
Huan He
Department of Physiology, Medical School, Nanchang University, Bayi Road 461, 330006 Nanchang, Jiangxi Province, People`s Republic of China
Hang Yan
Department of Physiology, Medical School, Nanchang University, Bayi Road 461, 330006 Nanchang, Jiangxi Province, People`s Republic of China
Yan Zhao
Department of Physiology, Medical School, Nanchang University, Bayi Road 461, 330006 Nanchang, Jiangxi Province, People`s Republic of China
Yanrong Yu
Jiangxi Academy of Medical Sciences, Bayi Road 461, 330006 Nanchang, Jiangxi Province, People`s Republic of China
Haibing Kuang
Department of Physiology, Medical School, Nanchang University, Bayi Road 461, 330006 Nanchang, Jiangxi Province, People`s Republic of China
Qiren Huang
Department of Pharmacology, Fuzhou Medical College, Nanchang University, Dongling Road 9, 344000 Fuzhou, Jiangxi Province, People`s Republic of China
Ming He
Department of Pharmacology, Fuzhou Medical College, Nanchang University, Dongling Road 9, 344000 Fuzhou, Jiangxi Province, People`s Republic of China
Dan Luo
Department of Physiology, Medical School, Nanchang University, Bayi Road 461, 330006 Nanchang, Jiangxi Province, People`s Republic of China
LiveDNA: 86.20650
Weijie Peng
Key Provincial Laboratory of Basic Pharmacology, Nanchang University, Bayi Road 461, 330006 Nanchang, Jiangxi Province, People`s Republic of China
International Journal of Pharmacology
Year: 2018 | Volume: 14 | Issue: 5 | Page No.: 698-706







ABSTRACT
Background and Objective: Gap junction (GJ) dysfunction induced by altered expression of connexin are associated with the pathogenesis of diabetes cardiovascular complications. The authors explored the effects of rutaecarpine, an active ingredient of Chinese herbal medicine Evodia, on GJ dysfunction induced by high glucose. Materials and Methods: Human umbilical vein endothelial cells (HUVECs) were treated with different concentration of high glucose (5.5, 22, 33 and 44 mM) for 48 h and mannitol group (5.5 mM glucose plus 27.5 mM mannitol) as osmotic control. To test the effects of rutaecarpine, cells were pretreated with 0.1, 0.3 and 1 μM rutaecarpine for 10 min before high glucose (33 mM) treatment. The protein level of connexins were detected by western blot. The GJ function of endothelial cells was measured by scrape-loading dye transfer experiments. Endothelial cell function was evaluated by cell viability (MTT), NO production and ROS level (DCFH-DA). Results: Western blot indicated that high glucose 33 or 44 mM reduced the level of Cx37, but had no impact on expression of Cx40 and Cx43. Scrape-loading dye transfer showed that high glucose impaired PI Cx37-permeant dye diffusion between HUVECs. Pretreatment with rutaecarpine (0.1, 0.3 and 1 μM) recovered the level of Cx37, thereby improving PI-mediated GJ communication and prevented endothelial dysfunction, as indicated by increased cell viability and NO production, as well as decreased ROS level. Additionally, flow cytometry showed that rutaecarpine induced the influx of Ca2+. These effects of rutaecarpine were attenuated by capsazepine, an antagonist of transient receptor potential vanilloid subtype 1 (TRPV1). Conclusion: These data indicate that rutaecarpine prevented high glucose-induced GJ dysfunction by recovering Cx37 level via activation of TRPV1.
PDF Abstract XML References Citation
Received: October 16, 2017;
Accepted: January 01, 2018;
Published: June 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Huan He, Hang Yan, Yan Zhao, Yanrong Yu, Haibing Kuang, Qiren Huang, Ming He, Dan Luo and Weijie Peng, 2018. Rutaecarpine Prevents High Glucose-induced Cx37 Gap Junction Dysfunction in Human Umbilical Vein Endothelial Cells. International Journal of Pharmacology, 14: 698-706.
DOI: 10.3923/ijp.2018.698.706
URL: https://scialert.net/abstract/?doi=ijp.2018.698.706
DOI: 10.3923/ijp.2018.698.706
URL: https://scialert.net/abstract/?doi=ijp.2018.698.706
INTRODUCTION
Cardiovascular complications are the main cause of morbidity and mortality in diabetes. Hyperglycemia appears to be an important etiologic factor in the development of micro- and macrovascular complications in diabetic patients1. However, the cellular mechanisms involved remain incompletely understood.
