
Wei Hu
Provincial Key Laboratory of Developmental Biology and Neuroscience, Fujian Normal University, 350108 Fujian, Fuzhou, China
Tongzhong He
Provincial Key Laboratory of Developmental Biology and Neuroscience, Fujian Normal University, 350108 Fujian, Fuzhou, China
Yuanhui Huo
Provincial Key Laboratory of Developmental Biology and Neuroscience, Fujian Normal University, 350108 Fujian, Fuzhou, China
Yanguo Hong
Provincial Key Laboratory of Developmental Biology and Neuroscience, Fujian Normal University, 350108 Fujian, Fuzhou, China
LiveDNA: 86.22784
International Journal of Pharmacology
Year: 2018 | Volume: 14 | Issue: 5 | Page No.: 601-608







ABSTRACT
Background and Objective: Adrenomedullin (AM) belongs to calcitonin gene-related peptide (CGRP) family. The pain peptide plays a role in pathological pain. This study investigated the possible involvement of AM in bone cancer pain. Methods: Carcinoma cells were injected into the medullary cavity of tibia in rats. Agent was administered intrathecally (i.t.). Nociceptive behaviors were assessed. Neurochemicals in the spinal dorsal horn and dorsal root ganglia (DRG) were assayed by immunohistochemistry and real-time PCR. Results: Carcinoma cells inoculated into the medullary cavity of right tibia induced a reduction of mechanical withdrawal threshold in the hind paw. This was accompanied by an increase in AM expression in small but not medium sized neurons in dorsal root ganglia (DRG). mRNA expression analysis showed that AM was upregulated in the spinal dorsal horn and DRG. Intrathecal (i.t.) administration of the selective AM receptor antagonist AM22-52 (10 nmol) on day 14 postinoculation attenuated bone cancer induced mechanical allodynia and inhibited the increase of AM protein and mRNA in the spinal cord and/or DRG. Bone cancer increased the expression of CGRP mRNA in the DRG but not in the spinal dorsal horn. Cancer cell inoculation induced an increase in neuronal nitric oxide synthesis (nNOS) expression in small but not medium sized neurons in DRG. Furthermore, the increase of CGRP mRNA and nNOS protein in DRG was abolished by i.t. AM22-52 (10 nmol).Conclusion: The present study suggests that the pronociceptive mediator AM was upregulated in autocrine and/or paracrine manners and involved in the pathogenesis of bone cancer pain. The enhanced AM bioactivity can facilitate the expression of nNOS and CGRP mRNA in DRG in bone cancer. Targeting AM receptors should be considered as a novel therapy to treat bone cancer pain.
PDF Abstract XML References Citation
Received: September 07, 2017;
Accepted: November 17, 2017;
Published: June 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Wei Hu, Tongzhong He, Yuanhui Huo and Yanguo Hong, 2018. Involvement of Adrenomedullin in Bone Cancer Pain in Rats. International Journal of Pharmacology, 14: 601-608.
DOI: 10.3923/ijp.2018.601.608
URL: https://scialert.net/abstract/?doi=ijp.2018.601.608
DOI: 10.3923/ijp.2018.601.608
URL: https://scialert.net/abstract/?doi=ijp.2018.601.608
INTRODUCTION
Bone cancer induced pain is common and consists of pain hypersensitivity and ongoing spontaneous pain1. Bone cancer pain is most severe, reduces quality of life and contributes significantly to increased morbidity2. It is an intractable problem in clinic as opiates that are primarily relied to the treatment of this pain are not effective even at a high dose1,3 but produce unacceptable side-effects4. This is partly due to the fact that the underlying mechanisms are unclear but may involve multiple changes including components of inflammatory and neuropathic pain as well as elements that are unique to bone cancer pain5. It is believed that upregulation of pronociceptive mediators in DRG or the spinal cord plays a role in bone cancer pain. The pronociceptive mediators that have been shown to be involved in the pathogenesis of bone cancer pain include CCL26, sodium channels Nav1.8 and 1.97, CGRP8, neuronal nitric oxide synthesis (nNOS)9, protease-activated receptor 2 (PAR2)10 and interleukin-1β11. However, information in this regard is still limited.
