
Nagla Abd El-Aziz El-Shitany
Department of Pharmacology and Toxicology, Faculty of Pharmacy, King Abdulaziz University, Jeddah, Saudi Arabia
LiveDNA: 20.23565
Aymn Tallat Abbas
Special Infectious Agents Unit, King Fahd Medical Research Center, King Abdulaziz University, Jeddah, Saudi Arabia
Soad Shaker Ali
Department of Anatomy Cytology and Histology, Faculty of Medicine, King Abdulaziz University, Jeddah, Saudi Arabia
LiveDNA: 20.23572
Basma Eid
Department of Pharmacology and Toxicology, Faculty of Pharmacy, King Abdulaziz University, Jeddah, Saudi Arabia
LiveDNA: 966.23486
Steve Harakeh
Special Infectious Agents Unit, King Fahd Medical Research Center, King Abdulaziz University, Jeddah, Saudi Arabia
Thikryat Neamatallah
Department of Pharmacology and Toxicology, Faculty of Pharmacy, King Abdulaziz University, Jeddah, Saudi Arabia
Ahmed Al-Abd
Department of Pharmacology and Toxicology, Faculty of Pharmacy, King Abdulaziz University, Jeddah, Saudi Arabia
LiveDNA: 20.899
Shaker Mousa
Department of Anatomy Cytology and Histology, Faculty of Medicine, King Abdulaziz University, Jeddah, Saudi Arabia
International Journal of Pharmacology
Year: 2019 | Volume: 15 | Issue: 4 | Page No.: 465-477







ABSTRACT
Background and Objective: Ellagic acid (ELL) is a phenolic constituent of natural sources with antioxidant and anti-inflammatory activities. The ELL has a low bio-availability, therefore a nanoparticles (NP ELL) was formulated to determine whether NP ELL could alleviate cisplatin (CISP)-induced hepatotoxicity in comparison to a high dose of regular ELL. Materials and Methods: The NP ELL was prepared using an emulsion solvent diffusion technique. Hepatotoxicity was induced in rats using CISP (7.5 mg kg–1). Rats were pre-treated with either 10 ELL (10 mg kg–1), 1 NP ELL (1 mg kg–1) or 2 NP ELL (2 mg kg–1). Results: All ELL regimens significantly decreased CISP-induced increases in liver enzymes, MDA, NO, TNF-α, NFkB and BAX. Additionally, all significantly restored hepatic GSH, GPx, CAT and Bcl-2. Rat’s liver showed no injury in all ELL pretreated groups. Importantly, NP ELL did not inhibit the cytotoxic effect CISP against HCT116 cell lines. Conclusion: The NP ELL has improved bio-availability and protected against CISP induced hepatotoxicity. This provides an encouraging way of ameliorating the bio-availability of ELL while sustaining its desirable therapeutic effects.
PDF Abstract XML References Citation
Received: December 05, 2018;
Accepted: February 08, 2019;
Published: April 15, 2019
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Nagla Abd El-Aziz El-Shitany, Aymn Tallat Abbas, Soad Shaker Ali, Basma Eid, Steve Harakeh, Thikryat Neamatallah, Ahmed Al-Abd and Shaker Mousa, 2019. Nanoparticles Ellagic Acid Protects Against Cisplatin-induced Hepatotoxicity in Rats Without Inhibiting its Cytotoxic Activity. International Journal of Pharmacology, 15: 465-477.
DOI: 10.3923/ijp.2019.465.477
URL: https://scialert.net/abstract/?doi=ijp.2019.465.477
DOI: 10.3923/ijp.2019.465.477
URL: https://scialert.net/abstract/?doi=ijp.2019.465.477
INTRODUCTION
Cisplatin (CISP) is cytotoxic agent that is used in the treatment of solid tumors, such as: Ovarian, bladder and testicular tumors1,2. However, its known nephrotoxicity, hepatotoxicity and neurotoxicity have hampered its clinical use in oncology3-5. Hepatotoxicity occurs at high doses due to the accumulation of cisplatin in liver cells, this results in inflammation, overproduction of free radicals and a decrease in the antioxidant enzymes6,7. The generated oxidative stress appears to be an important mechanism in liver dysfunction8. Studies have shown that CISP increases lipid peroxidation and decreases glutathione (GSH) levels in hepatic tissues9,10. It also elevates levels of hydroxyl (–OH), peroxynitrite, superoxide-anion (O2–) and hydrogen-peroxide radicals11,12. CISP-triggered hepatic immune events are indicated by increased tumor necrosis factor-α (TNF-α) and decreased interleukin-10 (IL-10)13. During CISP-induced toxicity, levels of Bax (a pro-apoptotic protein) have been shown to increase, while levels of Bcl-2 (an anti-apoptotic protein) have been shown to decrease13.
The protective effects of several anti-inflammatory agents and anti-oxidants against CISP-induced hepatotoxicity in experimental animals have been investigated10,14. Of interest are flavonoids-polyphenolic antioxidants that are found in many vegetables and fruits15-17. One such flavonoid, ellagic acid (ELL) is a phenolic constituent in green tea, walnuts and fruits such as grapes, berries and pomegranates. It possesses a variety of biological properties18 including antioxidant19-21 and anti-inflammatory22-24 activities. Researchers have shown that in rats’ livers, ELL attenuates cisplatin-enhanced lipid peroxidation and raises GPx and CAT levels. The ELL counteracts the detrimental effects25 of –OH and O2–. In addition, ELL has been found to decrease the level of TNF-α in the livers of rats with lipopolysaccharide-induced sepsis26. When combined with p-coumaric acid, ELL also increases cytokine Il-10 (an anti-inflammatory cytokine) in rats’ livers in the same sepsis model26.
The ELL has been also found to be useful in the retraction of several types of tumors including those in lung27, colorectal28, esophageal29 and breast cancers30. However, despite its promising properties, ELL has low solubility in water (<10 mg mL–1 in phosphate buffer pH) and permeability (0.13×10–6). In addition, it is unstable at the physiological pH31 of 7.4. When administered orally, it is poorly absorbed and metabolized in the gastrointestinal tract and is thus rapidly eliminated32,33. All of which limits the therapeutic benefits of ELL and its clinical uses34.
The utilization of nanotechnology in the field of alternative medicine is one of the fastest growing fields. The use of nanotechnology has many advantages that have led to a surge in the use of alternative medicine in the treatment of many tumors and chronic conditions. The loading of natural products on nano-particles increases their bio-availability, targeting and controlled-release pattern35. Therefore, this study was designed to: (1) Prepare ELL nanoparticles (ELL NP), (2) Determine the protective effects that both regular ELL and NP ELL formulations have on CISP-induced hepatotoxicity in rats, (3) Emphasis the possible protective mechanism of ELL against CISP-induced hepatotoxicity, (4) Examine the effect NP ELL treatment on the cytotoxic activity of CISP using human colorectal adenocarcinoma cells (HCT116).
