
Jin Wang
Department of Neurology, Tianjin Nankai Hospital, 300100 Tianjin, China
LiveDNA: 86.5799
Ying Guo
Department of Neurology, Tianjin Nankai Hospital, 300100 Tianjin, China
Shao Yue Zhang
Department of Neurology, Tianjin Nankai Hospital, 300100 Tianjin, China
International Journal of Pharmacology
Year: 2018 | Volume: 14 | Issue: 4 | Page No.: 488-494







ABSTRACT
Background and Objective: Vanillic Acid (VA) is a dihydroxybenzoic acid derivative found in in the roots of Angelica sinensis and used for treating various ailments. The current preclinical study was designed to demonstrate the neuroprotective activity of VA against cerebral Ischemia-Reperfusion (I/R) injured rat model. Materials and Methods: Healthy Sprague-Dawley (SD) rats (n = 40) were segregated into 4 groups. Rats received only saline (Group I), rats were induced by Middle Cerebral Artery Occlusion (MCAO) for 90 min and followed by reperfusion for 24 h (group II), group III and IV rats pretreated with VA (50 or 100 mg kg–1) for 14 days and followed by MCAO induction. Data were analyzed using Turkey’s test with SPSS software. Results: A considerable decline in the neurological deficit score and cerebral infarct area was observed in VA pretreated group. On administration with VA (50 or 100 mg kg–1) concomitantly lowered the levels of lipid peroxidation product (Malondialdehyde-MDA) with improved antioxidant status (superoxide dismutase, catalase). Whereas, the inflammatory markers such as interleukins-6 (IL-6), IL-1β, Tumor Necrosis Factor Alpha (TNF-α) and Nuclear Factor Kappa B (NF-κB) p65 subunit were remarkably decreased upon VA supplementation, on comparison with MCAO induced group. Furthermore, the relative protein expression of TNF-α and NF-κB p65 were significantly hampered on 14-day intervention with VA. Conclusion: The outcome of this study inferred that VA (100) could exert better neuroprotective activity by improving neuronal function via attenuating inflammatory cascade.
PDF Abstract XML References Citation
Received: May 21, 2017;
Accepted: August 17, 2017;
Published: April 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Jin Wang, Ying Guo and Shao Yue Zhang, 2018. Vanillic Acid Improve Neural Function after Focal Cerebral Ischemia-reperfusion Rats. International Journal of Pharmacology, 14: 488-494.
DOI: 10.3923/ijp.2018.488.494
URL: https://scialert.net/abstract/?doi=ijp.2018.488.494
DOI: 10.3923/ijp.2018.488.494
URL: https://scialert.net/abstract/?doi=ijp.2018.488.494
INTRODUCTION
Ischemic stroke is one of the deadliest conditions owing to thromboembolic occlusion of middle or central cerebral artery1. Studies have shown that stroke claims more lives than any other neurological disease due to serious physical deficits2,3. Abundant evidence has confirmed that the strong connection exists between oxidative stress and inflammatory response in several neurological diseases especially in IR induced strokes as it provokes secondary brain damage4,5. At present only recombinant tissue Plasminogen Activator (rtPA) is the acceptable therapeutic strategy for treating stroke. However, the usage of rtPA is much limited due to the risk of many contraindications and intracerebral hemorrhage6. Therefore, the need of an effective treatment to combat stroke related abnormalities is in greater demand.
Vanillic Acid (VA) is a dihydroxybenzoic acid derivative found in higher quantity in the roots of Angelica sinensis, the bark of Amburana cearensis and fruits of Euterpe oleracea5. Vanillic acid is an oxidative product of vanillin produced during the bioconversion of ferulic acid to vanillin. Vanillin is commonly used flavoring agent in food industry owing to its pleasant and creamy odor7. Experimental studies have highlighted its various biological properties such as antioxidant, anti-inflammatory and anti-tumor activities8,9. Also, VA can act as hepatoprotective, gastroprotective and nephroprotective agent10-12. Furthermore, vanillic acid is shown to exhibit cardioprotective activity against cardiac ischemia-reperfusion model13.