Adrenomedullin (AM), an important pain mediator, may be involved in bone cancer pain. AM is a 52 amino acid peptide and a member of calcitonin gene related peptide (CGRP) family12. This peptide is expressed in superficial laminae of the spinal cord and small as well as medium diameter neurons in dorsal root ganglia (DRG)13, the key structures involved in nociceptive processing14. The expression of AM in these tissues is greatly enhanced in inflammatory pain13,15 and morphine tolerance16. An intrathecal (i.t.) administration of the AM receptor antagonist AM22–52 abolishes inflammatory15 and morphine induced16 hyperalgesia, indicating that AM plays a vital role in the induction of pathological pain. Interestingly, AM is in the upstream of cascade induced by these disorders as it recruits CGRP15, nNOS17 and inflammatory cytokines18,19. The present study aimed to investigate the putative role of AM in bone cancer pain and its possible mechanisms.
MATERIALS AND METHODS
Animals: Male Sprague-Dawley rats (250-320 g) supplied by the Animal Center of Fujian Medical University were housed at 22°C with 50% humidity under a 12 h light/dark cycle and given free access to food and water. Care and treatment of animals were performed according to the guidelines for investigations of experimental pain in conscious animals20 and were approved by the Animal Care Committee of Fujian Normal University. All efforts were made to minimize animal suffering and the number of animals used in our experiments.
Inoculation of cancer cells: Tumors were generated by injecting Walker 256 rat mammary gland carcinoma cells (purchased from Baili Biotechnology Company, Shanghai, China) into the abdominal cavity of Sprague-Dawley rats under sodium pentobarbital anesthesia (50 mg kg–1, i.p.). Carcinoma cells (1×108, cancer group) or heat killed carcinoma cells (sham group) in 4 μL PBS were injected into the medullary cavity of right tibia as described previously21. The tibia bone destruction by tumor was confirmed by radiology on day 14 in some rats.
Intrathecal catheter implantation: Animals were implanted with chronic indwelling catheters22. Briefly, rat was injected with i.p. pentobarbital (50 mg kg–1, Shenwgong Co., Shanghai, China) and shaved along the occiput and neck. The dura mater overlying the atlanto-occipital junction was exposed by blunt dissection and an incision was made in the dura. A polyethylene catheter (PE-10, Stoelting, Wood Dale, IL, USA), with a loose knot at 8.0 cm from the end, was threaded caudally to position its tip at the L4-L5 segments of the spinal cord and the knot was immobilized by suturing to the musculature. The rostral tip of the catheter was exteriorized at the back of the neck. The catheter was then flushed with 10 μL of saline and plugged. The rats were housed individually after surgery and allowed to recover for 7 days before being used for behavioral testing. Only the animals with no evidence of neurological deficits after catheter placement were used for the experiment.
Behavioral assessment of nociception: Mechanical threshold was measured in the hind paw (6 in each group) using an automated Von Frey type system (Dynamic Plantar Anesthesiometer 37400, Ugo Basile, Italy). Animals were acclimatized to the testing apparatus and environment for 1 h for 3 days and also habituated on the day of test. Rats were placed on a metal mesh surface under a plastic enclosure. The stimulator unit was placed beneath the right hind paw with the filament below the plantar surface of the rat. Paw withdrawal threshold (PWT) was measured by applying an increasing force (measured in grams) using a stainless-steel filament (0.5 mm diameter). To start, the electrodynamic actuator unit lifted the filament and exerted a force. The force was increased at a rate of 2.5 g sec–1 until the rat moved its paw. A force of 50 g for 30 sec was used as a cut-off point to preclude possible damage to the paw. The force was measured three times at 2 min interval to generate mean value. On day 14 after inoculation of cancer or heat killed carcinoma cells, saline or AM22-52 was administered i.t. The dose of AM22-52 (10 nmol) was referred to the previous studies15,16, PWT was measured for various times. The investigators were blind to the drug test conditions. AM22-52 was purchased from Huadatianyuan Biological Co. (Shanghai, China) and prepared by dissolving in 0.9% sterile saline.
