
Dongyun Yin
Department of Cardiology, Wenling Hospital of Wenzhou Medical University, Wenling, 317500 Zhejiang, China
Qilin Wan
Dapartment of Cardiology, Huaihe Hospital of Henan University, Kaifeng, 475000 Henan, China
Qunhui Ye
Dapartment of Cardiology, Huaihe Hospital of Henan University, Kaifeng, 475000 Henan, China
Yongzhi Deng
Department of Cardiovascular Surgery, The Cardiovascular Hospital of Shanxi Medical University, Provincial Cardiovascular Hospital Shanxi, Taiyuan, 030024 Shanxi, China
Bin Lin
Department of Cardiology, Wenling Hospital of Wenzhou Medical University, Wenling, 317500 Zhejiang, China
LiveDNA: 86.9830
Lin Wu
Department of Cardiology, Wenling Hospital of Wenzhou Medical University, Wenling, 317500 Zhejiang, China
LiveDNA: 86.25211
International Journal of Pharmacology
Year: 2019 | Volume: 15 | Issue: 4 | Page No.: 503-508







ABSTRACT
Background and Objective: Visfatin is an adipokine which is abundantly expression in visceral adipose tissue. It has insulin-like effect and hypoglycemic effect. Visfatin can induce the production of inflammatory factors and myocardial hypertrophy but the exact molecular mechanisms remain unclear. The purpose of this study was to investigate the underlying molecular mechanism that visfatin induces cardiomyocyte hypertrophy. Materials and Methods: H9c2 cells were cultured and treated with visfatin. Cell surface area was measured with ImageJ software. Protein synthesis was calculated by dividing the protein content by the cell number. BNP, α-SMA, p-ERK1/2 and ERK1/2 protein expression were detected by Western blotting. Results: The results showed that visfatin induced cell surface area, protein synthesis, BNP, α-SMA and p-ERK1/2 protein expression increase significantly. Pre-treatment with MAPK Kinase Inhibitor PD98059 of ERK1/2 signaling pathway significantly attenuated these effects of visfatin. Conclusion: The results suggested that visfatin induces cardiomyocyte hypertrophy through the activation of ERK1/2 signaling pathway.
PDF Abstract XML References Citation
How to cite this article
Dongyun Yin, Qilin Wan, Qunhui Ye, Yongzhi Deng, Bin Lin and Lin Wu, 2019. Cardiomyocyte Hypertrophy induced by Visfatin in H9c2 Embryonic Rat Cardiac Cells via ERK1/2 Signaling Pathway. International Journal of Pharmacology, 15: 503-508.
DOI: 10.3923/ijp.2019.503.508
URL: https://scialert.net/abstract/?doi=ijp.2019.503.508
DOI: 10.3923/ijp.2019.503.508
URL: https://scialert.net/abstract/?doi=ijp.2019.503.508
INTRODUCTION
A rapid rise in the rate of cardiovascular diseases is related to the obesity, which obesity occurs mostly due to the excessive increased adipose tissue1. Adipose tissue is an active endocrine organ which can secrete a variety of biological active substances known as adipokines2,3. Visfatin is an adipokine that is closely related to cardiovascular diseases. The levels of serum visfatin are helpful for early diagnosis of coronary heart disease4. The plasma levels of visfatin are reported to be higher in subjects with type 2 diabetes mellitus. Visfatin exerts a detrimental effect on diabetic nephropathy via the activation of the renin-angiotensin system5. Visfatin cause unstable plaque and then cause the occurrence of acute myocardial infarction6,7. In addition, visfatin promotes the expression of fibrosis proteins by activating the local renin angiotensin system in the kidney, which can be speculated that it may have a role in fibrosis5. Yu et al.8 also found that visfatin may cause fibrosis, which leads to the occurrence of heart failure. Recent studies suggest that visfatin is involved in the pathogenesis of cardiac hypertrophy and heart failure9-11.
A large number of studies have shown that ERK1/2 signaling pathway participates in cardiac hypertrophy, remodeling and fibrosis in the pathogenesis12-14. ERK1/2 signaling pathway is also found to be related to atherosclerosis. The specific inhibitors of ERK1/2 signaling pathway block myocardial cell growth caused by endothelin-1 and phenylephrine15. In Raf deficient mice, the expression of ERK1/2 is significantly inhibited, which obviously reduces myocardial hypertrophy induced16 by TAC.