Previously, vanillic acid has been reported to improve cognitive function (neuroprotective activity) against streptozotocin-induced in mice model14. Recently, vanillic acid is proved to attenuate Aβ1-42 induced oxidative stress and cognitive impairment in Alzheimer’s Disease (AD) mice model15. Nevertheless, no studies have been conducted with vanillic acid against the cerebral ischemia-reperfusion model. Hence, the current study was framed to assess the neuro-therapeutic effect of vanillic acid followed by MCAO induced focal cerebral ischemic reperfusion injury in a rat model.
MATERIALS AND METHODS
Chemicals and reagents: Vanillic acid, hydrogen peroxide, tween-20, phosphate buffered saline, 2, 3, 5-triphenyl tetrazolium chloride (TTC), Sodium Dodecyl Sulfate (SDS) and pentobarbital sodium were purchased from Sigma-Aldrich (MO, USA). Remaining chemicals and reagents used for this current study are of analytical grade.
Experimental rats: Forty healthy male Sprague-Dawley (SD) rats weighing 260-280 g were purchased from animal center of Tianjin Nankai Hospital University. Rats were maintained at 22-24°C on a 12 h day/light cycle in a metabolic steel cage with ad libitum (free access to water and food). All the experimental procedures employed in the current experiment was approved by the animal ethical board committee members of Tianjin Nankai Hospital University (TNHU-2402/16) by following the guidelines put forth by health guide for handling and care of laboratory animals.
Procedure for MCAO/IR insult: Middle Cerebral Artery Occlusion (MCAO) or focal IR insult was carried out by the method of Longa et al.16. Sham-operated control rats underwent midline neck incision but no ligation or occlusion and reperfusion process. The body temperature was maintained at 37°C by heating lamp/warm pad. Finally, the wound was then sutured and disinfected and allowed to completely recover from anesthesia and moved back to their respective cages.
Experimental grouping: This pre-clinical study was conducted at Tianjin Nankai Hospital University from Sep., 2016 to Nov., 2016. Group I rats received saline via i.p and served as sham-operated group, group II rats were induced by MCAO for 90 min and followed by reperfusion for 24 h and served as IR or vehicle group (Veh). Group III and IV rats pretreated orally with 50 and 100 mg kg–1 of VA (via oral gavage) for 14 days and followed by IR induction and served as VA 50+Veh group and VA 50+Veh group respectively.
Sample preparation: Rats were euthanized by i.p pentobarbital sodium injection after 24 h of reperfusion and brains samples were excised immediately and stored in -80°C. The cerebral cortex region was isolated and homogenized using lysis phosphate buffer and centrifuged at 3000 rpm for 10 min to obtain a supernatant. This supernatant was used for biochemical and molecular analysis.
Neurological deficit score: The neurological deficit score was evaluated after 24 h of reperfusion by an examiner blinded to all experimental groups by following Longa’s score scaling system16.
Cerebral infarct volume/size: The cerebral hemispheres were excised carefully and sliced into 2 mm coronal slices via a frontal pole and stained with 2% TTC solution for 20 min at 37°C and rinsed with phosphate buffered saline to remove the excessive stain. Then, the sections were fixed with 10% paraformaldehyde for 8-10 h. The coronal slices were placed in an Image scanner and the infarct volume (pale white) was quantified using Image J software (1.3 V; Image J, MD, USA). The infarct volume was quantified (Tureyen et al.17 method) by using the Eq. 1:
| (1) |
Antioxidants and lipid peroxidation products: The activities of superoxide dismutase (SOD), catalase (CAT) and the levels of lipid peroxidation products-malondialdehyde (MDA) in cerebral tissue were assessed using commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
Inflammatory markers: Inflammatory markers like IL-1β, IL-6, TNF-α in cerebral homogenate were measured by commercial ELISA kits provided by Thermo Fisher Scientific (MA, USA) in according to the supplier’s protocol. Whereas, nuclear factor NF-κB p65 subunits were evaluated in a nuclear fraction of cerebral homogenate using nuclear/cytosolic fractionation kit from Bio-vision (CA, USA) and the concentration of NF-κB p65 was determined by ActivELISA kit from Imgenex Corporation (CA, USA).
