
Guofeng Zhang
College of Environment and Mapping, China University of Mining and Technology, 221116 Xuzhou City, Jiangsu Province, China
LiveDNA: 86.21266
Shengxue Zhou
College of Chinese Medicine, Jilin Agricultural Science and Technology College, 132101 Jilin City, Jilin Province, China
LiveDNA: 86.16926
Hai Zhao
Reconstructive and Plastic Surgery, the General Hospital of Shenyang Military, 110084 Shenyang city, Liaoning Province, China
Guangsheng Xi
College of Chinese Medicine, Jilin Agricultural Science and Technology College, 132101 Jilin City, Jilin Province, China
LiveDNA: 86.20719
Yugang Gao
College of Chinese Medicine, Jilin Agricultural University, 130118 Changchun city, Jilin province, China
Yunjia Wang
College of Environment and Mapping, China University of Mining and Technology, 221116 Xuzhou City, Jiangsu Province, China
LiveDNA: 86.16525
International Journal of Pharmacology
Year: 2018 | Volume: 14 | Issue: 5 | Page No.: 615-623







ABSTRACT
Background and Objective: Ginsenosides are considered as key components to mediate the pharmacological activity of the ginseng. It is great significance for obtaining ginseng extract containing high concentration of ginsenosides. This study aimed to investigate the effects of rolC gene on ginsenosides content in ginseng hairy roots. Materials and Methods: Genomic DNA was extracted from ginseng hairy roots of Panax ginseng C.A. Meyer (P. ginseng) and rolC gene was cloned for pMD18-T/rolC recombinant plasmid construction. Then Agrobacterium rhizogenes A4 harboring rolC gene was used to infect the ginseng hairy roots obtained after inoculating root of P. ginseng. The mRNA and protein of rolC gene were then detected and the different types of ginsenosides were evaluated. Results: A 543 bp length DNA was cloned from ginseng hairy roots and pMD18-T/rolC recombinant plasmid was successfully constructed for rolC gene introduction. Increased growth ratio was found in rolC gene expressed ginseng hairy roots than 3 years old ginseng roots. Moreover, the highest total ginsenosides level (16.13 mg g–1) was found in ginseng hairy roots at 4 weeks, which was increased to 355% than 3 years old ginseng (4.54 mg g–1). Increased levels of both ginsenosides Rb (Rb1, Rb2, Rc, Rd) and Rg (Re, Rf and Rg1) subgroup were found from 2-8 weeks in ginseng hairy roots and 4 week was the optimal time point for the highest ginsenosides content. Conclusion: Introduction of rolC gene could promote ginsenosides content in ginseng hairy roots, which would be beneficial for obtaining ginseng extract containing high concentration of ginsenosides.
PDF Abstract XML References Citation
Received: September 18, 2017;
Accepted: November 20, 2017;
Published: June 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Guofeng Zhang, Shengxue Zhou, Hai Zhao, Guangsheng Xi, Yugang Gao and Yunjia Wang, 2018. High Levels of Ginsenosides Production Associated with rolC Gene Transcription and Expression in Ginseng Hairy Roots. International Journal of Pharmacology, 14: 615-623.
DOI: 10.3923/ijp.2018.615.623
URL: https://scialert.net/abstract/?doi=ijp.2018.615.623
DOI: 10.3923/ijp.2018.615.623
URL: https://scialert.net/abstract/?doi=ijp.2018.615.623
INTRODUCTION
Panax ginseng C.A. Mey (P. ginseng), belongs to family Araliaceae, is a perennial herb native to China, East Asia and Southeast Asia and has been employed as an herbal remedy for thousands of years. Ginseng exhibits multiple pharmacological effects, including anti-fatigue, anticancer, cardioprotective, immunomodulatory and anti-oxidant properties1-3. A number of ginsenosides as triterpene saponins were existed in ginseng and an abundance of saponins are presented in the roots4, including ginsenosides Rb1, Rb2, Rc, Rd, Re, Rf and Rg1 and these ginsenosides were believed to be involved in the pharmacological activity of the ginseng5. However, inadequate amount of ginsenosides in ginseng extract has hindered their employment for treatment purpose. Therefore, the development of ginseng extract containing an increased concentration of ginsenosides is of great interest.
