated from apple inhibit melanogenesis in B16 mouse melanoma cells. J. Agric. Food Chem., 53: 6105-6111.
CrossRefDirect Link - Lan, C.Z., L. Ding, Y.L. Su, K. Guo and L. Wang et al., 2015. Grape seed proanthocyanidins prevent DOCA-salt hypertension-induced renal injury and its mechanisms in rats. Food Funct., 6: 2179-2186.
CrossRefDirect Link - El-Beshbishy, H.A., A.M. Mohamadin, A.A. Nagy and A.B. Abdel-Naim, 2010. Amelioration of tamoxifen-induced liver injury in rats by grape seed extract, black seed extract and curcumin. Indian J. Exp. Biol., 48: 280-288.
PubMedDirect Link - Yin, W., B. Li, X. Li, F. Yu, Q. Cai, Z. Zhang and H. Gao, 2015. Anti-inflammatory effects of grape seed procyanidin B2 on a diabetic pancreas. Food Funct., 6: 3065-3071.
CrossRef - Yamakoshi, J., F. Otsuka, A. Sano, S. Tokutake, M. Saito, M. Kikuchi and Y. Kubota, 2003. Lightening effect on ultraviolet-induced pigmentation of guinea pig skin by oral administration of a Proanthocyanidin rich extract from grape seeds. Pigment Cell Melanoma Res., 16: 629-638.
CrossRefDirect Link - Zi, S.X., H.J. Ma, Y. Li, W. Liu, Q.Q. Yang, G. Zhao and S. Lian, 2009. Oligomeric proanthocyanidins from grape seeds effectively inhibit ultraviolet-induced melanogenesis of human melanocytes in vitro. Int. J Mol. Med., 23: 197-204.
CrossRefDirect Link - Choi, Y.K., Y.K. Rho, K.H. Yoo, Y.Y. Lim and K. Li et al., 2010. Effects of vitamin C vs. multivitamin on melanogenesis: Comparative study in vitro and in vivo. Int. J. Dermatol., 49: 218-226.
CrossRefDirect Link - Neitmann, M., M. Alexander, J. Brinckmann, P. Schlenke and M. Tronnier, 1999. Attachment and chemotaxis of melanocytes after ultraviolet irradiation in vitro. Br. J Dermatol., 141: 794-801.
CrossRef - Bai, J. and A.I. Cederbaum, 2003. Catalase protects HepG2 cells from apoptosis induced by dna-damaging agents by accelerating the degradation of p53. J. Biol. Chem., 278: 4660-4667.
CrossRefDirect Link - Rosenthal, M.H., J.W. Kreider and R. Shiman, 1973. Quantitative assay of melanin in melanoma cells in culture and in tumors. Anal. Biochem., 56: 91-99.
CrossRefDirect Link - Yagi, A., T. Kanbara and N. Morinobu, 1987. Inhibition of mushroom-tyrosinase by aloe extract. Planta Med., 53: 515-517.
CrossRef - Kojima, S., H. Yamaguchi, K. Morita and Y. Ueno, 1995. Inhibitory effect of sodium 5, 6-benzylidene ascorbate (SBA) on the elevation of melanin biosynthesis induced by ultraviolet-A (UV-A) light in cultured B-16 melanoma cells. Biol. Pharm. Bull., 18: 1076-1080.
Direct Link - Shibahara, S., K. Takeda, K.I. Yasumoto, T. Udono, K.I. Watanabe, H. Saito and K. Takahashi, 2001. Microphthalmia-associated transcription factor (MITF): Multiplicity in structure, function and regulation. J. Invest. Dermatol., 6: 99-104.
CrossRefDirect Link - Lin, C.B., L. Babiarz, F. Liebel, M. Kizoulis and G.J. Gendimenico et al., 2002. Modulation of microphthalmia-associated transcription factor gene expression alters skin pigmentation. J. Invest. Dermatol., 119: 1330-1340.
CrossRefDirect Link - Sun, Y., C. Xiu, W. Liu, Y. Tao, J. Wang and Y.I. Qu, 2016. Grape seed proanthocyanidin extract protects the retina against early diabetic injury by activating the Nrf2 pathway. Exp. Ther. Med., 11: 1253-1258.
CrossRefDirect Link