Western blot: The protein contents in the cerebral (Cortex) homogenate were determined by BCA assay kit from Beyotime, Biotechnology, Jiangsu, China with manufacturers’ instruction. Protein quantity of 40 μg per lane (both nuclear and cytosolic fraction) were loaded into each well of 12% sodium dodecyl sulfate (SDS)-polyacrylamide gels and electro-transferred to polyvinylidene fluoride (PVDF) membrane using the SDS-PAGE technique. The membranes were blocked with 5% skimmed milk and tween 20 with Tris-Buffered Saline (TBS) and then incubated with primary antibodies (shaker) for overnight at 4°C. The primary antibodies such as mouse polyclonal anti-NF p65, anti-TNF-α (1:1000, 1:1200; Abcam, Cambridge, UK), mouse anti-rat β-actin, anti-rabbit-histone H3 (1:800, 1:1000; Abcam, Cambridge, UK) were used. Both β-actin and histone H3 act as an internal control. The TBS was used to remove unbound antibodies, then membranes were probed with anti-mouse secondary antibodies conjugated to horseradish peroxidase (HRP) (1:1500 and 1:2000, respectively; Promega, WI, USA) in TBS at room temperature for 1 h and washed again by TBS. The absorbance (OD) was quantified using an enhanced chemiluminescent system (Thermo Fisher Scientific, MA, USA) and the signals were quantified using ImageJ software (MD, USA).
Statistical analysis: Values are expressed as the Mean±Standard Deviation (SD). The difference between each experimental group was quantified with one-way ANOVA followed by the Turkey’s test with SPSS software (Ver:21). The p<0.05 was considered as statistically significant5.
RESULTS
Effects of VA on neurological deficit score and cerebral infarct volume: The I/R induced cerebral injury was assessed by evaluating the neurological deficit score and infarct volume. Figure 1a and b show the efficacy of VA on neurological deficit score and cerebral infarct volume in experimental rats. The neurological deficit score and cerebral infarct volume were significantly increased (p<0.01) in Veh group (IR-group) as compared to sham-operated control rats. Pretreatment with VA 50 (p<0.05) and 100 (p<0.01) for 14 days could be considerably reduce the levels of both neurological deficit score and cerebral infarct volume.
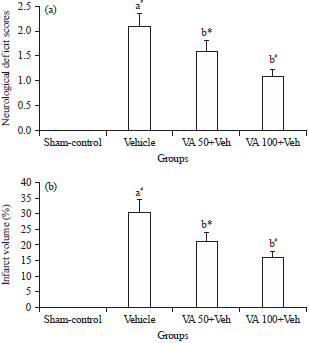 | |
| Fig. 1(a-b): | Effects of VA on (a) Neurological deficit score and (b) Cerebral infarct volume in experimental rats |
Data were expressed as the Mean±Standard Deviation (SD). Significant difference: *p<0.05, #p<0.01, aComparison with sham-control group and bComparison with vehicle (Veh) | |
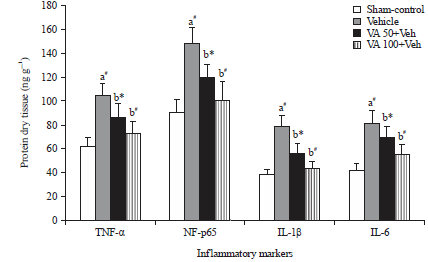 | |
| Fig. 2: | Effects of VA on inflammatory markers in experimental rats |
Data were expressed as the Mean±Standard Deviation (SD). Significant difference: *p<0.05, #p<0.01, aComparison with sham-control group, bComparison with vehicle (Veh) | |
| Table 1: | Effect of VA on the levels of lipid peroxidation products and antioxidant status in experimental rats |
 | |
Data were expressed as the Mean±Standard Deviation (SD). Significant difference: *p<0.05, #p<0.01, a: Comparison with sham-control group, b: Comparison with vehicle (Veh). One unit of SOD represent the amount of enzyme required to inhibit 50% of O2 production at 560 nm. One unit of CAT represent the amount that consumed to inhibit 1 μmol of H2O2, at 540 nm. MDA: Malondialdehyde (lipid peroxidation product), CAT: Catalase, SOD: Superoxide dismutase | |
Effect of VA on antioxidant status and lipid peroxidation products: The activities of cerebral antioxidants and lipid peroxidation products in experimental rats are epitomized in Table 1. The activities of SOD and CAT were substantially attenuated (p<0.01) with increased MDA levels in IR-induced rats (Veh). Meanwhile, supplementation with VA (50 and 100 mg kg–1) could markedly elevate the activities of SOD and CAT, with significantly lowered (p<0.01) MDA levels on comparison with Veh group.