The rolC gene derived from plant pathogen Agrobacterium rhizogenes, played an essential role in the development of hairy root disease and its expression in plants could result in substantial morphological and biochemical alterations6,7. Besides these known properties, Gorpenchenko et al.8, recently discovered a function of the rolC gene in formation of shoot meristems, indicating that it might play a critical role in pluripotent stem cells formation. Moreover, the rolC gene can also affect embryo development in normal animal (sea urchin), which can result in formation of teratoma-like structures in transformed embryos9. Shou-Jing et al.10 reported the rolC gene affecting the synthesis of ginsenosides could be amplified and cloned from the ginseng hairy roots by PCR. However, the effect of exogenous introduction of rolC gene on the ginsenosides content in ginseng hairy root has not yet been explored. Since the role of the rolC gene in hairy roots, it was hypothesized that relationship could be existed between the rolC gene level and ginsenosides concentration.
The present study, aimed to investigate the role of rolC gene on ginsenosides content in ginseng hairy roots, thereby seeking the way to improve the ginsenosides content in ginseng. The rolC gene recombinant expression plasmid was constructed for ginseng roots infection and the effects of rolC gene infection on the ginsenosides content were then evaluated. The results showed that rolC gene introduction could result in increased ginsenosides content in ginseng hairy roots and 4 weeks was confirmed as the optimal time for ginseng collection.
MATERIALS AND METHODS
Plant material: Two month old P. ginseng roots were served as the source of explants for hairy roots induction. These ginseng roots were wounded with sterilized needles and then infected with Agrobacterium rhizogenes (A. rhizogenes) strain A4 for 24 h. These inoculated explants were maintained at 25°C in order to induce the hairy roots and sub-cultured at 30 days intervals. The initial weight of inoculated ginseng roots were about 4 g and the weight of these explants after root induction was measured every 2 weeks under the sterile condition (Fresh weight). Three year old P. ginseng plants were obtained from Jilin Agricultural University, Changchun, China.
Polymerase chain reaction (PCR) analysis for determining rolC gene: Genomic DNA was extracted from hairy roots using the cetyl trimethyl ammonium bromide (CTAB) method11. These genomic DNA was then employed as template for PCR analysis of rolC gene. The forward and reverse primer for rolC gene was 5’ATGGCTGAAGACGACTTGTGTTC3’ and 5’TTAGCCGATTGCAAACTT3’, respectively, which could generate a 543 length PCR product. PCR reaction was composed as follows: 200 ng genomic DNA, 0.4 μM of each primer, 400 μM dNTPs (Dingguo Biotechnology, Beijing, China), 1.0 Unit Taq polymerase (TaKaRa Biotechnology Co., Ltd., Dalian, China) and 2.5 μL 10×PCR buffer into a final volume of 25 μL. The condition for the rolC gene amplification was as follows: Initial denaturation of template DNA at 96°C for 5 min, followed by 30 cycles of 94°C for 45 sec, annealing step at 50°C for 50 sec and extension step at 72°C for 70 sec and an extra cycle with a 5 min extension step at 72°C was finally performed. PCR products were separated on 1.0% (w/v) agarose gels at 65 V for 26 min and visualized with ethidium bromide staining. The DNA size marker was purchased from TaKaRa (DL 2000, TaKaRa Biotechnology Co Ltd., Dalian, China).
Construction and characterization of rolC gene recombinant plasmid: The PCR product of rolC gene was purified using Vetagene Kit (Baotaike biotechnology Co., Ltd., Changchun, China) following the manufacturer’s instructions. Cloning was performed as described by Widersten12. Briefly, PCR product and pMD18-T vector was digested with EcoRI enzyme and ligated using ligases (Takara). Then the ligation products were transformed into E. coli strain JM109 (Promega, Shanghai China) and subjected to Blue-White screening by employing X-gal and IPTG. The white clone was selected and grown for plasmid characterization. After plasmid DNA extraction by the alkaline lysis method described by Birnboim and Doly13, rolC gene was detected by PCR analysis using the same primer described above. Finally, the plasmids with right size PCR product of rolC gene were used for DNA sequencing.