MATERIALS AND METHODS
Synthesis of nanoparticles ellagic acid (NP ELL): In 10 mL ethanol mix 25 mg regular ELL with 150 mg of lecithin, 100 mg of stearic acid, 80 mg of cholesterol, 20 mg of DSPE-PEG. The mixture was then mixed with a solution containing 5 mL of 1% tween 80 and 5 mL of aqueous solution containing 25 mg of chitosan and was stirred for 40 min at 70°C. After that 40 mL of DI water was added and stirring for about 1 h at 70°C to evaporate the ethanol. The dialyzed product was lyophilized using 5% sucrose.
Particle size determination: A Malvern zetasizer (Malvern Instrumentation Co., Westborough, MA, USA) was used to determine the size distribution of the NP ELL in aqueous dispersions (Fig. 1).
Entrapment/loading efficiency: The quantity of ELL in the nanoparticles was determined by comparing the absorbance at 270 nm to standard curves for different ELL concentrations. The Eq. 1 was used to determine entrapment efficiency:
| (1) |
where, (Drug)f represents the the concentration of ELL in the nanoparticles. (Drug)t represents the theoretical concentration of ELL.
The entrapment efficiency was found to be around 55%. The overall loading of ELL in nanoparticles were found be around 0.84% w/w (Fig. 2).
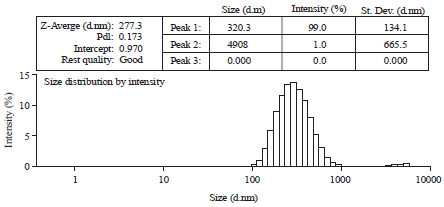 | |
| Fig. 1: | Size measurement of nanoparticles ellagic acid (NP ELL) using dynamic light scattering (DLS). Average particle size is around 277 nm in diameter, PDI = 0.173 |
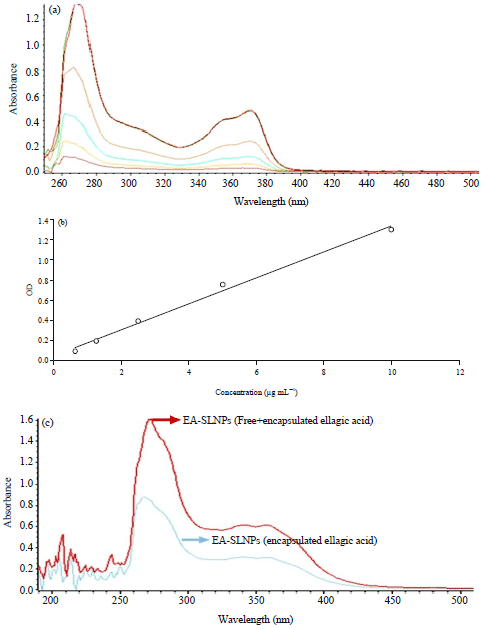 | |
| Fig. 2(a-c): | Determination of entrapment/loading efficiency of encapsulated nanoparticles ellagic acid (NP ELL), (a) UV-VIS spectra used to construct the standard curve of ELL, (b) With concentrations of PE from 0.625, 1.25, 2.5, 5 and 10 μg mL–1 and (c) Determination of entrapment efficiency by comparing OD from UV-Vis spectra of total amount ELL (free+encapsulated) and encapsulated ELL in the SLNPs |
Animals: Thirty male Sprague Dawley rats (150-180 g) were separated into five groups (n = 6). Control group (CONT): received no treatment, cisplatin group (CISP): Received a single-dose of CISP (7.5 mg kg–1) intraperitoneally (i.p.)36, regular ellagic acid group (10 E LL): Received a single dose of regular ellagic acid (10 mg kg–1)10, 6 h before CISP, 1st nanoparticle ellagic acid group (1 NP ELL): Received single dose of NP ELL (1 mg kg–1), 6 h before CISP, 2nd nanoparticle ellagic acid group (2 NP ELL): Received a single dose of NP ELL (2 mg kg–1), 6 h before CISP. The study protocol was approved (No. 1438-108) by the Scientific Research Ethics Committee of the Faculty of Pharmacy, KAU, KSA.
Sample collection: About 24 h after CISP injection rats were anesthetized with ether and blood samples were withdrawn. Serum was then separated and kept at -80°C. Rats were then killed by cervical dislocation and the livers were extracted. Parts of the livers were preserved in 10% neutral formalin solution while the remaining parts of the liver were kept at -80°C.
Liver functions measurement: Detection kits (Human, Germany) were used to determine the liver enzymes aspartate amino-transferase (AST), alanine amino-transferase (ALT) and alkaline phosphatase (ALP) according to the manufacturing instructions.
Oxidative stress markers and antioxidants enzymes measurement: Kits purchased from Abcam (USA) were used to determine the levels of malondialdehyde (MDA), nitric oxide (NO), reduced glutathione (GSH), glutathione peroxidase (GPx) and catalase (CAT) in the liver homogenates based on the manufacturer's instructions.
ELISA measurements: Nuclear factor kappa beta (NFkB), interleukin-10 (IL-10) and tumor necrosis factor alpha (TNF-α) levels were measured in the liver homogenates using ELISA kits of Abcam (USA) for NFkB, R and D Systems, Inc., kit., (USA) for TNF-α and Novex kit (USA) for IL-10. The kits use primary antibody raised against rat NFkB, IL-10 and TNF-α.
Hematoxylin and eosin staining: Hematoxylin and eosin (H and E) were used to stain 4 μm paraffin-embedded livers, which were then examined and photographed utilizing the light microscope (Nikon Eclipse TE2000-U, NIKON, Japan).
Immunohistochemical staining: Liver sections were stained utilizing an immunoperoxidase (PAP, peroxidase/ anti-peroxidase) technique using Lab Vision (Fremont, CA) NF-kB, BAX and Bcl-2 antibodies. The slides were filmed using light microscopy (Nikon Eclipse TE2000-U, NIKON, Japan) and quantified by Image J software (Image J, 1.46a, NIH, USA).
Cell culture: The HCT116 was obtained from the Vaccera (Giza, Egypt). The HCT116 cells were cultured in RPMI-1640 media containing streptomycin, penicillin and fetal bovine serum (10%). Cells were incubated at 37°C in a humidified incubator with 5% CO2.