Effects of VA on inflammatory markers: Figure 2 shows that a pronounced increase (p<0.01) in the concentration of nuclear factor NF-p65 subunit of NF-κB and cytosolic fraction like TNF-α, IL-1β and IL-6 in the cerebral tissue of IR-induced rats on equivalence with sham-operated control rats (Fig. 2). Administration with VA 50 (p<0.05) and 100 (p<0.01) significantly lowered the concentration of nuclear factor NF-p65 subunit of NF-κB and cytosolic fractions IL-1β, IL-6 and TNF-α than Veh group.
Effect of VA on the protein expressions of TNF-α and NF-p65 subunit: Figure 3 illustrates the effect of VA on the protein expression of TNF-α (3A) and NF-p65 subunit (3B) in cerebral tissue of experimental rats. Both TNF-α and NF-p65 protein expression were exponentially upregulated (p<0.01) in IR-induced group Vs sham-control rats. Whereas, treatment with VA 50 (p<0.05) and 100 (p<0.01) markedly down-regulated the protein expression of TNF-α and NF-p65 subunit on comparison with IR/Veh group.
DISCUSSION
The results indicate the beneficial/neuroprotective efficacy of VA (50 and 100 mg kg–1) against cerebral Ischemia-Reperfusion (I/R) injured rat model via improving the antioxidant status and attenuating inflammatory response. For this preclinical study, a common ischemic stroke MCAO/R model was preferred as it resembles most of the pathophysiological features of stroke18,19. To evaluate the neuroprotective activity of VA, we determined the neurological deficit score and infarct volume after MCAO/R induction. The neurological deficit score and cerebral infarct volume were significantly increased (p<0.01) in IR-induced/veh group due to MCAO/reperfusion-induced brain damage via excess free radical generation. These results are in corroboration with the studies of Pramila et al.20.
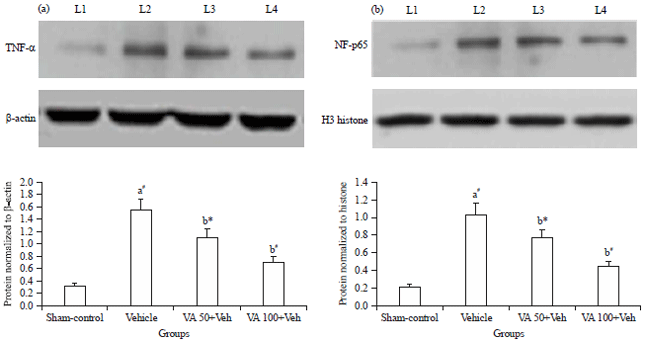 | |
| Fig. 3(a-b): | Effects of VA on cerebral protein expressions of (a) Tumor necrosis factor alpha (TNF-α) and (b) Nuclear factor κB p65 subunit (NF-κB p65) in experimental rats |
Data were expressed as the Mean±Standard Deviation (SD). Significant difference: *p<0.05, #p<0.01, aComparison with sham-control group and bComparison with vehicle (Veh), L1: Sham-control group, L2: Vehicle (IR-induced) group, L3: VA 50+Vehicle group, L4: VA 100+Vehicle group | |
Pretreatment with VA 50 and 100 for 14 days, would considerably lower the neurological deficit score and cerebral infarct volume is owing to antioxidant activity. Vanillic acid shows excellent free radical scavenging activity because of its two-free hydroxyl groups21. Moreover, Prince et al.22 indicated that supplementation with vanillic acid could substantially lower the infarct size/volume of myocardial tissue in isoproterenol-induced infarcted rats. In support of the mentioned finding, we would infer that VA (especially 100) showed better neuroprotective activity by improving motor activity and neural damage.