Although previous studies reported that visfatin induces cardiac hypertrophy but the underlying molecular mechanisms remain poorly defined. In this study, the changes of myocardial cell surface area, protein synthesis, BNP, α-SMA and p-ERK1/2 protein expression were observed. The aim of this study was to investigate whether visfatin induces myocardial hypertrophy via the activation of ERK1/2 signaling pathway.
MATERIALS AND METHODS
Reagents: This study was conducted from July, 1, 2015 to July, 1, 2017 in the Central Laboratory of Huaihe Hospital of Henan University. Visfatin was bought from PeproTech (Catalog number, 130-09, Rocky Hill, NJ, USA). PD98059 was ordered from Cayman (Catalog number, 10006726, Ann Arbor, MI, USA). The FBS (Sijiqing) was ordered from Zhejiang Tianhang Biological Polytron Technologies Inc. (Huzhou, Zhejiang, China). Rat cardiac muscle cell line H9c2 was ordered from American Type Culture Collection (Manassas, VA, USA). High glucose DMEM was ordered from GE Healthcare Life Sciences (Catalog number, SH30022.01, Logan, Utah, USA). RIPA lysis buffer was bought from Beyotime (Catalog number, P0013B, Beijing, China). BeyoECL Plus was bought from Beyotime (Catalog number, P0018, Beijing, China). BCA protein assay kit was bought from CWBIO (Catalog number, CW0014S, Beijing, China). About 0.45 μm of PVDF membranes was purchased from EMD Millipore (Catalog number, HVHP14250, Billerica, MA, USA). p-ERK1/2 was purchased from Cell Signaling Technology (Catalog number, 9101S, Danvers, MA, USA). T-ERK1/2 was purchased from Beyotime (Catalog number, AM076, Beijing, China). BNP (Q-16) was purchased from Santa Cruz Biotechnology Inc. (Catalog number, sc-18817, Dallas, TX, USA). α-SMA (EPR5368) was purchased from abcam (Catalog number, ab124964, Cambridge, MA, USA). β-actin (I102) was ordered from Bioworld (Catalog number, AP0060, Nanjing, China). Goat anti-Rabbit IgG (H+L) was obtained from Jackson (Catalog number, 111-035-003, West Grove, PA, USA). Donkey anti-Goat IgG (H+L) was bought from Sangon Biotech Co., Ltd., (Catalog number, D110115-0100, Shanghai, China).
Cell culture: The H9c2 rat cardiomyoblast cells were seeded into 10 cm dishes and cultured in DMEM medium containing 10% FBS, 1% penicillin and 1% streptomycin at a temperature of 37°C with a 5% CO2 atmosphere. Cells were assigned to three groups: Control, visfatin and PD98059+visfatin. The medium was changed every 2-3 days. Cells were grown up to about 70-80% and passaged to 3.5 cm dishes and then cultured for 24 h. Cells were changed with serum-free medium for 24 h and administered with visfatin for indicated time.
Determination of cell surface area: The 6×104 cells were seeded into 3.5 cm dishes. Cells were pre-treated17 with PD98059 at 50 μM for 2 h and then treated with visfatin18 at 100 ng mL–1 for 48 h. Cells were used to observe morphology under the inverted microscope by randomly selected 5 horizons of photographs per dish, each field arbitrarily selected 10 of cells. Cell surface area was determined using NIH ImageJ software (NIH, Bethesda, MD, USA)19,20.
Determination of protein synthesis: The 1×105 cells were seeded into 3.5 cm culture dishes. After cells were pre-treated with PD98059 at 50 μM for 2 h, cells were treated with visfatin at 100 ng mL–1 for 48 h. Cells were then trypsined and centrifuged. About 2 mL of fresh DMEM medium containing 10% FBS was added to the supernatant and the cell suspension was made into a single cell suspension. About 20 μL of cell suspension was mixed with 20 μL of 0.2% of the trypan blue solution and 20 μL of mixture was added to the automatic counting board to count cell number using Countstar counter. Each hole was counted 3 fields of view at least 5 times. The remaining cells were lysed with RIPA lysis buffer and protein concentration was measured with the BCA protein assay kit. Cell protein synthesis was determined by the protein content divided by the cell number19,21.