Emerging evidence suggests that gap junctions intercellular communication (GJIC) plays an important role in cardiovascular tissue homeostasis2. Gap junctions are intercellular membrane channels that allow the exchange of metabolites, ions and second messengers between adjacent cells. GJs are composed of connexins (Cxs), a large family of transmembrane proteins. To date, approximately 20 mammalian Cx isotypes have been identified. Vascular endothelial GJIC is mediated by the Cx proteins Cx37, Cx40 and Cx43, which are involved in maintenance of endothelial function3. Endothelial dysfunction is considered a major risk factor of cardiovascular complications of Type 1 and Type 2 diabetes. Previous studies had shown that high glucose level or a diabetic state alters the expression of connexin or GJ function in some types of vascular endothelial cells, which may contribute to endothelial cell dysfunction in diabetes1. High glucose inhibited GJIC activity by reducing Cx43 expression in rat microvascular endothelial4 and bovine retinal endothelial cells5, which may be related to diabetic retinopathy. Additionally, coronary ECs isolated from diabetic mice exhibited lowered protein levels of Cx37 and Cx40 (but not Cx43) and a loss of GJIC, which contribute to coronary vascular dysfunction in diabetes6. However, little is known about the effect of high glucose on connexin expression and GJIC function in macrovascular endothelial cells. In this study, the authors investigated the effects of different high glucose concentrations on connexin level and GJIC activity in cultured human umbilical vein endothelial cells (HUVECs).
Rutaecarpine is the major quinazolinocarboline alkaloidal compound of Evodia rutaecarpa ('Wu-Chu-Yu'), a well-known Chinese herbal drug. Rutaecarpine exerts multiple cardiovascular biological effects, such as positive inotropic and chronotropic, vasodilation and hypotensive effect, which were proved to be related to activation of the transient receptor potential vanilloid subtype 1 (TRPV1)7. TRPV1 is mainly distributed in the terminals and cell bodies of sensory neurons and also wildly presented in nonneurons tissues, such as vascular endothelial cell8. The authors have recently found that rutaecarpine attenuates Ox-LDL-induced endothelial dysfunction in cultured HUVECs. These protective effects are related to the regulation of Cx37, Cx40 and Cx43 expression and improvement of GJ function in endothelial cells. These effects can be blocked by capsazepine, a competitive TRPV1 antagonist, suggesting the key role of TRPV1 in the protective effects of rutaecarpine9. Thus, the present study was to evaluate the effects of rutaecarpine on the endothelial injury and GJ dysfunction induced by high glucose and investigate whether the effects of rutaecarpine are related to activation of TRPV1.
MATERIALS AND METHODS
Cell culture and treatment: HUVEC-12, an immortalized human umbilical vein cell line, was obtained from the American Type Culture Collection and cultured in DMEM containing 10% FBS. The cells were maintained in a humidified incubator at 37°C in a 5% CO2 atmosphere until reaching sub-confluence.
The first series experiments were designed to test the concentration-dependent effects of high glucose on the connexins expression and GJIC function in HUVECs. Endothelial cells were exposed to different dose of glucose (5.5, 22, 33 and 44 mM) for 48 h respectively. To exclude the effect of hyperosmolarity induced by high glucose, mannitol group (5.5 mM glucose plus 27.5 mM mannitol) was used as osmotic control at 33 mM.
The second series experiments were designed to test the effects of rutaecarpine on HUVECs exposed to high glucose. Cells were divided into 6 groups: (1) Normal control(5.5 mM glucose), (2) High glucose groups, where cell damage was induced by incubation with 33 mM glucose for 24 or 48 h; the (3, 4 and 5) rutaecarpine groups (+Rut), where cells were pretreated with 0.1, 0.3 and 1 μM rutaecarpine (Juntuo Biotechnology Co., Ltd., Shanghai, China), respectively, for 10 min before high glucose treatment and (6) the capsazepine plus rutaecarpine group (+CAPZ+Rut), where ECs were pretreated with 10 μM capsazepine (Sigma Aldrich, St. Louis, MO) for 10 min before rutaecarpine (1 μM) treatment and the subsequent incubation with high glucose for 24 or 48 h.