Ample amount of studies has demonstrated that the strong connection exists between oxidative stress and inflammatory response in several neurological diseases especially in IR-Induced strokes as it provokes secondary brain damage4,5. During MCAO/R induction the microglia/macrophages were activated and trigger the activity of NADPH oxidase and thus results in an excessive free radical generation (Oxidative stress). That excessive free radical in turn stimulate various inflammatory cytokines and thereby elicit inflammatory cascade with neuronal cell death23. Hence, the levels of antioxidants were considerably abolished (p<0.01) with elevated lipid peroxidation product production in IR group or Veh group. Whereas, supplementation with VA (50 and 100 mg kg–1) could markedly improve the antioxidant activities of SOD and CAT and thereby notably abrogated the production of lipid peroxidation products like MDA. Similar impression was also highlighted by the study conducted by Dianat et al.13, pointed out that vanillic acid treatment could significantly enhance (p<0.05) the activities of SOD and CAT and thereby effectively lowered the levels of MDA on cardiac ischemic-reperfusion rat model. As indicated earlier vanillic acid also quench free radical generation due to its two-free hydroxyl group.
As previously mentioned, that during the initial phase of IR-induced stroke various inflammatory markers (pro-inflammatory cytokines) were upregulated via NF-κB signaling pathway owing to excessive oxidative stress. Therefore, inhibition of NF-κB signaling pathway (inactivation of microglial cells) might be a potential therapeutic strategy for the treatment of IR-induced stroke24. A pronounced increase (p<0.01) in the concentration of various inflammatory markers in Veh group was due to microglial cell activation/oxidative stress. The VA pretreated rats (50 and 100 mg kg–1) could substantially abolish those inflammatory markers by effectively quenching excessive free radicals as well as downregulating the expression of NF-κB related downstream inflammatory genes. In addition, VA treatment would halt the activation of NF-κB and thereby inhibit the translocation of p65 from cytosol to nucleus. Also, vanillic acid has covalently modified sulfhydryl group and o-methoxy group which might inhibit the activation of an NF-κB signaling pathway in a rat model12.
For better understanding, the mechanism behind the anti-inflammatory activity of VA, pertaining to NF-κB signaling pathway were evaluated by assessing the protein expression of cytosolic TNF-α and nuclear NF-κB p65 subunit in cerebral tissue homogenate. The protein expression of cytosolic TNF-α and nuclear NF-κB p65 were dramatically upregulated (p<0.01) in vehicle group due to excessive inflammatory response. Nevertheless, VA administered rats displayed marked down regulation in the levels of protein expression of cytosolic TNF-α and nuclear NF-κB p65 due to anti-inflammatory activity. These outcomes are in conformity with the data of biochemical analysis (ELISA) of inflammatory markers.
Previously, vanillic acid is reported to inhibit NF-κB p65 subunit expression as well as pro-inflammatory cytokines (TNF-α) in LPS induced model7. Another experimental study also highlighted that administration of vanillic acid could significantly attenuate (p<0.01) the neuroinflammation by downregulating NF-κB p65 subunit in Aβ1-42 induced AD model15. Available evidence suggests that VA can downregulate the downstream pro-inflammatory cytokines gene like TNF-α and IL-1β and IL-6 via inhibiting the activation of NF-κB (p65) signaling pathway and thereby attenuating inflammatory response in IR-induced rats. Few limitations are skipping off apoptotic factors as well as standard drugs for comparison.