RT-PCR analysis of rolC gene transcript: RT-PCR was carried out to confirm the transcription of rolC gene in ginseng hairy roots. Total mRNA was extracted from ginseng hairy roots and 3 years old ginseng roots using the method described by Willey et al.14. Briefly, fresh ginseng hairy roots and 3 years old ginseng roots were collected and grinded into fine power in liquid nitrogen. mRNA was then isolated using the RNAGENTS total RNA isolation system kit (Z5110, Promega, USA) according to the manufacturer’s instruction. mRNA quality was assessed by electrophoresis on 1.2% formaldehyde/agarose gels and ethidium bromide staining. A total of 1 μg of mRNA from each sample was used for complementary DNAs synthesis and the RT system was as follows: 2 μL 10×RT buffer, 100 μM each type of 4 dNTPs, 0.1 μM of each of primer, 40 Units of RNase inhibitor and 1 Unit AMV transcriptase and the RT reaction condition was 25°C for 10 min, 42°C for 60 min and 92°C for 10 min. The cDNA products were stored in -20°C until further use. The PCR analysis of the rolC gene was performed with the same primers described above. To avoid sample contamination by plasmid DNA, all RNA samples were treated with DNase before PCR analysis.
SDS-polyacrylamide gels (SDS-PAGE) analysis of rolC protein: Approximately 0.05-0.1g of the ginseng hairy roots and 3 years old ginseng roots was grinded with liquid nitrogen and homogenized with appropriate volumes of extraction buffer that consisted of 125 mM Tris-HCl (pH 6.8), 5% β-mercaptoethanol, 4 M urea and 4% SDS. The protein solution was further incubated in dark at 50°C on a rotary shaker with a shaking speed of 80 rpm for 1 h. After that, these solution was centrifuged at 12,000 rpm for 20 min at 4°C and then proteins were precipitated by addition of 4 times volume of acetone according to previous description15. For SDS-PAGE, 3 mg of total proteins samples were concentrated, desalted, analyzed using the Bio-Rad ini-PROTEA Electrophoresis system (Bio-Rad, USA) according to the manufacturer’s instruction. A SDS-PAGE gel with 4% stacking and 10% separation gel was employed for protein electrophoresis and visualized after staining with Coomassie Brilliant Blue (CBB) R-250.
Ginsenosides extraction and analysis: Isolation and quantification of ginsenosides were carried out according to the procedure described previously16. Fully dried 3 g fresh induced ginseng hairy roots and 3 years old ginseng roots were grinded with liquid nitrogen into powder and dissolved in 50 mL methanol. After 30 min sonication, these roots were filtered and 25 filtered solution was dried in water bath. The residue was dissolved in 40 mL butanol-H2O and exacted with separatory funnel for 4 times. All the butanol-H2O exacted solution was combined and evaporated to dryness. The butanol residue was dissolved in methanol (10 mg mL–1), vortex and filtered through 0.45 μm filter membrane and 10 μL dissolved substance was examined by HPLC [column: ODS 5 μm, 4.6×250 mm (Capcell PAK, Shiseido, Japan), eluant: CH3CN-H2O (gradient from 1:4 to 3:2) at a flow rate of 100 μL min–1, detection: 204 nm].
Determination of growth ratio: Fresh weight was measured after washing three times with tap water and blotting away surface water. Dry weight (DW) was recorded after roots were dried at 75 for 3 days. Growth ratio was calculated from equation:
Data analysis: All experiments were conducted at least in triplicate and the results were expressed as the mean values. All the statistical analyses were performed using SPSS software version 18.0 (SPSS Inc., Chicago, IL). Student t-test and one-way ANOVA assay was employed for comparison of the differences between group and multiple groups. p<0.05 was considered as statistical significance.
RESULTS
Isolation and characterization of rolC gene: Genomic DNA extracted from Ginseng hair roots of P. ginseng was used as the template DNA for rolC gene cloning (Fig. 1a) and a 543 bp length fragment was amplified using specific primer for rolC gene (Fig. 1b).
Characterization of pMD18-T/rolC recombinant plasmid: The pMD18-T/rolC recombinant plasmid was constructed using enzyme digest of pMD18-T vector and the 543 bp length rolC gene fragment. After ligation, transformation, clone selection, plasmid extraction and characterization, several positive pMD18-T/rolC recombinant plasmid clones (Lane 2, 3, 5, 7-11 in Fig. 1c) were successfully obtained and agarose gel electrophoresis confirmed that the size of DNA fragment was 543-bp (Fig. 1d).