Western blot: The cells were lysed at 4°C and the supernatant was collected after centrifugation. The protein concentration was measured using a BCA protein assay kit (Generay Biotech Co., Ltd., Shanghai, China) and equal amounts of protein were then separated by 12% sodium dodecyl sulphate polyacrylamide gel electrophoresis and transferred onto a polyvinylidene fluoride membrane. After blocking for 2 h with 5% non-fat milk, the membrane was incubated with diluted primary antibodies (Abcam, Cambridge, UK) against Cx37, Cx40, Cx43 and β-actin at 4°C overnight. After five washes, the membrane was incubated with a 1:10,000 dilution of horseradish peroxidase-labelled goat anti-rabbit or goat anti-mouse antibody (Zhongshan Gold Bridge Bio-Tech Inc., Beijing, China) at room temperature for 1 h. The membrane was developed using enhanced chemiluminescence after five washes. All experiments were performed with three replicates each.
Scrape-loading dye transfer experiments: The GJIC in HUVEC-12 was evaluated using scrape-loading dye transfer experiments, as previously described9. HUVECs express Cx37, Cx40 and Cx43; thus, the authors investigated the diffusion of two dyes: LY, which readily passes through GJs formed by Cx40 and Cx43 but not through channels formed by Cx37 and PI, which can pass through Cx37-mediated GJs. Highly-confluent HUVEC-12 monolayers were rinsed with PBS and incubated in PBS containing 0.5% lucifer yellow (LY) and 5 g L–1 propidium iodide (PI; Sigma Aldrich, St. Louis, MO, USA). Cells were scraped by applying gentle pressure with a scalpel blade before they were incubated at 37°C in the dark for 5 min. The dye mixture was removed and the cells were washed thrice in PBS before they were immediately fixed in 4% paraformaldehyde. The cells were recorded and analysed using the Wildfield High-Content Screening System (Molecular Devices, USA). Cells stained with LY or PI from the scrape-loaded cells were considered to communicate via GJs. Thus, GJIC was determined by counting the number of cells stained by LY or PI and expressed as percentage of the control.
Flow cytometric measurement of intracellular calcium concentration: Cells cultured in 12-well plates were loaded with 5 μM Fluo-3-AM (Ca2+- sensitive dye) in the dark for 30 min at 37°C and then were rinsed 3 times with PBS to remove free dye. After trypsin digestion, cells were collected and resuspended in PBS. Fluo-3-loaded cells were then were stimulated with different doses of Rut for 5 min. Fluo-3-AM fluorescence was measured on the flow cytometer with emission wavelengths of 525 nm and excitation wavelengths of 488 nm.
Cell viability assay: Cell viability was determined by the tetrazolium salt MTT (3-[4,5-Dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide) assay (Sigma Aldrich, St. Louis, MO, USA), according to standard techniques. The optical density of each sample was immediately measured using an ELISA microplate reader (ELx800; USA) at 490 nm.
Nitric oxide (NO) concentration: The NO levels in the conditioned medium were measured using an NO assay kit (Jiancheng Bioengineering Institute, Nanjing, China). In brief, the nitrates in the medium were converted to nitrite and then measured using the Griess reagent. The absorbance of each sample was determined at 540 nm using a spectrophotometer.
Reactive oxygen species (ROS) assay: The level of cytosolic ROS was measured by use of DCFH-DA (Beyotime, China). Briefly, HUVECs cells were washed with PBS and incubated with DCFH-DA for 30 min at 37°C with gentle shaking. DCFH-DA was initially nonfluorescent and was converted by oxidation to the fluorescent molecule DCFH. DCFH fluorescence was then quantified by use of a SpectraMax Paradigm Multi-Mode Microplate Reader Fluoroskan Ascent, Molecular Devices,USA at kex 488 nm/kem 525 nm.
Statistical analysis: Data were expressed as Mean±SEM. All values were analyzed by using one-way ANOVA and multiple comparison test (the Student Newman Keuls t-test) by using SPSS version 13.0. The acceptable value of significance was p<0.05.
RESULTS AND DISCUSSION
High glucose reduces Cx37 level and inhibits GJIC function in HUVECs: The concentration-dependent effects of high glucose on the expression of connexins in HUVECs were assessed via Western blot analysis. Cultured HUVECS expressed Cx37, Cx40 and Cx43. Exposure to 22 mM glucose for 24 h failed to affect the level of connexins. The protein level Cx37 was remarkably down regulated when the concentration of glucose reached 33 and 44 mM. Whereas Cx40 and Cx43 remained unchanged. Mannitol (as osmotic control of 33 mM) had no effect on the level of connexin, eliminating the influence of medium hyperosmolarity Fig. 1a and b.