In vitro SRB cytotoxicity assays: The cytotoxicity was tested using sulforhodamine B (SRB) assay as previously described37. Breifly, HCT116 cells were plated in 96-well plates. Serial concentrations of CISP alone or in combination with NP ELL (1 and 10 μM) was applied to the cells. Cells were incubated for 72 h and then SRB (0.4%) was added and the cells were incubated for another 10 min. The plate was then kept drying overnight and the SRB-stained cells were dissolved using Tris-HCl. About 540 nm wavelength was used to determine color intensity. The Emax model was used to analyze the dose response curve of each compound38.
Statistical analysis: Results were represented as mean±standard error (SEM). Values were statistically analyzed utilizing one-way analysis of variance (ANOVA) followed by Tukey-Kramer test (package Minitab 18, Minitab. Inc., State College PA, USA). The level of significance was settled at p<0.05.
RESULTS
Serum liver function enzymes (AST, ALT and ALP): Animals treated with CISP (7.5 mg kg–1) showed significant increase in serum AST, ALT and ALP levels compared to the CONT group (Fig. 3a-c). Animals pretreated with 10 mg kg–1 regular ELL, 1 mg kg–1 NP ELL and 2 mg kg–1 NP ELL showed significant decrease in serum AST, ALT and ALP levels compared to the CISP group (Fig. 3a-c).
Liver oxidative stress markers (MDA, NO and GSH): Animals treated with CISP showed significant increase in liver MDA and NO contents compared to the CONT group (Fig. 4a and b). Animals pretreated with ELL, 1 NP ELL and 2 NP ELL showed significant decrease in liver MDA and NO contents compared to the CISP group (Fig. 4a and b).
On the other hand, animals treated with CISP showed significant decrease in liver GSH content compared to the CONT group (Fig. 4c). Animals pretreated with ELL, 1 NP ELL and 2 NP ELL showed significant increase in liver GSH content compared to the CISP group (Fig. 4c).
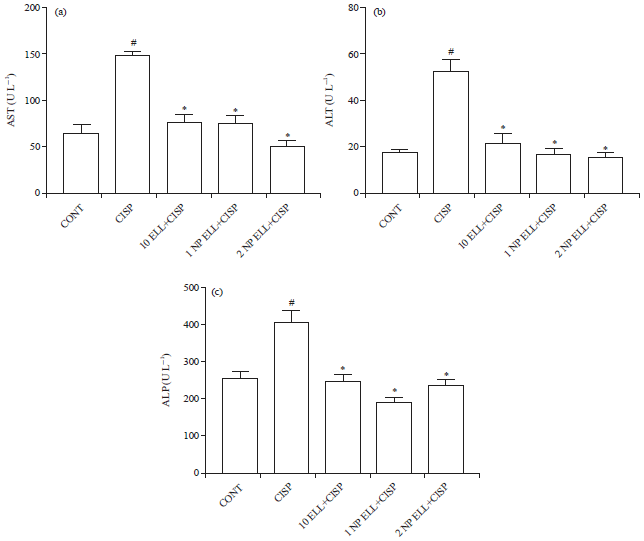 | |
| Fig. 3(a-c): | Levels of serum (a) AST, (b) ALT and (c) ALP measured in control (CONT), cisplatin (CISP, 7.5 mg kg–1), ellagic acid (10 ELL, 10 mg kg–1), nanoparticles ellagic acid (1 NP Ell, 1 mg kg–1) and nanoparticles ellagic acid (2 NP Ell, 2 mg kg–1). Data are expressed as mean±SEM of 6 rats/group. #p<0.05 compared with the CONT group. *p<0.05 compared with the CISP group |
| Data are compared using ANOVA followed by Tukey-Kramer post hoc test | |
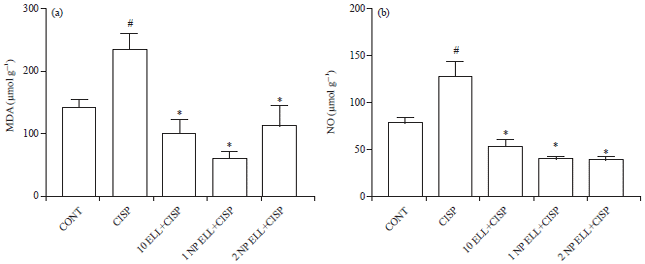 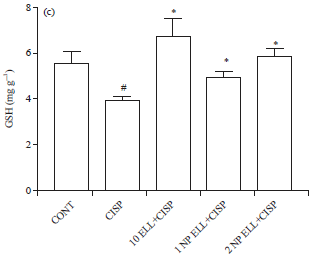 | |
| Fig. 4(a-c): | Levels of liver (a) MDA, (b) NO and (c) GSH measured in control (CONT), cisplatin (CISP, 7.5 mg kg–1), ellagic acid (10 ELL, 10 mg kg–1), nanoparticles ellagic acid (1 NP Ell, 1 mg kg–1) and nanoparticles ellagic acid (2 NP Ell, 2 mg kg–1). Data are expressed as mean±SEM of 6 rats/group. #p<0.05 compared with the CONT group. *p<0.05 compared with the CISP group |
| Data are compared using ANOVA followed by Tukey-Kramer post hoc test | |
Liver antioxidant enzymes (GPx and CAT): Animals treated with CISP showed significant decrease in liver GPx and CAT activities compared to the CONT group (Fig. 5a and b). Animals pretreated with ELL, 1 NP ELL and 2 NP ELL showed significant increase in liver GPx and CAT activities compared to the CISP group (Fig. 5a and b).
Liver cytokines (IL-10 and TNF-α): Animals treated with CISP showed significant decrease in liver IL-10 content compared to the CONT group (Fig. 6a). Animals pretreated with ELL, 1 NP ELL and 2 NP ELL showed significant increase in liver IL-10 content compared to the CISP group (Fig. 6a).
On the other hand, animals treated with CISP showed significant increase in liver TNF-α content compared to the CONT group (Fig. 6b). Animals pretreated with ELL, 1 NP ELL and 2 NP ELL showed significant decrease in liver TNF-α content compared to the CISP group (Fig. 6b).
Liver NFkB: Animals treated with CISP showed significant increase in liver NFkB content compared to the CONT group (Fig. 6c). Animals pretreated with ELL, 1 NP ELL and 2 NP ELL showed significant decrease in liver NFkB content compared to the CISP group (Fig. 6c).