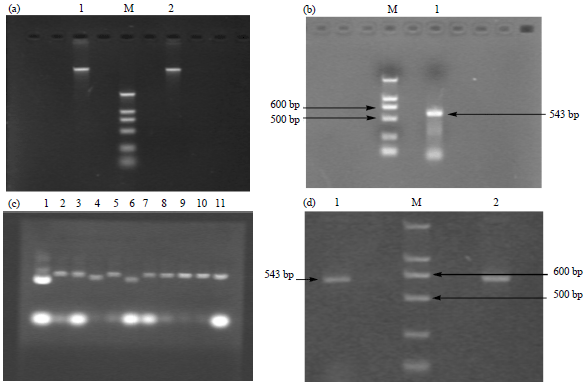 | |
| Fig. 1(a-d): | pMD18-T/rolC recombinant plasmid construction and characterization, (a) Genomic DNA was extracted from ginseng hairy roots and characterized by electrophoresis. Lane M, DNA marker DL2000, Lanes 1 and 2: Genomic DNA, (b) rolC gene was amplified by using genomic DNA template of ginseng hairy roots and characterized by agarose gel electrophoresis. A 543 bp length DNA fragment was confirmed by agarose gel electrophoresis. Lane M: DNA marker DL2000, Lane 1: rolC gene PCR product, (c) Positive clone was selected and plasmid DNA was exacted from these clones for characterization. According to the electrophoresis results, Lane 2, 3, 5 and 7-11 were confirmed as the positive clone while Lane 1, 4 and 6 were negative clones, (d) PCR characterization of recombinant pMD18-T/rolC plasmid. A 543 bp length DNA fragment was confirmed by agarose gel electrophoresis. Lane M: DNA marker DL2000, Lane 1and 2: rolC gene PCR product |
Since the right size of rolC gene was identified by agarose gel electrophoresis, gene sequencing was further performed to confirm that the amplified rolC fragment showed 99% homology of nucleotide sequence to previous reported nucleotide sequence (Fig. 2a). Only 2 positions were confirmed as mutations, which was located at 16 and 128 nt, respectively. The 16 nt mutation was C to T silent mutation, whereas the 128 nt mutation was T to A, resulting in that valine was replaced by glutamic acid (Fig. 2b).
Analysis of rolC gene transcription and translation: In order to analyze the rolC gene transcription, the tissues from ginseng hairy roots and 3 years old ginseng roots were collected to analyze the level of rolC gene using RT-PCR analysis. As shown in Fig. 3a, total RNA was successfully extracted from both ginseng hairy roots and 3 years old ginseng roots. Moreover, a higher level of rolC was found in ginseng hairy roots compared to 3 years old ginseng roots (Fig. 3b).
The protein expression of rolC was also determined using SDS-PAGE assay. As shown in Fig. 3c, the expression of rolC protein was verified in both ginseng hairy roots and 3 years old ginseng roots and the molecular weight of the protein coded by rolC gene was approximately 20 kDa, which was consistent to previous description17.
Ginsenoside content of hairy roots: In order to verify the relationship between the rolC gene and the ginsenoside content from hairy roots, growth rate determination and quantification of different types of ginsenoside were carried out. The growth rate results of ginseng hairy roots was listed in Table 1, fastest ginseng hairy roots growth was found between the time periods of 2nd and 6th weeks, which showed more than 30 times growth according to the fresh weight and dry weight compared to original inoculum (Table 1).
Rb group (Rb1, Rb2, Rc, Rd) and Rg group (Re, Rf and Rg1) were extracted and quantified from the ginseng hairy roots and 3 years old ginseng roots.
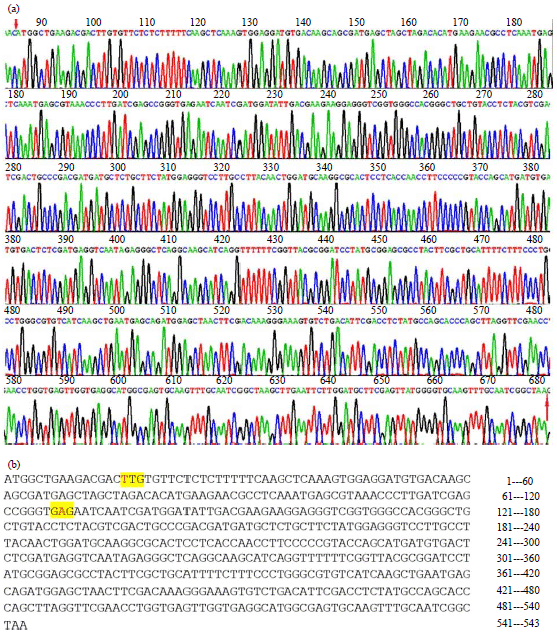 | |
| Fig. 2(a-b): | DNA sequencing of the pMD18-T/rolC recombinant plasmid, (a) Original DNA sequencing results of the pMD18-T/rolC recombinant plasmid. From number 86 to number 681 and (b) Exact results of rolC gene sequencing, according to the results, the red color represents mutated nucleotides. The C base at position 16 was replaced by T while the T base at position 128 was replaced by A |
| Table 1: | Growth rate of ginseng hairy roots from 2-8 week |
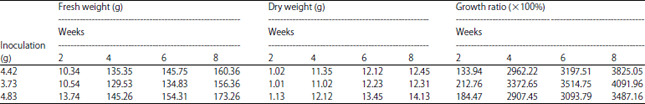 | |
The highest total ginsenosides level (16.13mg g–1 d.wt.) was found in the ginseng hairy roots at 4 weeks and amount of the total ginsenosides was reached to a number of 355% compared to 3 years cultivated ginseng roots (4.54 mg g–1 d.wt.). Significantly increased ginsenosides was also found in the ginseng hairy roots at 8 weeks, which showed a 279% increasing. The content of ginsenosides subtype Rb2, Rc and Rd showed a significant increasing (about 10 times) at 4 weeks compared to those in 3 years old ginseng roots.