Scrape-loading dye transfer experiments on HUVECs were performed to determine whether high glucose affected GJIC function in HUVECs. HUVECs express Cx37, Cx40 and Cx43; thus, the authors investigated the diffusion of two dyes: LY, which readily passes through GJs formed by Cx40 and Cx43 but not through channels formed by Cx37 and PI, which can pass through Cx37-mediated GJs. Scrape-loading of cultured HUVECs enabled a prominent diffusion of PI, whereas LY diffusion was mostly limited to the injured cells as shown in Fig. 1c.
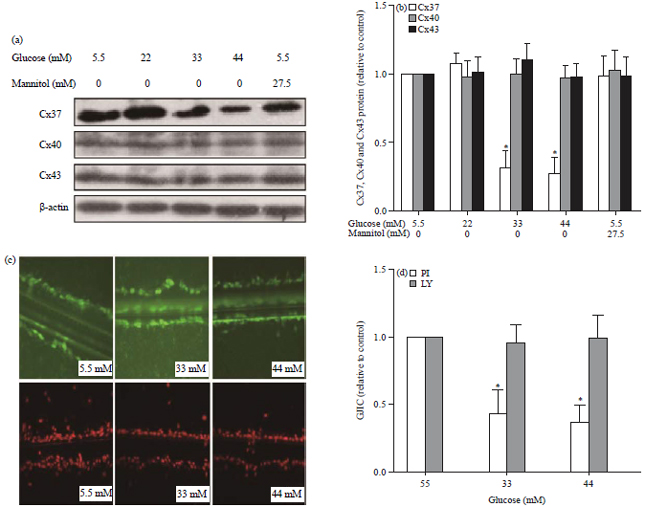 | |
| Fig. 1(a-d): | (a-b) High glucose reduced Cx37 level in HUVECs. Western blot showing the protein levels of Cx37, Cx40 and Cx43 in HUVECs exposed to different concentration of glucose (5.5, 22, 33 and 44 mM) and (c-d) High glucose inhibited GJIC function in HUVECs. Scrape-loading dye transfer experiments showing the effects of high glucose (5.5, 33 and 44 mM) on GJIC in HUVECs. GJIC was determined by the diffusion of two dyes, namely, lucifer yellow (LY, green) and propidium iodide (PI, red). (Image magnification of 200 times). Values are expressed as the Mean±SEM, n = 3. *p<0.01 vs. normal glucose 5.5 mM) |
Exposing HUVECs to high glucose (33 and 44 mM) significantly inhibited the intercellular transfer of PI (the number of stained cells versus controls: 36.3±5.3%), indicating that Cx37-mediated GJ communication is severely impaired by high glucose, whereas LY diffusion remained unchanged.
Rutaecarpine recovers Cx37 GJIC function via activation of TRPV1: Western blots showed that the reduction effect of high glucose on Cx37 was reversed by rutaecarpine, which recovered the level of Cx37 protein in a dose-dependent manner. Pretreatment with capsazepine, a competitive antagonist of TRPV1, abolished the effect of rutaecarpine (Fig. 2a and b).
As shown in the scrape-loading dye transfer experiments, exposing HUVECs to high glucose significantly inhibited the intercellular transfer of PI. Pretreatment with rutaecarpine significantly improved PI diffusion (which recovered to 116.2±3.8% versus control). By contrast, pretreatment with the TRPV1 antagonist capsazepine abolished the effects of rutaecarpine (Fig. 2c and d). These results indicate that rutaecarpine can prevent the GJIC dysfunction induced by high glucose in HUVECs via TRPV1 activation.
Additionally, flow cytometry demonstrated that rutaecarpine induced Ca2+ influx in a concentration-dependent manner, which was abolished by capsazepine (Fig. 2e).