Immunofluorescence: Saline or AM22-52 (10 nmol) was administered in sham or bone cancer rats on day 14 postinoculation (n = 5 each). Forty to sixty min later, rats were deeply anesthetized with sodium pentobarbital (50 mg kg–1 i.p.). Animals were perfused intracardially with cold 0.01 M phosphate buffered saline (PBS) and subsequently with 4% paraformaldehyde in 0.1 M phosphate buffer (PB). DRG at L4-L5 were dissected, post fixed in the same fixative overnight and then transferred to 30% sucrose in PB for cryoprotection. DRG was cut using a cryostat at a 10 μm thickness. Sections were collected on the slides. Immunostaining was performed at room temperature on slides. Tissue sections were incubated in a blocking solution of 0.3% H2O2 and 10% normal goat serum (Vector Laboratories, Burlingame, CA, USA) for 1 h at room temperature. The sections were then incubated with polyclonal rabbit AM (1:50, Santa Cruz Biotechnology Inc., Shanghai, China) or nNOS (1:150, Santa Cruz) antibody overnight at 4oC. The sections were then washed with PBS and incubated with donkey anti-rabbit IgG conjugated with FITC (1:200, Abcam, Shanghai, China) or with TRITC (1:200, Abcam) for 2 h at room temperature. After being thoroughly rinsed with PBS, the sections were dehydrated in a series of graded alcohol, air dried, cleared in xylene and coverslipped. For controls, the primary antibody against AM or nNOS was omitted in the immunostaining protocol. This procedure resulted in negative staining.
The sections were examined with a fluorescence digital microscope (BX51, Olympus, Japan). Images were captured with a Q-Fire cooled camera (DP70, Olympus, Japan). Quantification of AM and nNOS immunoreactivity positive and negative neurons was performed using image analysis software Image-Pro Plus 6.0 (Media Cybernetics, Silver Spring, MD, USA). A field of 210×210 μm was randomly selected from each of 7 DRG sections per animal. To determine the percentage of positive neurons in each DRG, a threshold of average cytoplasmic optical intensity of AM or nNOS immunofluorescence was set using the software. The optical density threshold was then applied to whole DRG section. All neurons sectioned through their nucleus with mean optical density exceeding the threshold were counted as AM or nNOS positive. Otherwise, the cells were considered as negative neurons. The number of positive cells was expressed as: in the same size subpopulation.
For neuron size measurements, the neuronal soma area was calculated by the imaging software. Small neurons were <600 μm2, medium neurons were 600-1200 μm2 and large neurons were >1200 μm2 23.
Quantitative real-time-PCR: The dorsal part of the lumbar (L4-L5) spinal cord and DRG at L4-L5 were harvested 40-60 min after i.t. administration of AM22-52 or saline in the bone cancer or sham group. Tissue samples were collected and frozen immediately in liquid nitrogen, then stored at -80°C for total RNA isolation. Total RNA was extracted using the RNA-prep pure Tissue Kit (Tiangen) according to the manufacturer’s instructions and treated with Deoxyribonuclease I (Tiangen) to remove DNA contamination. RNA concentrations were measured using an Ultraviolet-visible spectrophotometer (NanoDrop 2000, Thermo Scientific, USA). The quality and quantity of the RNA were assessed at 260/280 A and all samples showed absorbency ratios ranging from 1.8-2.0. A total of 1 g of RNA was reverse-transcript using Quant script Reverse Transcription Kit according to the protocol of the manufacturer (Tiangen). All agents were purchased from Tiangen Biochemical Technology (Beijing, China).
Primers for AM, CGRP and GAPDH were designed using Primer 3 software. The oligonucleotide primers were AM (forward), 5’-GTTTCCATCGCCCTGATGTTATT-3’, AM (reverse), 5’-GTAGTTCCCTCTTCCCACGACTTAG-3’, CGRP (forward), 5’-AACCTTAGAAAGCAGCCCAGGCATG-3’, CGRP (forward), 5’-GTGGGCACAAAGTTGTCCTTCACCA-3’, GAPDH (forward), 5’-GTTTGTGATGGGTGTGAAC-3’, GAPDH (reverse), 5’-TCTTCTGAGTGGCAGTGA-3’. After being placed at room temperature, 20 ng of cDNA from the same cDNA batch was subjected to real-time PCR to amplify all genes in triplicate in a total reaction volume of 20 μL using SYBR Premix Ex Taq, ROX as internal reference dye (Takara Biotechnology Technologies, Dalian, China) and the required amount of forward and reverse primers (Jierui Biotechnology Company, Shanghai, China). Reactions were conducted on a 7500 Fast Sequence Detection System (Applied Biosystems, Foster, CA, USA) using following cycling conditions, one cycle at 95°C for 30 sec, followed by 40 cycles at 95°C for 5 sec and 60°C for 30 sec. For each experiment, a non-template reaction served as negative control. Melting curve analysis of products as well as amplicon size verification on a 3% agarose gel confirmed the specificity of the PCR. The raw expression level for each gene was calculated using the same external standard curve made with a mixture of cDNA samples. The target gene expression data were then normalized to GAPDH gene expression to obtain relative concentrations and presented as relative expression units.