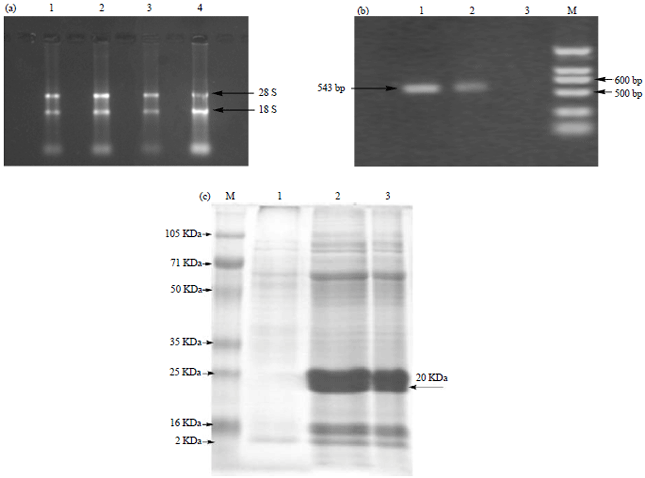 | |
| Fig. 3(a-c): | RT-PCR characterization of the rolC genes in ginseng hairy roots and 3 years old ginseng roots, (a) Total RNA was extracted from ginseng hairy roots and 3 years old ginseng roots and characterized by agarose gel electrophoresis. Lanes 1-3: Total RNA from ginseng hairy roots, Lane 4: Total RNA from 3 years old ginseng roots, (b) RT-PCR detection of rolC gene transcripts from ginseng hairy roots and 3 years old ginseng roots. A 543 bp length transcripts were confirmed by agarose gel electrophoresis. Lane M, DNA marker DL2000, Lanes 1 and 2, cDNA of ginseng hairy roots, Lane 3, cDNA of 3 years old ginseng roots and (c) Protein gel characterization of rolC gene protein from ginseng hairy roots and 3 years old ginseng roots. A 20 kDa protein band was confirmed by SDS-PAGE gel electrophoresis. Lane M, protein marker. Lane 1, protein from 3 years old ginseng roots, Lanes 2 and 3, protein from hairy roots of ginseng roots. |
| Table 2: | Content of different ginsenoside content from hairy roots and 3-year old ginseng roots |
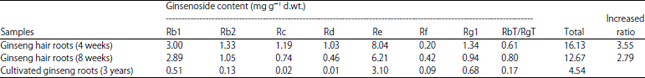 | |
Data was expressed as mg ginsenosides/g dry weight. In all the samples, the fresh weight was 0.5±0.05 g, the value was mean number obtained from 5 replicates of each sample | |
Moreover, the ginsenosides Re was the main type of ginsenosides and the minimum amount of Re was 8.04 and 3.10 mg g–1 d.wt., in ginseng hairy roots at 4 weeks and 3 years old ginseng roots, respectively. Although the best RbT/RgT was at 4-8 weeks was confirmed as the optimal time point due to absolute Re content (Table 2).
DISCUSSION
In the present study, the effects of rolC gene on ginsenosides content were investigated. The results showed that introduction of rolC gene into ginseng hairy roots resulted in an increasing content of ginsenosides, including ginsenosides Rb (Rb1, Rb2, Rc, Rd) and Rg (Re, Rf and Rg1) subgroup, from 2-8 weeks in ginseng hairy roots. Moreover, 4 weeks was confirmed as the optimal time point for the highest ginsenosides content.