CONCLUSION
This preclinical study demonstrates that VA (50 and 100 mg kg–1) could improve neurological motor ability through abolishing neuronal damage and thus reducing the cerebral infarct by improving the antioxidant status and attenuating inflammatory response or cascade. Moreover, VA (50 and 100 mg kg–1) could modulate the expression of various inflammatory factors related to NF-κB signaling pathway. However, 100 mg kg–1 of VA showed better neuroprotective activity as compared with 50 mg kg–1 of VA. Furthermore, studies are required to evaluate the detailed mechanism underlying the neuroprotective activity of VA.
SIGNIFICANCE STATEMENTS
This pre-clinical study discovers the novel neuroprotective agent-vanillic acid that can be beneficial for improving neuronal function against the cerebral ischemia-reperfusion model. This study would help the researchers to discover the pivotal area of ischemic stroke that many researchers were not able to explore. Thus, a new theory on treating cerebral ischemia-stroke vanillic acid (novel neuroprotective agent) with few standard drugs may be effective in combating stroke related complications.
ACKNOWLEDGMENTS
Authors would like to thank all the ethical committee members for approving this study. Also thanks to Tianjin Nankai Hospital University for supporting this project (TNH-5798).
REFERENCES
- Hong, J.H., J.Y. Kim, S.E. Baek, P. Ingkasupart, H.J. Park and S.G. Kang, 2015. Effects of rice bran extracts fermented with lactobacillus plantarum on neuroprotection and cognitive improvement in a rat model of ischemic brain injury. Biomed. Sci. Lett., 21: 92-102.
CrossRefDirect Link - Kim, J., D.Y.W. Fann, R.C.S. Seet, D.G. Jo, M.P. Mattson and T.V. Arumugam, 2016. Phytochemicals in ischemic stroke. Neuro Mol. Med., 18: 283-305.
CrossRefDirect Link - Feigin, V.L., M.H. Forouzanfar, R. Krishnamurthi, G.A. Mensah and M. Connor et al., 2014. Global and regional burden of stroke during 1990-2010: Findings from the Global Burden of Disease Study 2010. Lancet, 383: 245-255.
CrossRefDirect Link - Guan, J., X. Wei, S. Qu, T. Lv, Q. Fu and Y. Yuan, 2017. Osthole prevents cerebral ischemia/reperfusion injury via Notch signaling pathway. Biochem. Cell Biol., (In Press).
CrossRefDirect Link - Calixto-Campos, C., T.T. Carvalho, M.S.N. Hohmann, F.A. Pinho-Ribeiro and V. Fattori et al., 2015. Vanillic acid inhibits inflammatory pain by inhibiting neutrophil recruitment, oxidative stress, cytokine production and NFκB activation in mice. J. Nat. Prod., 78: 1799-1808.
CrossRefDirect Link - Cao, G., X. Ye, Y. Xu, M. Yin, H. Chen, J. Kou and B. Yu, 2016. YiQiFuMai powder injection ameliorates blood-brain barrier dysfunction and brain edema after focal cerebral ischemia-reperfusion injury in mice. Drug Des. Dev. Ther., 10: 315-325.
CrossRefDirect Link - Kim, M.C., S.J. Kim, D.S. Kim, Y.D. Jeon and S.J. Park et al., 2011. Vanillic acid inhibits inflammatory mediators by suppressing NF-κB in lipopolysaccharide-stimulated mouse peritoneal macrophages. Immunopharmacol. Immunotoxicol., 33: 525-532.
CrossRefDirect Link - Chang, W.C., J.S.B. Wu, C.W. Chen, P.L. Kuo, H.M. Chien, Y.T. Wang and S.C. Shen, 2015. Protective effect of vanillic acid against hyperinsulinemia, hyperglycemia and hyperlipidemia via alleviating hepatic insulin resistance and inflammation in High-Fat Diet (HFD)-fed rats. Nutrients, 7: 9946-9959.
CrossRefPubMedDirect Link - Liang, J.A., S.L. Wu, H.Y. Lo, C.Y. Hsiang and T.Y. Ho, 2009. Vanillin inhibits matrix metalloproteinase-9 expression through down-regulation of nuclear factor-κB signaling pathway in human hepatocellular carcinoma cells. Mol. Pharmacol., 75: 151-157.