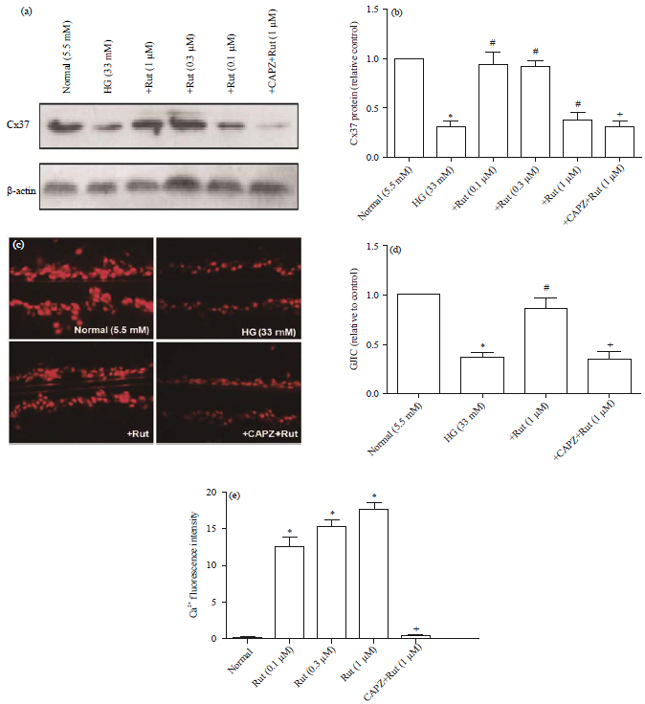 | |
| Fig. 2(a-e): | Rutaecarpine recovered the level of Cx37 and GJ function via TRPV1, (a-b) Western blot showing the effects of rutaecarpine on Cx37 protein in HUVECs exposed to high glucose, (c-d) Scrape-loading dye transfer experiments showing the effects of rutaecarpine on GJIC in HUVECs exposed to high glucose and (e) Flow cytometry showing rutaecarpine induced the Ca2+ influx via activation of TRPV1. Normal: Exposure to normal glucose (5.5 mM), High glucose: Exposure to high glucose (33 mM), +Rut: Pretreated with 0.1, 0.3 and 1 μM rutaecarpin (Rut) for 10 min, respectively, before high glucose treatment, +CAPZ+Rut (1 μM): Pretreated with 10 μM capsazepine (CAPZ) for 10 min before rutaecarpin treatment, then incubated with high glucose. Values are expressed as the Mean±SEM, n = 3. *p<0.01 vs. control, #p<0.01 vs. high glucose, +p<0.01 vs. +Rut (1 μM) |
Rutaecarpine prevents endothelial cell dysfunction induced by high glucose: Exposing HUVEC-12 to high glucose (33 mM) for 24 h led to remarkable endothelial cell damage as shown in Fig. 3, as indicated by the decreased cell viability and NO production as well as the increased ROS release, all of which were significantly attenuated by rutaecarpine (0.1, 0.3 and 1 μM) in a concentration-dependent manner.
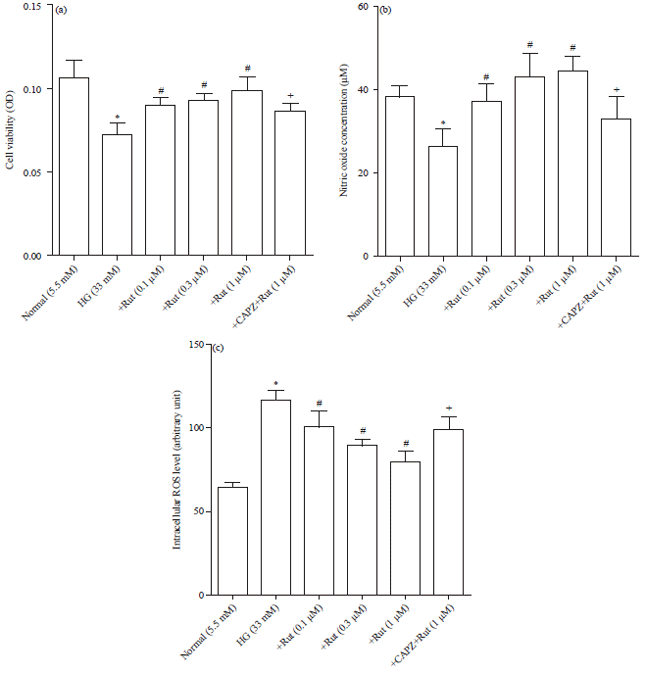 | |
| Fig. 3(a-c): | Rutaecarpine prevented the high glucose-induced endothelial cell dysfunction. Endothelial cell damage was assessed by cell viability, which was determined by (a) MTT metabolite, (b) NO level in the culture medium and (c) Intracellular ROS level. Values are expressed as the Mean±SEM, n = 6. *p<0.01 vs. normal, #p<0.01 vs. high glucose, +p<0.01 vs. +Rut (1 μM) |
These protective effects of rutaecarpine were attenuated by capsazepine.