There were four types of bacterium-plant interaction, including pathogenic, associative and symbiotic18. Plant growth promoting rhizobacteria (associative bacteria) were featured by their ability to colonize plant root without causing disease and to confer benefit to the plant19. In general, pathogens suppression could be brought by this benefit and generation of antifungal metabolites (for example secondary metabolites) is one of these biocontrol traits20. Rhizobia are symbiotic soil bacteria of the family Rhizobiaceae. Transient or localized plant defense responses after Rhizobial stimulation was reported in various legumes, especially during ineffective symbioses21. However, an recent evidence also indicated that plant defense repress reaction could be generated by a single genes of Rhizobium22. Bartsev et al.23 found that the nopL gene of Rhizobium species NGR234 could block inducible plant defense responses. Taken together, A. rhizogenes, a typical plant pathogen, could be used as a tool for alternation of secondary metabolism in hairy root cultures. Here, Rhizogenes A4 harboring rolC gene was used to infect the ginseng hairy roots obtained after inoculating ginseng root of Panax ginseng C.A. Meyer and RT-PCR and SDS-PAGE results confirmed the transcription and translation of the rolC gene in ginseng hairy roots.
The pharmacological active agents of ginseng were identified on the presence of a mixture of triterpene glycosides, which were known as ginsenosides and reported to exert anticancer, anti-fatigue, cardioprotective, immune-modulatory and anti-oxidant properties24. To date, more than 30 different ginsenosides was isolated and characterized from P. ginseng25. There are two main type of ginsenosides, including the Rb and Rg groups, which respectively possesses 20 (S) protopanaxadiol and 20 (S) protopanaxatriol moieties as the sapogenines26. Rb group ginsenosides including Rb1, Rb2, Rc, Rd and Rg groups including Re, Rf and Rg1, were the most abundant components that were involved in the pharmacological activity of the plant material27. Since the observation of increased growth ratio in ginseng hairy roots, the content of total ginsenosides and each type of Rb and Rg group ginsenosides were further examined. The results showed that the highest total ginsenosides level (16.13 mg g–1) was found in ginseng hairy roots at 4 weeks, which was increased to 355% than 3 years old ginseng (4.54 mg g–1). Moreover, increased level of both ginsenosides Rb (Rb1, Rb2, Rc, Rd) and Rg (Re, Rf and Rg1) subgroup were found from 2-8 weeks in ginseng hairy roots. Also, after calculation, 4 weeks was confirmed as the optimal time point for highest ginsenosides content, which may be further employed for ginseng hairy roots collection. In addition, previous studies also demonstrated a stimulatory effect of rolC gene on secondary metabolism in different groups of secondary metabolites, including tropane alkaloids28,29, pyridine alkaloids30, indole alkaloids31, ginsenosides7 and anthraquinones32. However, Bulgakov et al. showed decreased levels of antimicrobial polyphenols (rabdosiin and rosmarinic acid) and naphthoquinones (shikonin derivatives) in rolC-transformed Eritrichium sericeum and Lithospermum erythrorhizon cell cultures33. Data suggested that A. rhizogenes could perturb secondary metabolism of host plants via a T-DNA oncogene-mediated signaling mechanism. Result declared that after rolC gene introduction, increased growth ratio was found in rolC gene expressed ginseng hairy roots than 3 years old ginseng roots, suggesting that rolC gene could affect the growth of ginseng hairy roots but not aged ginseng roots. Since the confirmation of rolC gene over expression, the exact expression level of rolC gene was not measured and only the levels of different type of ginsenosides were identified. Further study to quantification of rolC gene level in these ginseng hairy roots is needed for exploration of the relationship between rolC gene level and ginsenosides content.
CONCLUSION
Taken together, the results of this study revealed that introduction of rolC gene could promote ginsenosides content in ginseng hairy roots, which would be beneficial for obtaining ginseng extract containing high concentration of ginsenosides. Due to increased level of industrialization, decreased farm land could result in decreased level of ginseng and ginsenosides. The rolC gene of A. rhizogenes possessed with an environment friend role may be a very interesting candidate for biotechnological secondary metabolism research of ginsenosides biosynthesis.
SIGNIFICANCE STATEMENT
This study discovers the effect of rolC gene introduction on ginsenosides content in ginseng hairy roots and the optimal time for ginseng collection that can be beneficial for obtaining ginseng extract containing high concentration of ginsenosides. This study help the researchers to uncover the crucial roles of rolC gene in regulating ginsenosides content in ginseng hairy roots that many researchers were not able to explore. Therefore, a new theory on the relationship between rolC on ginsenosides content is expected to be provided.