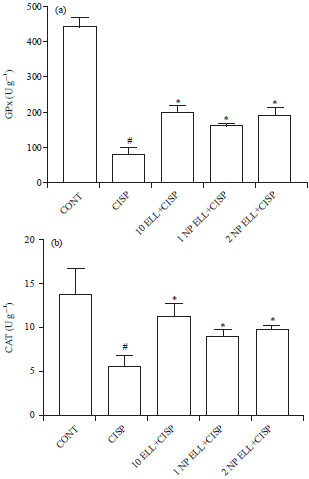 | |
| Fig. 5(a-b): | Levels of liver (a) Gpx and (c) CAT measured in control (CONT), cisplatin (CISP, 7.5 mg kg–1), ellagic acid (10 ELL, 10 mg kg–1), nanoparticles ellagic acid (1 NP Ell, 1 mg kg–1) and nanoparticles ellagic acid (2 NP Ell, 2 mg kg–1). Data are expressed as mean±SEM of 6 rats/group. #p<0.05 compared with the CONT group. *p<0.05 compared with the CISP group |
| Data are compared using ANOVA followed by Tukey-Kramer post hoc test | |
In addition, immunohistochemical expression of NFkB revealed that animals treated with CISP showed significant increase in liver NFkB expression compared to the CONT group (Fig. 7a and d). Animals pre-treated with ELL, 1 NP ELL and 2 NP ELL showed significant decrease in liver NFkB expression compared to the CISP group (Fig. 7a and d).
Liver apoptotic markers (BAX and Bcl-2): Immunohistochemical expression of BAX revealed that animals treated with CISP showed significant increase in liver BAX expression compared to the CONT group (Fig. 7b and e). Animals pretreated with ELL, 1 NP ELL and 2 NP ELL showed significant decrease in liver BAX expression compared to the CISP group (Fig. 7b and e).
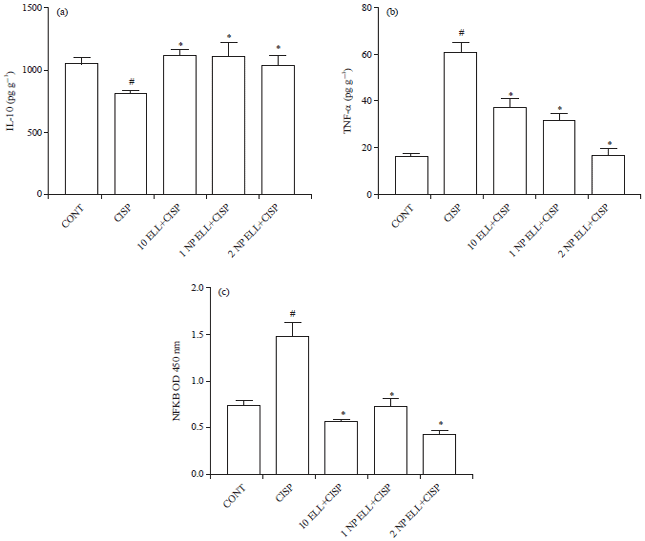 | |
| Fig. 6(a-c): | Levels of liver IL-10 (a), TNF-α (b) and NfkB (c) measured in control (CONT), cisplatin (CISP, 7.5 mg kg–1), ellagic acid (10 ELL, 10 mg kg–1), nanoparticles ellagic acid (1 NP Ell, 1 mg kg–1) and nanoparticles ellagic acid (2 NP Ell, 2 mg kg–1). Data are expressed as mean±SEM of 6 rats/group. #p<0.05 compared with the CONT group. *p<0.05 compared with the CISP group |
| Data are compared using ANOVA followed by Tukey-Kramer post hoc test | |
On the other hand, immunohistochemical expression of Bcl-2 revealed that animals treated with CISP showed significant decrease in liver Bcl-2 expression compared to the CONT group (Fig. 7c and f). Animals pretreated with ELL, 1 NP ELL and 2 NP ELL showed significant increase in liver Bcl-2 expression as compared to the CISP group (Fig. 7c and f).
Liver H and E histopathology: In Fig. 8a and b, control animals showed normal hepatocytes and blood sinusoids. In Fig. 8c and d, CISP treated animals showed swollen hepatocytes with compressing blood sinusoids, hydropic degeneration and pyknotic nuclei. In Fig. 8e and f, Ell treated animals showed normal hepatocytes, dilated sinusoids and few hepatocytes still showed dark pyknotic nuclei. In Fig. 8g and h, 1 NP ELL treated animals showed marked protection where most hepatocytes showed intact outlines, rounded vesicular active nuclei and stained cytoplasm. In Fig. 8i and j, 2 NP ELL treated animals showed that most hepatocytes are of well-defined outlines, stained cytoplasm and large vesicular active nuclei.
Anticancer activity of CISP: Pre-treatment with 1 or 10 μM NP ELL non-significantly increased the IC50 of CISP as a single agent from 1.4-1.6 μM and 2.0 μM, respectively, confirming that NP ELL did not inhibit CISP cytotoxicity (Fig. 9a and b).
DISCUSSION
Nanotechnology is a useful tool for the amelioration of the bioavailability of natural products. This allows us to improve their effectiveness in the prevention and treatment of many diseases. Since many natural products are highly lipophilic, they are not suitable for drug delivery because they do not dissolve well in the blood.