High glucose inhibits Cx37 gap junction in HUVECs: Endothelial dysfunction is the initiating event of the micro- and macroangipathy associated with diabetes. GJIC play an important role in maintenance of endothelial function10. Vascular endothelial cells express three types of connexin, Cx37, Cx40 and Cx432. In current study, exposure HUVECs to high glucose 33 mM for 24 h significantly reduced the protein level of Cx37, whereas Cx40 and Cx43 remained unchanged even when the concentration of glucose reached 44 mM. These events were concomitant with the impaired propidium iodide PI diffusion between ECs, whereas LY diffusion remained limited. PI is a Cx37-permeant molecule, which readily passes through the Cx37-mediated GJ, whereas LY diffusion represents Cx40- or Cx43-mediated GJ11. This finding is partially consistent with the report of Hou et al.12, which found that STZ-induced diabetes (on an ApoE deficient background) down-regulated endothelial Cx37 and Cx40 outside the plaque areas, which was associated with an increase in the burden of atheroma. Alterations of the expression patterns of microvascular endothelial Cxs also been reproted during the high glucose level or diabetic state and exhibited site and tissue-specific property. Li and Roy4 reported that high glucose 30 mM reduced Cx43 expression in the rat microvascular endothelial cells4. On the contrary, a diabetic state down-regulated the protein levels of Cx37 and Cx40, but not Cx43 in coronary ECs isolated from diabetic mice6. The present study showed, for the first time, high glucose also altered the expression pattern of Cxs in human macrovascular endothelial cells.
Cx37 was mainly expressed in arteries of a healthy individual. It appears that endothelial Cx37 are important in the maintenance of endothelial quiescence, by favoring a proper NO release13 and by inhibiting leukocyte adhesion14. Multiple pathological factors, such as atherosclerosis14, tumor necrosis factor (TNF)-α15 and disturbed blood flow16 have been shown to reduce the expression of Cx37,which contribute to the endothelial dysfunction. Additionally, connexin37 polymorphism has proved to be associated with the risk of peripheral arterial disease in subjects with type 2 diabetes17. Therefore, it was speculated that impaired Cx37 GJ contribute partially to the endothelial dysfunction induced by high glucose.
Rutaecarpine improves Cx37 GJIC in HUVECs and prevents high glucose-induced endothelial dysfunction: Rutaecarpine is the major active ingredients of Evodia, a traditional Chinese herbal medicine, which has been proved to exerts multiple cardiovascular protective effects7. Researchers have recently reported that rutaecarpine improves GJIC function of HUVECs exposed to Ox-LDL by increasing the levels of Cx37 and Cx40 but inhibiting Cx43 expression, which contributed to the endothelial protection of rutaecarpine9. In the present study, the effects of rutaecarpine on endothelial dysfunction induced by high glucose were observed. As shown in the results, high glucose (33 mM) for 24 h could induce remarkable endothelial cell dysfunction, which was observed as decreased cell viability and NO production, as well as increased ROS level. Pretreatment with rutaecarpine recovered the Cx37 protein level and improved GJ communication by PI in HUVEC treated by high glucose and alleviated the high glucose-induced damage in a concentration-dependent manner, including increase the endothelial cells viability and NO release and inhibited ROS production. As mentioned above, Cx37-mediated GJs play an important role in the maintenance of endothelial function. Hence, these results suggest that rutaecarpine prevents the GJ dysfunction of endothelial cells induced by high glucose, which may contribute to, at least in part, endothelial protective effects of rutaecarpine.
TRPV1 mediates the protective effects of rutaecarpine on GJIC and endothelial function: The ligand-gated nonselective cation channel (TRPV1), is highly expressed in sensory neurons but has also been identified in blood vessels. Authors previous study confirmed the expression of TRPV1 in HUVECs8. Increasing evidence shows that rutaecarpine can activate TRPV1, which mediates multiple pharmacological actions of rutaecarpine, such as it’s inotropic and chronotropic18, vasodilation19 and cardioprotective effects20. Most recently, using TRPV1-expressing HEK293 cells and patch-clamp recording, Wang et al.21 verified that rutaecarpine acted as partial agonist of TRPV1. In the present study, we observed that the beneficial effects of rutaecarpine on endothelial function and GJ communication were attenuated by pretreatment with capsazepine, a competitive antagonist of TRPV1. Therefore, TRPV1 mediates, at least in part, the protective effect of rutaecarpine on ECs.