Western blot analysis: Cells were lyses with RIPA lysis buffer and protein concentration was quantified with the BCA protein assay kit. The loading samples were 10-120 μg (BNP, 120 μg, α-SMA, ERK1/2, p-ERK1/2, 10 μg). The electrophoresis for α-SMA, BNP, ERK1/2 and p-ERK1/2 was used with 10% denatured SDS-PAGE gels. The proteins were transferred to 0.45 μm PVDF membranes and the membranes were blocked with 5% non-fat milk in TBST buffer at room temperature (BNP, 2 h; p-ERK1/2, α-SMA, ERK1/2 and β-actin 1 h). The primary antibodies (BNP, 1:100, α-SMA, 1:50000, ERK1/2, 1:2000, p-ERK1/2, 1:2000 and β-actin, 1:5000) were diluted with 5% non-fat milk TBST buffer and added to the membranes shaking at 4°C overnight. The secondary antibodies were diluted to 1:5000 and incubated with membranes at room temperature for 1 h. The bands were detected by BeyoECL Plus and analyzed with the NIH ImageJ software. The ratio of phosphorylated protein antibodies over corresponding total protein antibodies was calculated.
Statistical analysis: All experiment data were expressed as means±SD. The difference between the two groups was analyzed by using independent samples t-test and the SPSS17.0 statistical software was used for the analysis of the experimental data. The difference was considered statistically significant if p<0.05.
RESULTS
Visfatin induces cardiomyocyte hypertrophy that is inhibited by PD98059: In order to observe the effects of visfatin and PD98059 inhibitor of ERK1/2 signaling pathway on cardiomyocyte hypertrophy, cells were pre-treated with PD98059 and then stimulated with visfatin. The results showed that visfatin induced H9c2 cell surface area increase significantly. After pre-treatment of PD98059, cell surface area was decreased significantly as compared with the visfatin group (Fig. 1a). Visfatin induced protein synthesis increase, while PD98059 pre-treatment caused protein synthesis reduction significantly (Fig. 1b). Furthermore visfatin induced α-SMA and BNP protein expression increase obviously, whereas, PD98059 pre-treatment caused α-SMA (Fig. 1c, d) and BNP (Fig. 1e, f) protein expression decrease significantly compared to the visfatin group.
Visfatin increases phosphorylation of ERK1/2 protein that is decreased by PD98059: The inhibitor PD98059 of ERK1/2 signaling pathway was used to treat cells and visfatin was again employed. The results displayed that visfatin induced p-ERK1/2 protein expression increase significantly. After pre-treatment with PD98059, p-ERK1/2 protein expression was significantly decreased compared to the visfatin group (Fig. 2a, b) suggesting that visfatin can activate ERK1/2 signaling pathway in cardiomyocytes.
DISCUSSION
Myocardial hypertrophy is a basic adaptive response to the hemodynamic overload caused by various kinds of cardiac muscle, which is mainly characterized by the increase of intracellular protein synthesis and the change of cell volume. The experiment results showed: (1) Visfatin induced cardiomyocyte hypertrophy, (2) Visfatin activated ERK1/2 signaling pathway in cardiac myocytes and (3) Inhibition of ERK1/2 signaling pathway attenuated visfatin-induced cardiomyocyte hypertrophy.
Visfatin was discovered by Fukuhara et al.22, which is a cDNA fragment identical to the 5' non-coding region of PBEF (pre-B cell colony-enhancing factor). Visfatin was subsequently found in human and mouse abdominal visceral fat. So it is called visfatin23,24. Some studies have indicated that visfatin has a close relationship with atherosclerosis, cardiac hypertrophy and acute myocardial infarction10,25. Visfatin induces cardiac hypertrophy via activation of MAPK pathway. The amount of visfatin is significantly increased in the plasma of patients with acute heart failure, which is positively correlated with NT-proBNP25. In the experiments, visfatin induced increased myocyte surface area and protein synthesis, enhanced hypertrophic marker BNP19 and α-SMA26 protein expression, which are consistent with previous studies27.
Many studies have shown that ERK1/2 signaling pathway is involved in the cardiac hypertrophy28-30. Yu et al.8 showed that visfatin can cause myocardial fibrosis via the activation of p38MAPK and ERK1/2 pathways. Kim et al.31,32 found that visfatin not only promotes the migration of human umbilical vein endothelial cells (HUVECs) by activating ERK1/2 signaling pathway but also accelerates the formation of new blood vessels.