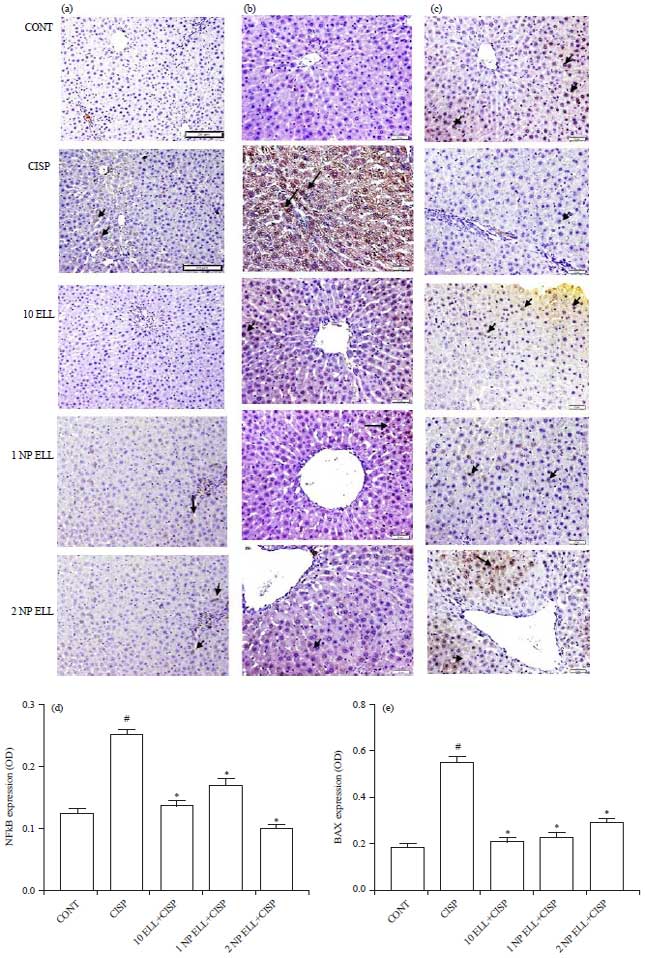 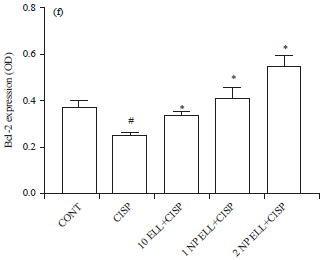 | |
| Fig. 7(a-f): | Immunohistochemistry photomicrographs of liver sections showing the effect of ellagic acid (10 ELL, 10 mg kg–1), nanoparticles ellagic acid (1 NP Ell, 1 mg kg–1) and nanoparticles ellagic acid (2 NP Ell, 2 mg kg–1) on cisplatin (CISP)-induced alteration of liver expression of NFkB (a), BAX (b) and Bcl-2 (c). The bar chart d, e and f show liver NFkB, BAX and Bcl-2 OD in the different experimental groups. Data are expressed as Mean±SEM of 6 rats/group. #p<0.05 compared with the CONT group. *p<0.05 compared with the CISP group |
| Data are compared using ANOVA followed by Tukey-Kramer post hoc test | |
Because of the lack of bioavailability of these products, administration of high doses is usually required to produce a therapeutic response. The use of these compounds in such a high dose can lead to symptoms of acute toxicity as well as the inconsistency of patients use. Only converting these lipophilic products into nanoparticles can increase their solubility and efficacy35. Although ELL has promising therapeutic potential, it is poorly soluble and has low permeability and stability at physiological pH31. In order to overcome this problem preparing ELL in nanoparticles may be a fruitful way to improve its biopharmaceutical properties and render it more clinically useful. Furthermore, nano-encapsulation of ellagic acid was shown to be effective in preventing cyclosporine-A nephrotoxicity at a three-fold lower concentration39. The focus of the current study was whether a very low dose (1 and 2 mg) of NP ELL can alleviate CISP-induced hepatotoxicity compared to a high dose (10 mg) of regular ELL.
The current study, highlighted that NP ELL is as effective or in some instances superior to ELL in protecting against CISP induced hepatotoxicity in rats. Administration ELL, 1 NP ELL and 2 NP ELL effectively restored: i) Serum levels of AST, ALT and ALP, ii) Liver levels of MDA, NO, GSH, GPx, CAT, IL-10, TNF-α and NFkB. Furthermore, immunohistochemical examination revealed that the administration of ELL, 1 NP ELL and 2 NP ELL restored the CISP-induced alteration of liver expression of NFkB, BAX and Bcl-2 to pre-treatment levels.
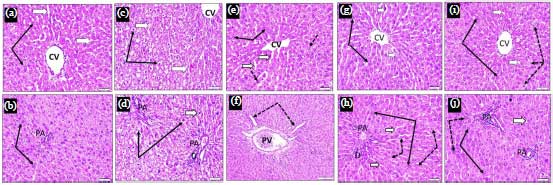 | |
| Fig. 8(a-j): | Liver cross sections (H and E stain x 400) obtained from control (CONT) (a and b), cisplatin (CISP, 7.5 mg kg–1) (c and d), ellagic acid (10 ELL, 10 mg kg–1), nanoparticles ellagic acid (1 NP Ell, 1 mg kg–1) (e and f) and nanoparticles ellagic acid (2 NP Ell, 2 mg kg–1) (g and h) showing hepatocytes at Central vein (CV) and portal area (PA) regions. a and b CONT: hepatocytes are arranged in cell plates they have acidophilic cytoplasm and large central vesicular active nuclei (thin black arrows). Blood sinusoids between cell plates are normal and lined by endothelial cells (white arrows). c and d CISP: hepatocytes are swollen compressing blood sinusoids (white arrows), their cytoplasm is unstained (hydropic degeneration) and their nuclei (thin black arrows) are small dark stained and degenerated (pyknotic). e and f 10 Ell: showing normal hepatocytes (black arrows), dilated sinusoids (white arrows). Few hepatocytes still showed dark pyknotic nuclei or vacuolated cytoplasm (dotted arrows). g and h 1 NP Ell: Showing marked protection where most hepatocytes showed intact outlines, rounded vesicular active nuclei and stained cytoplasm (thin black arrows). Few scattered cells showed ill-defined outlines, slightly vacuolated cytoplasm and small or absence of nuclei (dotted arrows). i and j 2 NP Ell: Most hepatocytes showed well defined outlines, stained cytoplasm and large vesicular active nuclei (thin black arrows) |
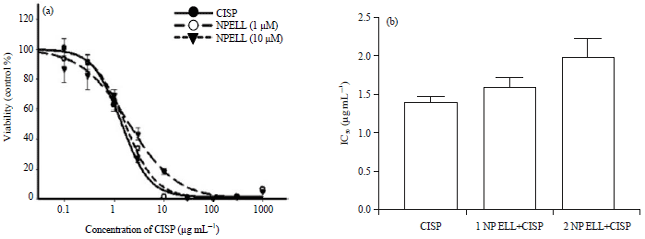 | |
| Fig. 9(a-b): | Cytotoxicity of CISP was assessed in vitro with and without NP ELL against HCT 116 colorectal cancer cells (a) CISP, CISP+NP ELL (1 μM) and CISP+NP ELL (10 μM) using SRB viability assay and (b) Calculated IC50 of CISP in presence of NP ELL (1 and 10 μM) in HCT116 cells, respectively. Data are expressed as mean±SEM of 6 rats/group. *p<0.05 compared with the CISP group |
| Data are compared using ANOVA followed by Tukey-Kramer post hoc test | |
Examination of liver sections showed that ELL, 2 NP ELL and 2 NP ELL resulted in a marked protective effect on hepatocytes in comparison to the CISP group, with 1 NP ELL and 2 NP ELL showing a higher protective effect.
The findings in this study agree with Yuce et al.10 in which ELL prevented CISP induced liver toxicity. Yuce et al.10 reported that regular formulation of ELL restored levels of MDA, GSH, GSH-Px and CAT as well as ameliorated the histopathological changes caused by CISP on liver and heart tissue in rats10. Restoration of serum levels of AST, ALT and ALP in the current study further supports the protection of ELL in its various formulations against CISP induced hepatotoxicity. Liver cells contain high levels of hepatic enzymes. Increased levels of liver enzymes in the serum are a strong sign for cell membrane function loss and even hepatic cell damage. The CISP toxicity leads to liver cell damage and leakage of AST, ALT and ALP enzymes into the blood stream40.