Statistical analysis: Data are expressed as mean±standard error of mean (SEM). Statistical significance between groups was examined using one-way analysis of variance (ANOVA). A p-value less than 0.05 was considered statistically significant.
RESULTS
Effect of intrathecal administration of AM22-52 on bone cancer-induced mechanical allodynia: Animals reliably developed hypersensitivity to mechanical stimulus in the hind paw on the bone cancer side, starting on day 4 and maintaining thereafter throughout the experiment (3 weeks) after inoculation of cancer cells but not in the sham group (data not shown). PWT was 26.7±0.6 g on day 14 postinoculation and this was significantly lower than the baseline value of 33.3±0.6 g (p<0.05), indicating mechanical allodynia. Intrathecal administration of AM22-52 at a dose of 10 nmol15,16 increased PWT to 32.3±0.6 g 20 min later. This value was significantly higher than the pre-administration level (p<0.05) and close to the pre surgery level (p>0.05). The increase of mechanical threshold was maintained for approximately 60 min (Fig. 1). However, i.t. administration of saline did not change bone cancer induced mechanical allodynia compared to the pre-administration level (p>0.05).
Expression of AM in DRG with or without intrathecal administration of AM22-52 in bone cancer: As behavioral results suggested the involvement of AM receptor signaling in the pathogenesis of bone cancer pain, the expression of AM at protein level in DRG in bone cancer was examined. As previous studies show autocrine and/or paracrine regulations of AM expression in the spinal cord and DRG in pathological pain15,16, the effect of blockade of AM receptors on AM expression was also determined. Saline or AM22-52 (10 nmol) was administered i.t. on day 14 after postinoculation in sham or bone cancer animals. The DRG at L4-L5 were harvested 40-60 min after the i.t. injection. Each protocol was repeated in 5 animals.
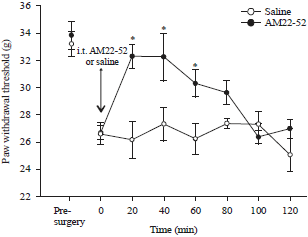 | |
| Fig. 1: | Effect of i.t. administration of AM22-52 on bone cancer induced mechanical hypersensitivity in the hind paw, cancer cells were injected into the medullary cavity of right tibia on day 1 and saline or AM22-52 (10 nmol) was administered i.t. on day 14. PWT was measured |
Data are presented as Mean±SEM, *p<0.05 compared to the pre-administration, N = 6 each | |
Immunofluorescence staining showed that AM was expressed in the soma of small and medium diameter neurons in the DRG. The percentages of AM staining neurons were 22.2±1.2 in small sized population and 70.8±17.2 in medium sized population in the sham group (Fig. 2a). Cancer cell inoculation upregulated the expression of AM to 39.4±1.9 and 77.1±7.9% in small and medium populations, respectively (cancer/saline group, Fig. 2b and d). One-way ANOVA analysis shows that the expression of AM in small (p<0.05) but not medium, population was significantly higher than the sham group. However, AM expressing neurons in small sized population were reduced to 24.0±1.7% in the cancer group treated with AM22-52 which was significantly lower than that in the cancer/saline group (p<0.05, Fig. 2c and d).
Bone cancer induced expression of AM and CGRP mRNA in DRG and the spinal cord with or without AM receptor blockade: The experimental protocol was as the same as that described above and dorsal half of the lumbar spinal cord and DRG at L4-L5 were harvested. mRNA expression analysis revealed that cancer cell inoculation increased the AM mRNA levels by 1.7±0.2 and 2.8±0.5 in DRG and the spinal dorsal horn, respectively (Fig. 3a). These values were significantly higher than those in the sham group (p<0.05). However, following the treatment of AM22-52, the relative expression of AM mRNA was reduced to 1.1±0.2 in DRG and 1.3±0.2 in the spinal cord. These reductions were statistically significant compared to the cancer/saline group (p<0.05).