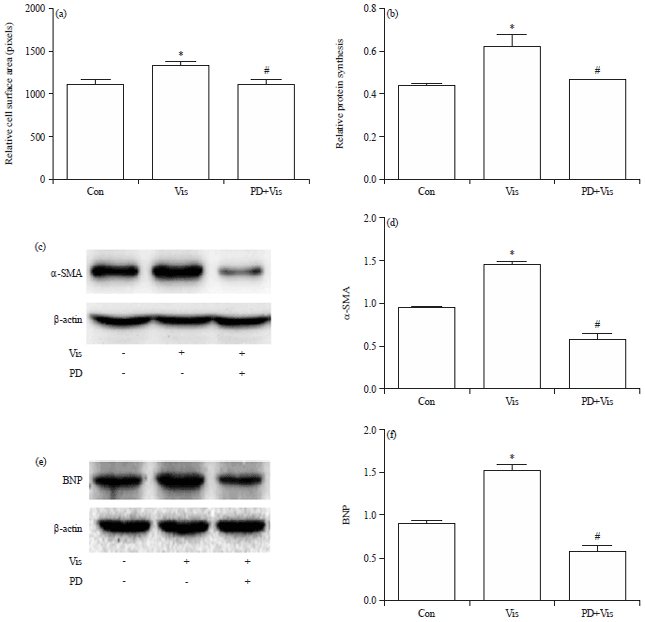 | |
| Fig. 1(a-f): | PD98059 attenuates visfatin-induced cardiomyocyte hypertrophy, (a) Measurement of cell surface area, (b) Changes of protein synthesis in H9c2 cells, (c, e) Western blot analysis of α-SMA and BNP protein expression and (d, f) Quantitative results of α-SMA and BNP protein expression |
| *p<0.05 vs. the control group, #p<0.05 vs. the visfatin group, Con: Control group, Vis: Visfatin group, PD+Vis: PD98059+visfatin group | |
 | |
| Fig. 2(a-b): | PD98059 decreases p-ERK1/2 that is increased by visfatin in cardiomyocytes, (a) Western blot analysis of p-ERK1/2 protein expression and (b) Quantification of p-ERK1/2 protein expression |
| *p<0.05 vs. the control group, #p<0.05 vs. the visfatin group, Con: Control group, Vis: Visfatin group, PD+Vis: PD98059+visfatin group | |
PD98059 is a specific inhibitor of the ERK1/2 signaling pathway, which exerts its effect by inhibiting the upstream kinase27 of ERK1/2. The experiments showed that visfatin causes increased phospho-ERK1/2 protein expression, which is decreased by PD98059. PD98059 could significantly reduce myocardial cell surface area and protein synthesis. Western blotting also showed that PD98059 decreased cardiomyocyte BNP and α-SMA protein expression, suggesting that ERK1/2 signaling pathway is involved in the regulation of myocardial hypertrophy.
CONCLUSION
In general, the experiments displayed that visfatin induced cardiomyocyte hypertrophy, increased ERK1/2 signaling pathway. The inhibitor of ERK1/2 signaling pathway could inhibit visfatin-induced cardiomyocyte hypertrophy. These results suggested that visfatin induces cardiomyocyte hypertrophy by the activation of ERK1/2 signaling pathway. This will undoubtedly provide a new target for the treatment of cardiac hypertrophy.
SIGNIFICANCE STATEMENT
This study showed that visfatin induces cardiomyocyte hypertrophy via ERK1/2 signaling pathway. It will help the researchers to explore the underlying molecular mechanisms of visfatin-induced cardiomyocyte hypertrophy and provides the new insight that inhibition of visfatin expression may be beneficial for treatment of cardiac hypertrophy.
ACKNOWLEDGMENTS
This work was supported by the Key Research and Development Program of Shanxi Province, China (Grant No. 201603D321057), Scientific Research Foundation of Health and Family Planning Commission of Shanxi Province, China (Grant No. 201601093), Natural Science Foundation of Shanxi, China (Grant No: 201701D121162), Zhejiang Medical and Health Research Project, China (Grant No: 2018KY915) and Wenling Science and Technology Project, China (Grant No: 2017C311105).
REFERENCES
- Kahn, B.B. and J.S. Flier, 2000. Obesity and insulin resistance. J. Clin. Invest., 106: 473-481.
CrossRefDirect Link - MacDougald, O.A. and C.F. Burant, 2007. The rapidly expanding family of adipokines. Cell Metab., 6: 159-161.
CrossRefDirect Link - Cao, H., K. Gerhold, J.R. Mayers, M.M. Wiest, S.M. Watkins and G.S. Hotamisligil, 2008. Identification of a lipokine, a lipid hormone linking adipose tissue to systemic metabolism. Cell, 134: 933-944.