The CISP is a generator of reactive oxygen species and its administration results in an up regulation of free radicals, which ultimately lead to the destruction of cellular structures1,41. It has been postulated that higher levels of MDA found in the CISP treated animals as found in the present study, is due to a reduction of anti-oxidants in these tissues36. GSH is a key-player in cellular metabolism and possesses reducing properties, which are essential in preserving the integrity of cells41. The CAT and GPx have a protective role against reactive oxygen species. Therefore, the restoration of CAT and GPx is key to the hepatoprotection caused by ELL in its various formulation. This restoration of anti-oxidants was consistent with other studies, which examined the role that anti-oxidants had in relation to CISP toxicity in animals36,42. These results are concurrent with the study by Yuce et al.10. They reported an elevation of hepatic markers of oxidative stress in CISP treated rats, with a protective antioxidant effect exerted by ELL10. The ELL possesses a strong ability to remove free radicals both in vivo and in vitro43,44.
The roles of TNF-α and IL-10 have been well recognized in linking three different mechanisms, which underlie acute toxicity cases: Inflammation, oxidative stress and apoptosis45-47. CISP toxicity caused an enormous inflammatory reaction in liver cells, which is confirmed by elevated blood level of TNF-α and other anti-inflammatory mediators as well as decreased level of IL-1045,48. The TNF-α supported the liver tissue inflammation through increasing the chemotaxis of immune cells and activation of other inflammatory cytokines49. Moreover, the fall in IL-10 exacerbates liver damage because it down regulates antigen presentation and the pro-inflammatory cytokines liberation50,51. Liver levels of IL-10 and TNF-α were also restored by ELL in the present study further supporting its hepatoprotective role. The 10 ELL, 1 NP ELL and 2 NP ELL showed a marked protective effect to CISP-induced hepatotoxicity in rats. This was mainly attributed to the down regulation of TNF-α and increased levels of IL-10. Furthermore, it also acted by indirectly by inhibiting ROS liberation. These effects collectively supported the use of 10 ELL, 1 NP ELL and 2 NP ELL in treating CISP-induced liver complications.
Similar to a number of other reports, this study found that the expression of NFkB was up regulated in CIS treated animals42,52,53. In addition, the current data showed that, 10 ELL, 1 NP ELL and 2 NP ELL pre-treatment decreased NF-kB expression in the liver of CISP-treated rats54. It is well known that NFkB plays a central role in regulating inflammatory mediators and its activation results in apoptosis53,55. The NF-kB is a cytoplasmic nuclear transcription factor bound to I-kB (an inhibitory subunit). In the presence of excess reactive oxygen species, NFkB is activated and dissociated from I-kB. Consequently, NF-kB moves to the nucleus of the hepatocyte binds to DNA and enhances the transcription of plenty of the inflammatory genes like cytokine, chemokine which are involved in inflammatory and immune responses and the predisposition to fibrosis, apoptosis and acute phase responses that cause tissue and organ damage56.
The BAX and Bcl-2, which are pro and anti-apoptotic respectively were shown to play a role in CISP-induced cell death57. In the current study, BAX was increased by CIS, whereas Bcl-2 levels were decreased which is consistent with previous studies36,42. This effect was reversed in the presence of ELL.
The cytotoxicity of CISP in the presence of NP ELL was assessed using HCT116 colorectal cancer cell. The results confirmed that both 1 NP ELL and 2 NP ELL did not inhibit the cytotoxic effect CISP against HCT116 colorectal cancer cells.
CONCLUSION
Nanoformulation of ELL improves its bio-availability and protects against CISP induced hepatotoxicity. This provides an encouraging way of ameliorating the bio-availability of ELL while sustaining its desirable therapeutic effects.
SIGNIFICANCE STATEMENT
The results of this study showed for the first time that two very low doses of NP ELL are very effective in mitigating CISP-induced hepatotoxicity. The effect is comparable to that of a high dose of regular ELL. This finding will be beneficial to many patients suffering from various cancers treated with CISP. In addition, it opens the field in front of researchers to search the protective effect of the NP ELL in human beings.
ACKNOWLEDGMENT
This project was funded by the Deanship of Scientific Research (DSR) at King Abdulaziz University, Jeddah, under Grant No. G-203-249-38. The authors, therefore, acknowledge with thanks the DSR for technical and financial support.
REFERENCES
- Dasari, S. and P.B. Tchounwou, 2014. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol., 740: 364-378.
CrossRefDirect Link - Cui, W., E.M. Yazlovitskaya, M.S. Mayo, J.C. Pelling and D.L. Persons, 2000. Cisplatin-induced response of c‐jun N‐terminal kinase 1 and extracellular signal‐regulated protein kinases 1 and 2 in a series of cisplatin‐resistant ovarian carcinoma cell lines. Mol. Carcinogenesis, 29: 219-228.
CrossRefDirect Link - Barabas, K., R. Milner, D. Lurie and C. Adin, 2008. Cisplatin: A review of toxicities and therapeutic applications. Vet. Comparat. Oncol., 6: 1-18.
CrossRefDirect Link - Kuhlmann, M.K., G. Burkhardt and H. Kohler, 1997. Insights into potential cellular mechanisms of cisplatin nephrotoxicity and their clinical application. Nephrol. Dial. Transplant., 12: 2478-2480.
Direct Link - Zicca, Z., S. Cafaggi, M.A. Mariggio, M.O. Vannozzi and M. Ottone et al., 2002. Reduction of cisplatin hepatotoxicity by procainamide hydrochloride in rats. Eur. J. Pharmacol., 442: 265-272.
CrossRefPubMedDirect Link - Mora, L.de.O., L.M.G. Antunes, H.D.C. Francescato and M.de.L.P. Bianchi, 2003. The effects of oral glutamine on cisplatin-induced nephrotoxicity in rats. Pharmacol. Res., 47: 517-522.
CrossRefPubMedDirect Link - Kim, Y.K., J.S. Jung, S.H. Lee and Y.W. Kim, 1997. Effects of antioxidants and Ca2+ in cisplatin-induced cell injury in rabbit renal cortical slices. Toxicol. Applied Pharmacol., 146: 261-269.
CrossRefDirect Link - Gong, C., L. Qian, H. Yang, L.L. Ji and H. Wei et al., 2015. Hepatotoxicity and pharmacokinetics of cisplatin in combination therapy with a traditional Chinese medicine compound of Zengmian Yiliu granules in ICR mice and SKOV-3-bearing nude mice. BMC Complement. Altern. Med., Vol. 15.
CrossRefDirect Link - Pratibha, R., R. Sameer, P.V. Rataboli, D.A. Bhiwgade and C.Y. Dhume, 2006. Enzymatic studies of cisplatin induced oxidative stress in hepatic tissue of rats. Eur. J. Pharmacol., 532: 290-293.