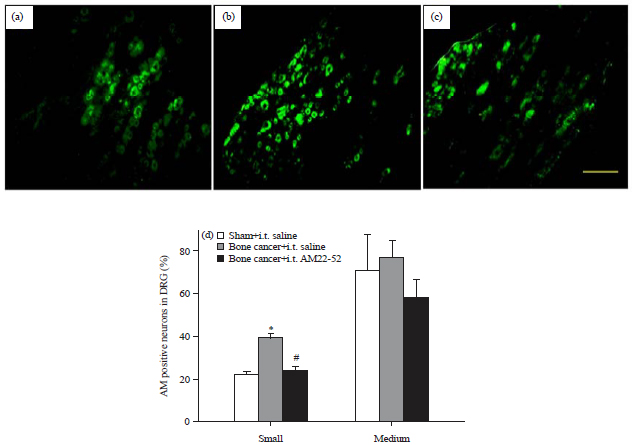 | |
| Fig. 2(a-d): | Effect of i.t. AM22-52 on cancer induced AM expression in DRG, cancer cells or heated cancer cells were injected into the medullary cavity of right tibia on day 1 and saline or AM22-52 (10 nmol) was administered i.t. on day 14, DRG at L4-5 on the inoculation side were harvested 40-60 min later and assayed by immunofluorescence staining technique, representative photomicrographs show the AM immunofluorescence images from the groups of (a) Sham/saline, (b) Bone cancer/saline, (c) Bone cancer/AM22-52 and (d) Quantification of AM immunofluorescence is represented as a percentage of AM positive cells over the total neurons in small or medium sized subpopulations |
*p<0.05 compared to the sham group, #p<0.05 compared to the bone cancer/AM22-52 group, N = 5 each, Scale bar = 50 μm | |
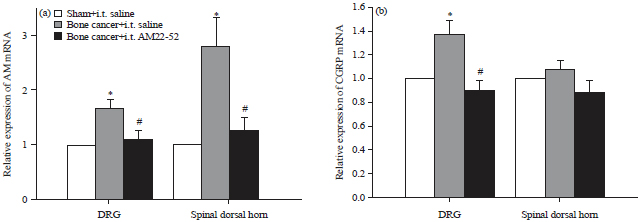 | |
| Fig. 3(a-b): | Effect of i.t. AM22-52 on cancer induced expression of AM and CGRP mRNA in DRG and spinal dorsal horn, cancer cells or heated cancer cells were injected into right tibia on day 1 and saline or AM22-52 (10 nmol) was administered i.t. on day 14, DRG and dorsal half of the lumbar spinal cord were harvested and mRNA levels of (a) AM and (b) CGRP were assayed |
*p<0.05 compared with the sham group, #p<0.05 compared to the bone cancer/AM22-52 group, N = 4 each | |
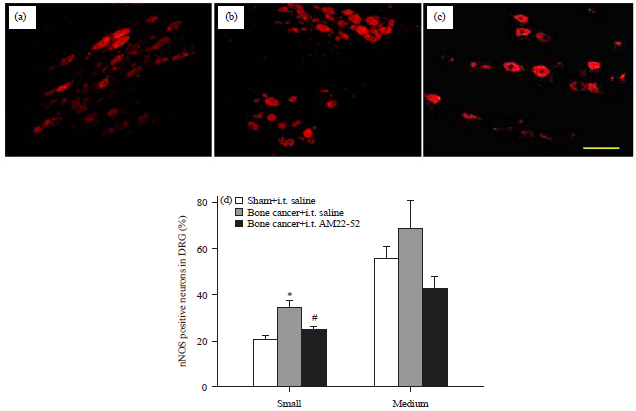 | |
| Fig. 4(a-d): | Effects of i.t. AM22-52 on cancer induced expression of nNOS in DRG. Cancer cells or heated cancer cells were injected into right tibia on day 1 and saline or AM22-52 (10 nmol) was administered i.t. on day 14, DRG at L4-5 on the inoculation side were harvested and assayed by immunofluorescence staining technique, representative photomicrographs show the nNOS immunofluorescence images from the groups of (a) Sham/saline, (b) Bone cancer/saline, (c) Bone cancer/AM22-52 and (d) Quantification of nNOS immunofluorescence is represented as a percentage of nNOS positive cells over the total neurons in small or medium sized subpopulations |
*p<0.05 compared to the sham group, #p<0.05 compared to the bone cancer/AM22-52 group, N = 5 each, Scale bar = 50 μm | |
After cancer cell inoculation, the levels of CGRP mRNA were increased in the DRG by 1.4±0.1 folds (Fig. 3b). This change was statistically significant compared to the sham group (p<0.05). Following the treatment of AM22-52, the cancer induced expression of CGRP mRNA were reduced to 0.9±0.1 (p<0.05 vs cancer/saline). However, cancer cell inoculation only induced CGRP mRNA by 1.1±0.1. This value was very close to the level of CGRP mRNA in the sham group (Fig. 3b).