CrossRefDirect Link - Takebayashi, K., M. Suetsugu, S. Wakabayashi, Y. Aso and T. Inukai, 2007. Association between plasma visfatin and vascular endothelial function in patients with type 2 diabetes mellitus. Metabolism, 56: 451-458.
CrossRefDirect Link - Huang, Q., Y. Guo, H. Zeng, W. Xie, H. Yan and H. Ding, 2011. Visfatin stimulates a cellular renin-angiotensin system in cultured rat mesangial cells. Endocr. Res., 36: 93-100.
CrossRefDirect Link - Dahl, T.B., A. Yndestad, M. Skjelland, E. Oie and A. Dahl et al., 2007. Increased expression of visfatin in macrophages of human unstable carotid and coronary atherosclerosis: Possible role in inflammation and plaque destabilization. Circulation, 115: 972-980.
CrossRefDirect Link - Zhong, M., H.W. Tan, H.P. Gong, S.F. Wang, Y. Zhang and W. Zhang, 2008. Increased serum visfatin in patients with metabolic syndrome and carotid atherosclerosis. Clin. Endocrinol., 69: 878-884.
CrossRefDirect Link - Yu, X.Y., S.B. Qiao, H.S. Guan, S.W. Liu and X.M. Meng, 2010. Effects of visfatin on proliferation and collagen synthesis in rat cardiac fibroblasts. Hormone Metab. Res., 42: 507-513.
CrossRefPubMedDirect Link - Pillai, V.B., N.R. Sundaresan, G. Kim, S. Samant, L. Moreno-Vinasco, J.G. Garcia and M.P. Gupta, 2012. Nampt secreted from cardiomyocytes promotes development of cardiac hypertrophy and adverse ventricular remodeling. Am. J. Physiol.-Heart Circ. Physiol., 304: H415-H426.
CrossRefDirect Link - Chang, L., R. Yang, M. Wang, J. Liu, Y. Wang, H. Zhang and Y. Li, 2012. Angiotensin II type-1 receptor-JAK/STAT pathway mediates the induction of visfatin in angiotensin II-induced cardiomyocyte hypertrophy. Am. J. Med. Sci., 343: 220-226.
CrossRefDirect Link - Yang, R., L. Chang, S. Liu, X. Jin and Y. Li, 2014. High glucose induces Rho/ROCK-dependent visfatin and type I procollagen expression in rat primary cardiac fibroblasts. Mol. Med. Rep., 10: 1992-1998.
CrossRefDirect Link - Zong, J., F.F. Li, K. Liang, R. Dai and H. Zhang et al., 2018. Nuclear localization leucine-rich-repeat protein 1 deficiency protects against cardiac hypertrophy by pressure overload. Cell. Physiol. Biochem., 48: 75-86.
CrossRefDirect Link - Bao, M.W., X.J. Zhang, L. Li, Z. Cai and X. Liu et al., 2015. Cardioprotective role of growth/differentiation factor 1 in post-infarction left ventricular remodelling and dysfunction. J. Pathol., 236: 360-372.
CrossRefDirect Link - Wang, L.P., X.H. Yang, X.J. Wang, S.M. Li, N. Sun and T. Zhang, 2016. Erythropoietin decreases the occurrence of myocardial fibrosis by inhibiting the NADPH-ERK-NF-κB pathway. Cardiology, 133: 97-108.
CrossRefDirect Link - Yue, T.L., J.L. Gu, C. Wang, A.D. Reith and J.C. Lee et al., 2000. Extracellular signal-regulated kinase plays an essential role in hypertrophic agonists, endothelin-1 and phenylephrine-induced cardiomyocyte hypertrophy. J. Biol. Chem., 275: 37895-37901.
CrossRefDirect Link - Harris, I.S., S. Zhang, I. Treskov, A. Kovacs, C. Weinheimer and A.J. Muslin, 2004. Raf-1 kinase is required for cardiac hypertrophy and cardiomyocyte survival in response to pressure overload. Circulation, 110: 718-723.
CrossRefPubMedDirect Link - Cerioni, L., L. Palomba and O. Cantoni, 2003. The Raf/MEK inhibitor PD98059 enhances ERK1/2 phosphorylation mediated by peroxynitrite via enforced mitochondrial formation of reactive oxygen species. FEBS Lett., 547: 92-96.
CrossRefDirect Link - Buldak, R.J., R. Polaniak, L. Buldak, L. Mielanczyk and M. Kukla et al., 2013. Exogenous administration of visfatin affects cytokine secretion and increases oxidative stress in human malignant melanoma Me45 cells. J. Physiol. Pharmacol., 64: 377-385.