The mechanisms implicated in the regulatory effect of TRPV1 on Cx remain unexplored. TRPV1 activation causes a transient Ca2+ influx([Ca2+]i)22 and the latter has been shown to affect GJ communication. Ca2+ may exert an indirect effect on GJ by activating calmodulin (CaM), which can binds to different Cx types and affect GJ function23. Previous study has reported that rutaecarpine promoted inward Ca2+ currents in cultured ECs isolated from rat thoracic aorta24. Authors present study confirmed that rutaecarpine moderately increased [Ca2+]i in HUVECs, which was completely abolished by capsazepine, indicating rutaecarpine can promote Ca2+ influx through activation of TRPV1. However, whether the TRPV1/ [Ca2+]i signal pathway is involved in the regulatory effect of rutaecarpine on GJ still warrants further investigation.
CONCLUSION
The present study shows high glucose reduced Cx37 protein level, leading to GJIC dysfunction in HUVECs. Rutaecarpine prevented high glucose-induced endothelial dysfunction by recovering Cx37 GJIC function via activation of TRPV1.
SIGNIFICANCE STATEMENT
These results discovers the protective effect of rutaecarpine against high glucose-induced GJ dysfunction in endothelial cells that can be beneficial for endothelial function. This study provided a novel mechanism by which rutaecarpine may contribute to prevent macrovascular complications of diabetes and will help the researchers to uncover the role of TRPV1 activation on endothelial dysfunction in hyperglycaemia that many researchers were not able to explore.
ACKNOWLEDGMENT
This study is supported by grants from National Natural Science Foundation of China (No. 81360493 and 30801399) and the Natural Science Foundation of Jiangxi province (No. S2016QNJJB0420).
REFERENCES
- Wright, J.A., T. Richards and D.L. Becker, 2012. Connexins and diabetes. Cardiol. Res. Pract.
CrossRefDirect Link - Kar, R., N. Batra, M.A. Riquelme and J.X. Jiang, 2012. Biological role of connexin intercellular channels and hemichannels. Arch. Biochem. Biophys., 524: 2-15.
CrossRefDirect Link - Brisset, A.C., B.E. Isakson and B.R. Kwak, 2009. Connexins in vascular physiology and pathology. Antioxidants Redox Signal., 11: 267-282.
CrossRefDirect Link - Li, A.F. and S. Roy, 2009. High glucose-induced downregulation of connexin 43 expression promotes apoptosis in microvascular endothelial cells. Investi. Ophthalmol. Visual Sci., 50: 1400-1407.
CrossRefDirect Link - Bobbie, M.W., S. Roy, K. Trudeau, S.J. Munger, A.M. Simon and S. Roy, 2010. Reduced connexin 43 expression and its effect on the development of vascular lesions in retinas of diabetic mice. Investi. Ophthalmol. Visual Sci., 51: 3758-3763.
Direct Link - Makino, A., O. Platoshyn, J. Suarez, J.X.J. Yuan and W.H. Dillmann, 2008. Downregulation of connexin40 is associated with coronary endothelial cell dysfunction in streptozotocin-induced diabetic mice. Am. J. Physiol.-Cell Physiol., 295: C221-C230.
Direct Link - Jia, S. and C. Hu, 2010. Pharmacological effects of rutaecarpine as a cardiovascular protective agent. Molecules, 15: 1873-1881.
CrossRefDirect Link - Luo, D., Y.W. Zhang, W.J. Peng, J. Peng and Q.Q. Chen et al., 2008. Transient receptor potential vanilloid 1-mediated expression and secretion of endothelial cell-derived calcitonin gene-related peptide. Regul. Pept., 150: 66-72.
CrossRefDirect Link - Peng, W.J., Y. Liu, Y.R. Yu, Y.Q. Fu and Y. Zhao, 2015. Rutaecarpine prevented dysfunction of endothelial gap junction induced by Ox-LDL via activation of TRPV1. Eur. J. Pharmacol., 756: 8-14.
CrossRefPubMedDirect Link - Figueroa, X.F. and B.R. Duling, 2009. Gap junctions in the control of vascular function. Antioxidants Redox Signal., 11: 251-266.