CrossRefPubMedDirect Link - Yüce, A., A. Ateşşahin, A.O. Çeribaşı and M. Aksakal, 2007. Ellagic acid prevents cisplatin-induced oxidative stress in liver and heart tissue of rats. Basic Clin. Pharmacol. Toxicol., 101: 345-349.
CrossRefPubMedDirect Link - Giridharan, V.V., R.A. Thandavarayan, H.N. Bhilwade, K.M. Ko, K. Watanabe and T. Konishi, 2012. Schisandrin B, attenuates cisplatin-induced oxidative stress, genotoxicity and neurotoxicity through modulating NF-κB pathway in mice. Free Radic. Res., 46: 50-60.
CrossRefDirect Link - Srivastava, R.C., A. Farookh, N. Ahmad, M. Misra, S.K. Hasan and M.M. Husain, 1996. Evidence for the involvement of nitric oxide in cisplatin-induced toxicity in rats. Biometals, 9: 139-142.
CrossRefDirect Link - Omar, H.A., W.R. Mohamed, H.H. Arab and E.S.A. Arafa, 2016. Tangeretin alleviates cisplatin-induced acute hepatic injury in rats: Targeting MAPKs and apoptosis. PloS one, Vol. 11.
Direct Link - Omar, H.A., W.R. Mohamed, E.S.A. Arafa, B.A. Shehata, G.A. El Sherbiny, H.H. Arab and A.N.A. Elgendy, 2016. Hesperidin alleviates cisplatin-induced hepatotoxicity in rats without inhibiting its antitumor activity. Pharmacol. Rep., 68: 349-356.
CrossRefDirect Link - Chen, K.H., M.S. Weng and J.K. Lin, 2007. Tangeretin suppresses IL-1β-induced cyclooxygenase (COX)-2 expression through inhibition of p38 MAPK, JNK and AKT activation in human lung carcinoma cells. Biochem. Pharmacol., 73: 215-227.
CrossRefDirect Link - Sak, K., 2014. Cytotoxicity of dietary flavonoids on different human cancer types. Pharmacogn. Rev., 8: 122-146.
CrossRefDirect Link - Soong, Y.Y. and P.J. Barlow, 2004. Antioxidant activity and phenolic content of selected fruit seeds. Food Chem., 88: 411-417.
CrossRefDirect Link - Vattem, D.A. and K. Shetty, 2005. Biological functionality of ellagic acid: A review. J. Food Biochem., 29: 234-266.
CrossRefDirect Link - Baek, B., S.H. Lee, K. Kim, H.W. Lim and C.J. Lim, 2016. Ellagic acid plays a protective role against UV-B-induced oxidative stress by up-regulating antioxidant components in human dermal fibroblasts. Korean J. Physiol. Pharmacol., 20: 269-277.
CrossRefDirect Link - Festa, F., T. Aglitti, G. Duranti, R. Ricordy, P. Perticone and R. Cozzi, 2001. Strong antioxidant activity of ellagic acid in mammalian cells in vitro revealed by the comet assay. Anticancer Res., 21: 3903-3908.
Direct Link - Han, D.H., M.J. Lee and J.H. Kim, 2006. Antioxidant and apoptosis-inducing activities of ellagic acid. Anticancer Res., 26: 3601-3606.
Direct Link - Rogerio, A.P., C. Fontanari, E. Borducchi, A.C. Keller and M. Russo et al., 2008. Anti-inflammatory effects of Lafoensia pacari and ellagic acid in a murine model of asthma. Eur. J. Pharmacol., 580: 262-270.
CrossRefPubMedDirect Link - Marin, M., R.M. Giner, J.L. Rios and M.C. Recio, 2013. Intestinal anti-inflammatory activity of ellagic acid in the acute and chronic dextrane sulfate sodium models of mice colitis. J. Ethnopharmacol., 150: 925-934.
CrossRefDirect Link - Favarin, D.C., M.M. Teixeira, E.L. de Andrade, C.F. Alves and J.E.L. Chica et al., 2013. Anti-inflammatory effects of ellagic acid on acute lung injury induced by acid in mice. Mediators Inflamm.
CrossRefDirect Link - Priyadarsini, K.I., S.M. Khopde, S.S. Kumar and H. Mohan, 2002. Free radical studies of ellagic acid, a natural phenolic antioxidant. J. Agric. Food Chem., 50: 2200-2206.
CrossRefDirect Link - Urfalioglu, A., F.M. Yazar, B. Bilal, F.I. Tolun, H. Oksuz, O.F. Boran and G. Oksuz, 2017. The effect of p-coumaric acid and ellagic acid on the liver and lungs in a rat model of sepsis. Asian Biomed., 11: 217-225.
Direct Link - Dubey, A., D.W. Park, J.E. Kwon, Y.J. Jeong and T. Kim et al., 2015. Investigation of the biological and anti-cancer properties of ellagic acid-encapsulated nano-sized metalla-cages. Int. J. Nanomed., 10: 227-240.
Direct Link - Mady, F.M. and M.A. Shaker, 2017. Enhanced anticancer activity and oral bioavailability of ellagic acid through encapsulation in biodegradable polymeric nanoparticles. Int. J. Nanomed., 12: 7405-7417.
Direct Link - Zhang, H.M., L. Zhao, H. Li, H. Xu, W.W. Chen and L. Tao, 2014. Research progress on the anticarcinogenic actions and mechanisms of ellagic acid. Cancer Biol. Med., 11: 92-100.
Direct Link - Ahire, V., A. Kumar, K.P. Mishra and G. Kulkarni, 2017. Ellagic acid enhances apoptotic sensitivity of breast cancer cells to γ-radiation. Nutr. Cancer, 69: 904-910.
CrossRefDirect Link - Bala, I., V. Bhardwaj, S. Hariharan and M.R. Kumar, 2006. Analytical methods for assay of ellagic acid and its solubility studies. J. Pharm. Biomed. Anal., 40: 206-210.
CrossRefDirect Link - Lei, F., D.M. Xing, L. Xiang, Y.N. Zhao, W. Wang, L.J. Zhang and L.J. Du, 2003. Pharmacokinetic study of ellagic acid in rat after oral administration of pomegranate leaf extract. J. Chromatogr. B, 796: 189-194.
CrossRefDirect Link - Seeram, N.P., R. Lee and D. Heber, 2004. Bioavailability of ellagic acid in human plasma after consumption of ellagitannins from pomegranate (Punica granatum L.) juice. Clin. Chim. Acta, 348: 63-68.
CrossRefDirect Link - Bala, I., V. Bhardwaj, S. Hariharan, S.V. Kharade, N. Roy and M.N.V.R. Kumar, 2006. Sustained release nanoparticulate formulation containing antioxidant-ellagic acid as potential prophylaxis system for oral administration. J. Drug Target., 14: 27-34.