PubMedDirect Link - Liu, P., G.C. Cheng, Q.H. Ye, Y.Z. Deng and L. Wu, 2016. Lkb1/ampk pathway mediates resistin-induced cardiomyocyte hypertrophy in h9c2 embryonic rat cardiomyocytes. Biomed. Rep., 4: 387-391.
CrossRefDirect Link - Hernandez, J.S., G. Barreto-Torres, A.V. Kuznetsov, Z. Khuchua and S. Javadov, 2014. Crosstalk between AMPK activation and angiotensin II-induced hypertrophy in cardiomyocytes: The role of mitochondria. J. Cell. Mol. Med., 18: 709-720.
CrossRefDirect Link - Tu, V.C., J.J. Bahl and Q.M. Chen, 2002. Signals of oxidant-induced cardiomyocyte hypertrophy: Key activation of p70 S6 kinase-1 and phosphoinositide 3-kinase. J. Pharmacol. Exp. Therapeut., 300: 1101-1110.
CrossRefDirect Link - Fukuhara, A., M. Matsuda, M. Nishizawa, K. Segawa and M. Tanaka et al., 2005. Visfatin: A protein secreted by visceral fat that mimics the effects of insulin. Science, 307: 426-430.
CrossRefDirect Link - Moschen, A.R., A. Kaser, B. Enrich, B. Mosheimer, M. Theurl, H. Niederegger and H. Tilg, 2007. Visfatin, an adipocytokine with proinflammatory and immunomodulating properties. J. Immunol., 178: 1748-1758.
CrossRefDirect Link - McGlothlin, J.R., L. Gao, T. Lavoie, B.A. Simon and R.B. Easley et al., 2005. Molecular cloning and characterization of canine pre-B-cell colony-enhancing factor. Biochem. Genet., 43: 127-141.
CrossRefDirect Link - Schulze, P.C., A. Biolo, D. Gopal, K. Shahzad and J. Balog et al., 2011. Dynamics in insulin resistance and plasma levels of adipokines in patients with acute decompensated and chronic stable heart failure. J. Cardiac Failure, 17: 1004-1011.
CrossRefDirect Link - Trivedi, C.M., Y. Luo, Z. Yin, M. Zhang and W. Zhu et al., 2007. Hdac2 regulates the cardiac hypertrophic response by modulating Gsk3β activity. Nat. Med., 13: 324-331.
CrossRefDirect Link - Pannu, J., S. Nakerakanti, E. Smith, P. ten Dijke and M. Trojanowska, 2007. Transforming growth factor-β receptor type I-dependent fibrogenic gene program is mediated via activation of Smad1 and ERK1/2 pathways. J. Biol. Chem., 282: 10405-10413.
CrossRefDirect Link - Sbroggio, M., A. Bertero, S. Velasco, F. Fusella and E. De Blasio et al., 2011. ERK1/2 activation in heart is controlled by melusin, focal adhesion kinase and the scaffold protein IQGAP1. J. Cell Sci., 124: 3515-3524.
CrossRefDirect Link - Bueno, O.F., L.J. De Windt, K.M. Tymitz, S.A. Witt and T.R. Kimball et al., 2000. The MEK1-ERK1/2 signaling pathway promotes compensated cardiac hypertrophy in transgenic mice. EMBO J., 19: 6341-6350.
CrossRefDirect Link - Purcell, N.H., B.J. Wilkins, A. York, M.K. Saba-El-Leil, S. Meloche, J. Robbins and J.D. Molkentin, 2007. Genetic inhibition of cardiac ERK1/2 promotes stress-induced apoptosis and heart failure but has no effect on hypertrophy in vivo. Proc. Natl. Acad. Sci. USA., 104: 14074-14079.
CrossRefDirect Link - Kim, S.R., S.K. Bae, K.S. Choi, S.Y. Park and H.O. Jun et al., 2007. Visfatin promotes angiogenesis by activation of extracellular signal-regulated kinase 1/2. Biochem. Biophys. Res. Commun., 357: 150-156.
CrossRefDirect Link - Kim, M., J.K. Oh, S. Sakata, I. Liang, W. Park, R.J. Hajjar and D. Lebeche, 2008. Role of resistin in cardiac contractility and hypertrophy. J. Mol. Cell. Cardiol., 45: 270-280.
CrossRefDirect Link