Direct Link - Pfenniger, A., C. Wong, E. Sutter, S. Cuhlmann and S. Dunoyer-Geindre et al., 2012. Shear stress modulates the expression of the atheroprotective protein Cx37 in endothelial cells. J. Mol. Cell. Cardiol., 53: 299-309.
CrossRefDirect Link - Hou, C.J.Y., C.H. Tsai, C.H. Su, Y.J. Wu and S.J. Chen et al., 2008. Diabetes reduces aortic endothelial gap junctions in ApoE-deficient mice: Simvastatin exacerbates the reduction. J. Histochem. Cytochem., 56: 745-752.
CrossRefDirect Link - Meens, M.J., F. Alonso, L. Le Gal, B.R. Kwak and J.A. Haefliger, 2015. Endothelial Connexin37 and Connexin40 participate in basal but not agonist-induced NO release. Cell Commun. Signal., Vol. 13.
CrossRefDirect Link - Wong, C.W., T. Christen, I. Roth, C.E. Chadjichristos and J.P. Derouette et al., 2006. Connexin37 protects against atherosclerosis by regulating monocyte adhesion. Nature Med., 12: 950-954.
Direct Link - Van Rijen, H.V., M.J. Van Kempen, S. Postma and H.J. Jongsma, 1998. Tumour necrosis factor α alters the expression of Connexin43, Connexin40 and Connexin37, in human umbilical vein endothelial cells. Cytokine, 10: 258-264.
CrossRefDirect Link - Ebong, E.E., S. Kim and N. DePaola, 2006. Flow regulates intercellular communication in HAEC by assembling functional Cx40 and Cx37 gap junctional channels. Am. J. Physiol.-Heart Circulat. Physiol., 290: H2015-H2023.
CrossRefDirect Link - Katakami, N., K.Y. Sakamoto, H. Kaneto, M. Matsuhisa and I. Shimizu et al., 2009. Association between the connexin37 polymorphism and peripheral arterial disease in subjects with type 2 diabetes. Diabet. Care, 32: e53-e54.
CrossRefDirect Link - Kobayashi, Y., K. Hoshikuma, Y. Nakano, Y. Yokoo and T. Kamiya, 2001. The positive inotropic and chronotropic effects of evodiamine and rutaecarpine, indoloquinazoline alkaloids isolated from the fruits of Evodia rutaecarpa, on the Guinea-pig isolated right atria: Possible involvement of vanilloid receptors. Planta Med., 67: 244-248.
PubMedDirect Link - Deng, P.Y., F. Ye, W.J. Cai, G.S. Tan, C.P. Hu, H.W. Deng and Y.J. Li, 2004. Stimulation of calcitonin gene-related peptide synthesis and release: Mechanisms for a novel antihypertensive drug, rutaecarpine. J. Hypert., 22: 1819-1829.
Direct Link - Hu, C.P., L. Xiao, H.W. Deng and Y.J. Li, 2002. The cardioprotection of rutaecarpine is mediated by endogenous calcitonin related-gene peptide through activation of vanilloid receptors in Guinea-pig hearts. Planta Medica, 68: 705-709.
PubMedDirect Link - Wang, S., S. Yamamoto, Y. Kogure, W. Zhang, K. Noguchi and Y. Dai, 2016. Partial activation and inhibition of TRPV1 channels by evodiamine and rutaecarpine, two major components of the fruits of Evodia rutaecarpa. J. Natural Prod., 79: 1225-1230.
CrossRefDirect Link - Wu, Z.Z. and H.L. Pan, 2007. Role of TRPV1 and intracellular Ca2+ in excitation of cardiac sensory neurons by bradykinin. Am. J. Physiol.-Regulat. Integrat. Comparat. Physiol., 293: R276-R283.
CrossRefDirect Link - Xu, Q., R.F. Kopp, Y. Chen, J.J. Yang, M.W. Roe and R.D. Veenstra, 2012. Gating of connexin 43 gap junctions by a cytoplasmic loop calmodulin binding domain. Am. J. Physiol.-Cell Physiol., 302: C1548-C1556.
CrossRefDirect Link - Wang, G.J., X.C. Wu, C.F. Chen, L.C. Lin, Y.T. Huang, J. Shan and P.K. Pang, 1999. Vasorelaxing action of rutaecarpine: Effects of rutaecarpine on calcium channel activities in vascular endothelial and smooth muscle cells. J. Pharmacol. Exp. Therapeut., 289: 1237-1244.
Direct Link