Expression of nNOS in DRG and its change following intrathecal administration of AM22-52 in bone cancer: To investigate the mechanisms underlying the contribution of AM to cancer pain, the effect of AM receptor blockade on the expression of other pronociceptive mediator was determined in bone cancer. nNOS staining was shown in small (20.8±1.5) and medium sized (55.6±5.6) DRG neurons in sham rats (Fig. 4a and d). Cancer cell inoculation remarkably increased the expression of nNOS in small (34.7±2.8, p<0.05) but not medium (68.8±12.0), sized neurons (Fig. 4b and d). Following i.t. administration of AM22-52, the proportion of nNOS positive neurons in small population was clearly reduced (24.9±1.3). The reduction was statistically significant compared to the cancer/saline group (p<0.05, Fig. 4c and d).
DISCUSSION
The present study demonstrated that the blockade of AM receptors by i.t. administration of the AM receptor antagonist AM22-52 reversed bone cancer induced mechanical allodynia. AM expression at protein and mRNA levels was increased in the spinal cord and/or DRG in bone cancer. These changes were abolished by the i.t. AM22-52. Bone cancer also increased the levels of nNOS and CGRP mRNA in DRG. The increase of all these molecules was abolished by the treatment with AM22-52. These results suggest that enhanced AM bioactivity is involved in the pathogenesis of bone cancer pain.
Mechanical allodynia is a characterized behavioral response for the rat model of bone cancer pain. This behavioral develops in a few days after inoculation of cancer cells in tibia bone and remains thereafter21,24. The present study showed the reversal of bone cancer induced mechanical allodynia on day 14 postinoculation following i.t. administration of the selective AM receptor antagonist AM22-5225. The attenuation of allodynia was seen at 20 min after the administration of AM22-52, the earliest time examined and maintained for 1 h. In contrast, i.t. administration of AM22-52 does not alter the pain threshold in untreated rats16. AM is expressed in small and medium sized neurons in DRG13,15. This study demonstrated that the expression of AM protein was increased only in small but not medium sized neurons. This was slightly different from that seen in inflammatory pain, in which AM expression is increased in both small and medium sized neurons in DRG15. AM mRNA expression was also increased in the spinal dorsal horn and DRG, the important structures involved in nociceptive processing14. The increased AM receptor signaling in the spinal dorsal horn and DRG has been demonstrated in inflammatory pain13,15, neuropathic pain26 and morphine induced hyperalgesia16. Interestingly, the increase of AM protein and mRNA levels was abolished by the blockade of AM receptors. This was correlated with a significant attenuation of mechanical allodynia. These results implied that AM can facilitate its own synthesis or expression by acting on AM receptors in DRG neurons in bone cancer pain. This notion is supported by the anatomical colocalization of AM with AM receptor components, calcitonin receptor like receptor (CLR) and receptor activity modifying protein 2 (RAMP2), in DRG neurons16. Acting via an autocrine or paracrine pathway, AM can induce a positive feedback to stimulate its own production and keep AM concentration at a high level. This regulation also occurs in inflammatory pain13,15 and morphine tolerance16. The present study suggests that enhanced biological activity of AM plays an important role in nociceptive processing in bone cancer.
Previous studies show that AM receptor signaling can trigger a sequence of events leading to pain hypersensitivity. Enhanced AM bioactivity contributing pain hypersensitivity is partly mediated by other pronociceptive mediators, such as CGRP, nNOS15-17 and inflammatory cytokines18,19 in inflammatory pain and morphine tolerance. Therefore, we next determined if the blockade of AM receptors reversed the expression of CGRP and nNOS in bone cancer pain as these are important molecules involved in pathological pain, including cancer pain. Activity evoked release of CGRP is enhanced in superficial lamina of the spinal dorsal horn and CGRP expression is increased in small diameter cell bodies in DRG in cancer pain8,27 while i.t. administration of the CGRP antagonist CGRP8-37 reduces cancer induced referred allodynia8. nNOS is also increased in the spinal cord in cancer pain9 and the inhibition of nNOS by i.t. administration of L-NAMA attenuates cancer pain28. In accordance with previous studies8,27, this study showed the increase of CGRP mRNA expression in DRG but not in the spinal cord. The study further demonstrated that the expression of nNOS was increased in small but not medium, sized neurons in DRG in bone cancer pain. Importantly, the blockade of AM receptors abolished the increase of CGRP mRNA and nNOS expressions in the DRG. In agreement with the suggestions that these pronociceptive mediators were regulated by AM, AM has been demonstrated to be co-localized with CGRP13 and nNOS in DRG17. Therefore, the results in the present study added an evidence to support the notion that AM is in the upstream of a cascade and can recruit other pronociceptive molecules contributing to pathological pain.