CrossRefDirect Link - Itoh, A., K. Isoda, M. Kondoh, M. Kawase and A. Watari et al., 2010. Hepatoprotective effect of syringic acid and vanillic acid on CCl4-induced liver injury. Biol. Pharm. Bull., 33: 983-987.
CrossRefDirect Link - Kim, S.J., M.C. Kim, J.Y. Um and S.H. Hong, 2010. The beneficial effect of vanillic acid on ulcerative colitis. Molecules, 15: 7208-7217.
CrossRefDirect Link - Sindhu, G., E. Nishanthi and R. Sharmila, 2015. Nephroprotective effect of vanillic acid against cisplatin induced nephrotoxicity in wistar rats: A biochemical and molecular study. Environ. Toxicol. Pharmacol., 39: 392-404.
CrossRefDirect Link - Dianat, M., G.R. Hamzavi, M. Badavi and A. Samarbafzadeh, 2014. Effects of losartan and vanillic acid co-administration on ischemia-reperfusion-induced oxidative stress in isolated rat heart. Iranian Red Crescent Med. J., Vol. 16.
CrossRefDirect Link - Singh, J.C.H., R.M. Kakalij, R.P. Kshirsagar, B.H. Kumar, S.S.B. Komakula and P.V. Diwan, 2015. Cognitive effects of vanillic acid against streptozotocin-induced neurodegeneration in mice. Pharm. Biol., 53: 630-636.
CrossRefDirect Link - Amin, F.U., S.A. Shah and M.O. Kim, 2017. Vanillic acid attenuates Aβ1-42-induced oxidative stress and cognitive impairment in mice. Sci. Rep., Vol. 7.
CrossRefDirect Link - Longa, E.Z., P.R. Weinstein, S. Carlson and R. Cummins, 1989. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke, 20: 84-91.
CrossRefDirect Link - Tureyen, K., R. Vemuganti, K.A. Sailor and R.J. Dempsey, 2004. Infarct volume quantification in mouse focal cerebral ischemia: A comparison of triphenyltetrazolium chloride and cresyl violet staining techniques. J. Neurosci. Methods, 139: 203-207.
CrossRefPubMedDirect Link - Durukan, A. and T. Tatlisumak, 2007. Acute ischemic stroke: Overview of major experimental rodent models, pathophysiology and therapy of focal cerebral ischemia. Pharmacol. Biochem. Behav., 87: 179-197.
CrossRefDirect Link - Yan, J., M. Zheng and D. Zhang, 2014. Chrysophanol liposome preconditioning protects against cerebral ischemia-reperfusion injury by inhibiting oxidative stress and apoptosis in mice. Int. J. Pharmacol., 10: 55-68.
CrossRefDirect Link - Pramila, B., P. Kalaivani, A. Anita and C.S. Babu, 2015. L-NAME combats excitotoxicity and recuperates neurological deficits in MCAO/R rats. Pharmacol. Biochem. Behav., 135: 246-253.
CrossRefDirect Link - Tai, A., T. Sawano and H. Ito, 2012. Antioxidative properties of vanillic acid esters in multiple antioxidant assays. Biosci. Biotechnol. Biochem., 76: 314-318.
CrossRefDirect Link - Prince, P.S.M., K. Dhanasekar and S. Rajakumar, 2011. Preventive effects of vanillic acid on lipids, Bax, Bcl-2 and myocardial infarct size on isoproterenol-induced myocardial infarcted rats: A biochemical and in vitro study. Cardiovasc. Toxicol., 11: 58-66.
CrossRefDirect Link - Davalos, D., J. Grutzendler, G. Yang, J.V. Kim and Y. Zuo et al., 2005. ATP mediates rapid microglial response to local brain injury in vivo. Nat. Neurosci., 8: 752-758.
CrossRefDirect Link - Zhao, S., J. Yin, L. Zhou, F. Yan and Q. He et al., 2016. Hippo/MST1 signaling mediates microglial activation following acute cerebral ischemia-reperfusion injury. Brain Behav. Immun., 55: 236-248.
CrossRefDirect Link