ACKNOWLEDGMENTS
This study was supported by National Natural Science Foundation of China subsidization project study on insect resistance of ginsenosides and its effect on the evolution of environmental suitability of ginseng (No. 31470420), Chinese Medicine Key advantages of the provincial subjects of Jilin Agricultural Science and Technology College in Jilin province Screening and chemical modification of Chinese medicine target active ingredient against cerebral ischemic stroke disease (JNYHZ 2015-X035), National Program on Key Research Project: Linxia shen New product development and utilization technology (2016YFC0500303 2016-2020 ), Chinese Medicine Key advantages of the provincial subjects of Jilin Agricultural Science and Technology College in Jilin province: Study on Soil Remediation Technology of conversion of ginseng-plant to Forestland in Eastern Mountainous Area of Jilin Province JNYHZ 2015-X038.
REFERENCES
- De Wang, X., G.Y. Su, C. Zhao, F.Z. Qu, P. Wang and Y.Q. Zhao, 2016. Anticancer activity and potential mechanisms of 1C, a ginseng saponin derivative, on prostate cancer cells. J. Ginseng Res.
CrossRefDirect Link - Park, S.B., J.H. Chun, Y.W. Ban, J.Y. Han and Y.E. Choi, 2016. Alteration of Panax ginseng saponin composition by overexpression and RNA interference of the protopanaxadiol 6-hydroxylase gene (CYP716A53v2). J. Ginseng Res., 40: 47-54.
CrossRefDirect Link - Ha, L.T., N. Pawlicki-Jullian, M. Pillon-Lequart, M. Boitel-Conti, H.X. Duong and E. Gontier, 2016. Hairy root cultures of Panax vietnamensis, a promising approach for the production of ocotillol-type ginsenosides. Plant Cell Tiss. Organ Cult., 126: 93-103.
CrossRefDirect Link - Lee, C.H. and J.H. Kim, 2014. A review on the medicinal potentials of ginseng and ginsenosides on cardiovascular diseases. J. Ginseng Res., 38: 161-166.
CrossRefDirect Link - Grishchenko, O.V., K.V. Kiselev, G.K. Tchernoded, S.A. Fedoreyev, M.V. Veselova, V.P. Bulgakov and Y.N. Zhuravlev, 2016. RolB gene-induced production of isoflavonoids in transformed Maackia amurensis cells. Applied Microbiol. Biotechnol., 100: 7479-7489.
CrossRefDirect Link - Matveeva, T.V., S.V. Sokornova and L.A. Lutova, 2015. Influence of Agrobacterium oncogenes on secondary metabolism of plants. Phytochem. Rev., 14: 541-554.
CrossRefDirect Link - Gorpenchenko, T.Y., K.V. Kiselev, V.P. Bulgakov, G.K. Tchernoded and E.A. Bragina et al., 2006. The Agrobacterium rhizogenes rolC-gene-induced somatic embryogenesis and shoot organogenesis in Panax ginseng transformed calluses. Planta, 223: 457-467.
CrossRefDirect Link - Bulgakov, V.P., K.V. Kisselev, K.V. Yakovlev, Y.N. Zhuravlev, A.A. Gontcharov and N.A. Odintsova, 2006. Agrobacterium-mediated transformation of sea urchin embryos. Biotechnol. J., 1: 454-461.
CrossRefPubMedDirect Link - Shou-Jing, Z., L. Chang-Yu, L. Qing-Chang, Q. Yan-Chun and S. Jing, 2004. Cloning and sequence analysis of rolC gene affecting ginsenoside content in hairy roots of Panax ginseng CA Mey. Chinese J. Agric. Biotechnol., 1: 61-65.
Direct Link - Healey, A., A. Furtado, T. Cooper and R.J. Henry, 2014. Protocol: A simple method for extracting next-generation sequencing quality genomic DNA from recalcitrant plant species. Plant Meth., Vol. 10.
CrossRefDirect Link - Widersten, M., 2014. Protein engineering for development of new hydrolytic biocatalysts. Curr. Opin. Chem. Biol., 21: 42-47.
CrossRefDirect Link - Birnboim, H.C. and J. Doly, 1979. A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res., 7: 1513-1523.
CrossRefPubMedDirect Link - Marsoni, M., C. Vannini, M. Campa, U. Cucchi, L. Espen and M. Bracale, 2005. Protein extraction from grape tissues by two-dimensional electrophoresis. Vitis, 44: 181-186.