CONCLUSION
The increase in the activity of pronociceptive mediators is one of mechanisms underlying bone cancer pain. The present study extends this notion by adding AM, a recently characterized pronociceptive mediator, to the list of molecules involved in the pathogenesis of cancer pain. The key contribution of AM to bone cancer pain was evidenced not only by its upregulation in the spinal dorsal horn and DRG but also by the inhibition of pain and cancer induced increase in AM, CGRP and nNOS following the blockade of AM receptors. These findings suggest that inhibition of AM receptor signaling might be a valuable therapy for bone cancer pain.
SIGNIFICANCE STATEMENT
This study discovers a pharmacological characterization of treatment that can be beneficial for bone cancer pain rats. This study help the researchers to uncover the critical area of pharmacology of a potential therapy for bone cancer pain that many researchers were not able to explore. Thus, a new theory on the mechanism and treatment of cancer pain may be arrived at.
ACKNOWLEDGMENT
This study was supported by the grants from the Natural Science Foundation of China (31371124, 81571084).
REFERENCES
- Portenoy, R.K., D. Payne and P. Jacobsen, 1999. Breakthrough pain: Characteristics and impact in patients with cancer pain. Pain, 81: 129-134.
PubMedDirect Link - Thurlimann, B. and N.D. de Stoutz, 1996. Causes and treatment of bone pain of malignant origin. Drugs, 51: 383-398.
CrossRefDirect Link - Portenoy, R.K., 1999. Managing cancer pain poorly responsive to systemic opioid therapy. Oncology, 13: 25-29.
PubMedDirect Link - Mercadante, S., 1997. Malignant bone pain: Pathophysiology and treatment. Pain, 69: 1-18.
CrossRefDirect Link - Falk, S. and A.H. Dickenson, 2014. Pain and nociception: Mechanisms of cancer-induced bone pain. J. Clin. Oncol., 32: 1647-1654.
CrossRefPubMedDirect Link - Khasabova, I.A., C.L. Stucky, C. Harding-Rose, L. Eikmeier and A.J. Beitz et al., 2007. Chemical interactions between fibrosarcoma cancer cells and sensory neurons contribute to cancer pain. J. Neurosci., 27: 10289-10298.
CrossRefDirect Link - Qiu, F., Y. Jiang, H. Zhang, Y. Liu and W. Mi, 2012. Increased expression of tetrodotoxin-resistant sodium channels Nav1.8 and Nav1.9 within dorsal root ganglia in a rat model of bone cancer pain. Neurosci. Lett., 512: 61-66.
CrossRefDirect Link - Hansen, R.R., V. Vacca, T. Pitcher, A.K. Clark and M. Malcangio, 2016. Role of extracellular calcitonin gene-related peptide in spinal cord mechanisms of cancer-induced bone pain. Pain, 157: 666-676.
CrossRefDirect Link - Sun, Y.E., J. Zhang, Y. Lei, C.E. Lu, B. Hou, Z. Ma and X. Gu, 2016. Activation of spinal MrgC-Gi-NR2B-nNOS signaling pathway by Mas oncogene-related gene C receptor agonist bovine adrenal medulla 8-22 attenuates bone cancer pain in mice. Am. J. Transl. Res., 8: 1144-1154.
Direct Link - Lam, D.K., D. Dang, J. Zhang, J.C. Dolan and B.L. Schmidt, 2012. Novel animal models of acute and chronic cancer pain: A pivotal role for PAR2. J. Neurosci., 32: 14178-14183.
CrossRefDirect Link - Zhang, R.X., A. Li, B. Liu, L. Wang and K. Ren et al., 2007. Electroacupuncture attenuates bone cancer pain and inhibits spinal interleukin-1β expression in a rat model. Anesth. Anal., 105: 1482-1488.