CrossRefDirect Link - Watkins, R., L. Wu, C. Zhang, R.M. Davis and B. Xu, 2015. Natural product-based nanomedicine: Recent advances and issues. Int. J. Nanomed., 10: 6055-6074.
Direct Link - El‐Shitany, N.A. and B. Eid, 2017. Proanthocyanidin protects against cisplatin‐induced oxidative liver damage through inhibition of inflammation and NF‐κβ/TLR‐4 pathway. Environ. Toxicol., 32: 1952-1963.
CrossRefDirect Link - Skehan, P., R. Storeng, D. Scudiero, A. Monks and J. McMahon et al., 1990. New colorimetric cytotoxicity assay for anticancer-drug screening. J. Nat. Cancer Inst., 82: 1107-1112.
CrossRefPubMedDirect Link - Mahmoud, A.M., A.M. Al-Abd, D.A. Lightfoot and H.A. El-Shemy, 2012. Anti-cancer characteristics of mevinolin against three different solid tumor cell lines was not solely p53-dependent. J. Enzyme Inhib. Med. Chem., 27: 673-679.
CrossRefPubMedDirect Link - Sonaje, K., J.L. Italia, G. Sharma, V. Bhardwaj, K. Tikoo and M.R. Kumar, 2007. Development of biodegradable nanoparticles for oral delivery of ellagic acid and evaluation of their antioxidant efficacy against cyclosporine A-induced nephrotoxicity in rats. Pharm. Res., 24: 899-908.
CrossRefDirect Link - Yilmaz, H.R., M. Iraz, S. Sogut, H. Ozyurt, Z. Yildirim, O. Akyol and S. Gergerlioglu, 2004. The effects of erdosteine on the activities of some metabolic enzymes during cisplatin-induced nephrotoxicity in rats. Pharmacol. Res., 50: 287-290.
PubMed - Conklin, K.A., 2000. Dietary antioxidants during cancer chemotherapy: Impact on chemotherapeutic effectiveness and development of side effects. Nutr. Cancer, 37: 1-18.
PubMedDirect Link - Neamatallah, T., N.A. El-Shitany, A.T. Abbas, S.S. Ali and B.G. Eid, 2018. Honey protects against cisplatin-induced hepatic and renal toxicity through inhibition of NF-κB-mediated COX-2 expression and the oxidative stress dependent BAX/Bcl-2/caspase-3 apoptotic pathway. Food Funct., 9: 3743-3754.
Direct Link - Omur, A.D. and K. Coyan, 2016. Protective effects of the antioxidants curcumin, ellagic acid and methionine on motility, mitochondrial transmembrane potential, plasma membrane and acrosome integrity in freeze-thawed Merino ram sperm. Vet. Med., 61: 10-16.
CrossRefDirect Link - Yousef, A.I., O.S. El-Masry and M.A. Abdel Mohsen, 2016. Impact of cellular genetic make-up on colorectal cancer cell lines response to ellagic acid: Implications of small interfering RNA. Asian Pac. J. Cancer Prev., 17: 743-748.
Direct Link - Tadagavadi, R.K. and W.B. Reeves, 2010. Endogenous IL-10 attenuates cisplatin nephrotoxicity: Role of dendritic cells. J. Immunol., 185: 4904-4911.
CrossRefDirect Link - Chou, W.Y., C.N. Lu, T.H. Lee, C.L. Wu and K.S. Hung et al., 2006. Electroporative interleukin‐10 gene transfer ameliorates carbon tetrachloride‐induced murine liver fibrosis by MMP and TIMP modulation 1. Acta Pharmacol. Sinica, 27: 469-476.
CrossRefDirect Link - Shaw, J., B. Chen, W.H. Huang, A.R. Lee and F.A. Valeriote, 2011. The small-molecule TNF-α modulator, UTL-5g, reduces side effects induced by cisplatin and enhances the therapeutic effect of cisplatin in vivo. J. Exp. Ther. Oncol., 9: 129-137.
Direct Link - Rehman, M.U., N. Ali, S. Rashid, T. Jain and S. Nafees et al., 2014. Alleviation of hepatic injury by chrysin in cisplatin administered rats: Probable role of oxidative and inflammatory markers. Pharmacol. Rep., 66: 1050-1059.
CrossRefDirect Link - Lawrence, T., 2009. The nuclear factor NF-κB pathway in inflammation. Cold Spring Harb Perspect Biol., Vol. 1.
Direct Link - Koppelman, B., J.J. Neefjes, J.E. de Vries and R. de Waal Malefyt, 1997. Interleukin-10 down-regulates MHC class II αβ peptide complexes at the plasma membrane of monocytes by affecting arrival and recycling. Immunity, 7: 861-871.
CrossRefDirect Link - Knolle, P.A., A. Uhrig, S. Hegenbarth, E. Loser, E. Schmitt, G. Gerken and A.W. Lohse, 1998. IL-10 down-regulates T cell activation by antigen-presenting liver sinusoidal endothelial cells through decreased antigen uptake via the mannose receptor and lowered surface expression of accessory molecules. Clin. Exp. Immunol., 114: 427-433.
Direct Link - Kang, K.P., D.H. Kim, Y.J. Jung, A.S. Lee and S. Lee et al., 2009. Alpha-lipoic acid attenuates cisplatin-induced acute kidney injury in mice by suppressing renal inflammation. Nephrol. Dial. Transplant., 24: 3012-3020.
CrossRefDirect Link - Ma, Z.N., Y.Z. Li, W. Li, X.T. Yan and G. Yang et al., 2017. Nephroprotective effects of saponins from leaves of Panax quinquefolius against cisplatin-induced acute kidney injury. Int. J. Mol. Sci., Vol. 18.
CrossRefDirect Link - Gu, L., W.S. Deng, Y. Liu, C.H. Jiang and L.C. Sun et al., 2014. Ellagic acid protects Lipopolysaccharide/D-galactosamine-induced acute hepatic injury in mice. Int. Immunopharmacol., 22: 341-345.
CrossRefDirect Link - Morgan, M.J. and Z.G. Liu, 2010. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res., 21: 103-115.
CrossRefDirect Link - Hritcu, L., E. Bagci, E. Aydin and M. Mihasan, 2015. Antiamnesic and antioxidants effects of Ferulago angulata essential oil against scopolamine-induced memory impairment in laboratory rats. Neurochem. Res., 40: 1799-1809.
CrossRefDirect Link - Jiang, M., C.Y. Wang, S. Huang, T. Yang and Z. Dong, 2009. Cisplatin-induced apoptosis in p53-deficient renal cells via the intrinsic mitochondrial pathway. Am. J. Physiol.-Renal Physiol., 296: F983-F993.
CrossRefDirect Link