Direct Link - Li, Z., R. Zhang, X. Wang, X. Hu, Y. Chen and Q. Liu, 2015. Simultaneous determination of seven ginsenosides in rat plasma by high-performance liquid chromatography coupled to time-of-flight mass spectrometry: Application to pharmacokinetics of Shenfu injection. Biomed. Chromatogr., 29: 167-175.
CrossRefDirect Link - Nabiabad, H.S., K. Piri and M. Amini, 2016. Expression of active chimeric-tissue plasminogen activator in tobacco hairy roots, Identification of a DNA aptamer and purification by aptamer functionalized-MWCNTs chromatography. Protein Expression Purif.
CrossRefDirect Link - Hardoim, P.R., L.S. van Overbeek, G. Berg, A.M. Pirttila and S. Compant et al., 2015. The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev., 79: 293-320.
Direct Link - Kiselev, K.V., M.I. Kusaykin, A.S. Dubrovina, D.A. Bezverbny, T.N. Zvyagintseva and V.P. Bulgakov, 2006. The rolC gene induces expression of a pathogenesis-related β-1, 3-glucanase in transformed ginseng cells. Phytochemistry, 67: 2225-2231.
CrossRefDirect Link - Schillinger, W.F. and T.C. Paulitz, 2014. Natural suppression of Rhizoctonia bare patch in a long-term no-till cropping systems experiment. Plant Dis., 98: 389-394.
CrossRefDirect Link - Moreau, S., J. Fromentin, F. Vailleau, T. Vernie and S. Huguet et al., 2014. The symbiotic transcription factor MtEFD and cytokinins are positively acting in the Medicago truncatula and Ralstonia solanacearum pathogenic interaction. New Phytol., 201: 1343-1357.
CrossRefDirect Link - Breakspear, A., C. Liu, S. Roy, N. Stacey and C. Rogers et al., 2014. The root hair "infectome" of Medicago truncatula uncovers changes in cell cycle genes and reveals a requirement for auxin signaling in rhizobial infection. Plant Cell, 26: 4680-4701.
CrossRefPubMedDirect Link - Bartsev, A.V., W.J. Deakin, N.M. Boukli, C.B. McAlvin and G. Stacey et al., 2004. NopL, an effector protein of Rhizobium sp. NGR234, thwarts activation of plant defense reactions. Plant Physiol., 134: 871-879.
Direct Link - Kim, H.J., P. Kim and C.Y. Shin, 2013. A comprehensive review of the therapeutic and pharmacological effects of ginseng and ginsenosides in central nervous system. J. Ginseng Res., 37: 8-29.
Direct Link - Wan, J.Y., Y. Fan, Q.T. Yu, Y.Z. Ge and C.P. Yan et al., 2015. Integrated evaluation of malonyl ginsenosides, amino acids and polysaccharides in fresh and processed ginseng. J. Pharm. Biomed. Anal., 107: 89-97.
CrossRefDirect Link - Wanderi, C., E. Kim, S. Chang, C. Choi and K. Choi, 2016. Ginsenoside 20 (S)-protopanaxadiol suppresses viability of human glioblastoma cells via down-regulation of cell adhesion proteins and cell-cycle arrest. Anticancer Res., 36: 925-932.
Direct Link - Chen, C.F., W.F. Chiou and J.T. Zhang, 2008. Comparison of the pharmacological effects of Panax ginseng and Panax quinquefolium. Acta Pharmacol. Sin., 29: 1103-1108.
PubMedDirect Link - Vereshchagina, Y.V., V.P. Bulgakov, V.P. Grigorchuk, V.G. Rybin and G.N. Veremeichik et al., 2014. The rolC gene increases caffeoylquinic acid production in transformed artichoke cells. Appl. Microbiol. Biotechnol., 98: 7773-7780.
CrossRefDirect Link - Almagro, L., F. Fernandez-Perez and M.A. Pedreno, 2015. Indole alkaloids from Catharanthus roseus: Bioproduction and their effect on human health. Molecules, 20: 2973-3000.
CrossRefDirect Link - Xu, A., J.C. Zhan and W.D. Huang, 2015. Effects of ultraviolet C, methyl jasmonate and salicylic acid, alone or in combination, on stilbene biosynthesis in cell suspension cultures of Vitis vinifera L. cv. Cabernet Sauvignon. Plant Cell. Tissue Organ Cult., 122: 197-211.
CrossRefDirect Link