CrossRefDirect Link - Poyner, D.R., P.M. Sexton, I. Marshall, D.M. Smith and R. Quirion et al., 2002. International union of pharmacology. XXXII. The mammalian calcitonin gene-related peptides, adrenomedullin, amylin and calcitonin receptors. Pharmacol. Rev., 54: 233-246.
PubMedDirect Link - Ma, W., J.G. Chabot and R. Quirion, 2006. A role for adrenomedullin as a pain-related peptide in the rat. Proc. Natl. Acad. Sci. USA., 103: 16027-16032.
CrossRefDirect Link - Millan, M.J., 1999. The induction of pain: An integrative review. Prog. Neurobiol., 57: 1-164.
CrossRefDirect Link - Hong, Y., Y. Liu, J.G. Chabot, A. Fournier and R. Quirion, 2009. Upregulation of adrenomedullin in the spinal cord and dorsal root ganglia in the early phase of CFA-induced inflammation in rats. Pain, 146: 105-113.
CrossRefDirect Link - Hong, Y., D. Wang, J.G. Chabot, W. Ma, P. Chen and R. Quirion, 2010. A role for protein kinase C-dependent upregulation of adrenomedullin in the development of morphine tolerance in male rats. J. Neurosci., 30: 12508-12516.
CrossRefDirect Link - Wang, D., L. Ruan, Y. Hong, J.G. Chabot and R. Quirion, 2013. Involvement of PKA-dependent upregulation of nNOS-CGRP in adrenomedullin-initiated mechanistic pathway underlying CFA-induced response in rats. Exp. Neurol., 239: 111-119.
CrossRefDirect Link - Zeng, X., M.Y. Lin, D. Wang, Y. Zhang and Y. Hong, 2014. Involvement of adrenomedullin in spinal glial activation following chronic administration of morphine in rats. Eur. J. Pain, 18: 1323-1332.
CrossRefDirect Link - Chen, Y., Y. Zhang, Y. Huo, D. Wang and Y. Hong, 2016. Adrenomedullin mediates tumor necrosis factor-α-induced responses in dorsal root ganglia in rats. Brain Res., 1644: 183-191.
CrossRefDirect Link - Zimmermann, M., 1983. Ethical guidelines for investigations of experimental pain in conscious animals. Pain, 16: 109-110.
CrossRefPubMedDirect Link - Mao-Ying, Q.L., J. Zhao, Z.Q. Dong, J. Wang and J. Yu et al., 2006. A rat model of bone cancer pain induced by intra-tibia inoculation of Walker 256 mammary gland carcinoma cells. Biochem. Biophys. Res. Commun., 345: 1292-1298.
CrossRefDirect Link - Pogatzki, E.M., P.K. Zahn and T.J. Brennan, 2000. Lumbar catheterization of the subarachnoid space with a 32-gauge polyurethane catheter in the rat. Eur. J. Pain, 4: 111-113.
CrossRefDirect Link - Gendron, L., A.L. Lucido, F. Mennicken, D. O'Donnell, J.P. Vincent, T. Stroh and A. Beaudet, 2006. Morphine and pain-related stimuli enhance cell surface availability of somatic δ-opioid receptors in rat dorsal root ganglia. J. Neurosci., 26: 953-962.
CrossRefDirect Link - Medhurst, S.J., K. Walker, M. Bowes, B.L. Kidd and M. Glatt et al., 2002. A rat model of bone cancer pain. Pain, 96: 129-140.
CrossRefDirect Link - Hay, D.L., A.C. Conner, S.G. Howitt, D.M. Smith and D.R. Poyner, 2004. The pharmacology of adrenomedullin receptors and their relationship to CGRP receptors. J. Mol. Neurosci., 22: 105-113.
CrossRefDirect Link - Honore, P., S.D. Rogers, M.J. Schwei, J.L. Salak-Johnson and N.M. Luger et al., 2000. Murine models of inflammatory, neuropathic and cancer pain each generates a unique set of neurochemical changes in the spinal cord and sensory neurons. Neuroscience, 98: 585-598.
CrossRefDirect Link - Yang, Y., J. Zhang, Y. Liu, Y. Zheng and J. Bo et al., 2016. Role of nitric oxide synthase in the development of bone cancer pain and effect of L‑NMMA. Mol. Med. Rep., 13: 1220-1226.
CrossRefDirect Link