
Ayrat Usmanovich Ziganshin
Kazan State Medical University, Kazan, Russia
LiveDNA: 712.6715
Rafis Rustemovich Kamaliev
Kazan State Medical University, Kazan, Russia
Azat Iskhakovich Gabdrakhmanov
Department of Fundamental and Clinical Pharmacology, Kazan Federal (Volga Region) University, Kazan, Russia
Adel Evgenyevich Khairullin
Department of Biochemistry, Kazan State Medical University, Kazan, Russia
Sergey Nikolaevich Grishin
Department of Medical and Biological Physics, Kazan State Medical University, Kazan, Russia
International Journal of Pharmacology
Year: 2018 | Volume: 14 | Issue: 8 | Page No.: 1198-1202







ABSTRACT
Background and Objective: Inhibitory action of ATP on contractions of frog isolated sartorius muscle was shown earlier, while hydrocortisol prevented that effect of ATP. The aim of this study was to investigate whether ATP also inhibits the contractility of sartorius muscle on stress-induced animals. Materials and Methods: The influence of ATP on contractions and end-plate current (EPC) of sartorius muscles isolated from frogs exposed to electric foot-shock stimulation (EFSS) was analyzed. Results: In the muscles of frogs exposed to EFSS, the inhibitory effect of ATP on contractions was significantly decreased and on EPC was abolished. The level of cortisol was much higher in the blood obtained from EFSS animals comparing with control ones. Conclusion: The EFSS has inhibitory action on the effect of ATP on the frog sartorius muscle and that could be due to increase of cortisol blood level.
PDF Abstract XML References Citation
Received: April 03, 2018;
Accepted: August 14, 2018;
Published: October 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ayrat Usmanovich Ziganshin, Rafis Rustemovich Kamaliev, Azat Iskhakovich Gabdrakhmanov, Adel Evgenyevich Khairullin and Sergey Nikolaevich Grishin, 2018. Foot-shock Stimulation Decreases the Inhibitory Action of ATP on Contractility and End-plate Current of Frog Sartorius Muscle. International Journal of Pharmacology, 14: 1198-1202.
DOI: 10.3923/ijp.2018.1198.1202
URL: https://scialert.net/abstract/?doi=ijp.2018.1198.1202
DOI: 10.3923/ijp.2018.1198.1202
URL: https://scialert.net/abstract/?doi=ijp.2018.1198.1202
INTRODUCTION
There is much evidence now that ATP is a co-transmitter and a neuromodulator in the peripheral and central nervous systems influencing many important body functions. In skeletal muscles, ATP released with acetylcholine (ACh) can modify exocytosis of ACh by negative feedback mechanism1 as well as increase the sensitivity of postsynaptic nicotinic receptors2.
It is well known that steroid hormones do not influence the contractile responses of striated muscles, although they can regulate the release of the neurotransmitters leading to inhibition or facilitation of end-plate current (EPC)3. For example, steroid hormones can lead to calcium influx reduction in C2C12 striated cell muscles4, while estrogen hormones cause ATP-induced Ca2+ flux declining5. The ATP and adenosine significantly inhibited the contractions and reduced the amplitude of end-plate currents in frog isolated sartorius muscle6. Results of the earlier experiments showed that the inhibitory actions of ATP but not adenosine can be prevented by hydrocortisol in vivo and in vitro7. Those experiments lead to hypothesis that corticosteroids might antagonize the effects of ATP by some non-genomic influence.
In order to study whether that glucocorticoid-ATP antagonism can be seen also in ex vivo experiments, this paper investigated the influence of high level of glucocorticoid concentration in blood (induced by the acute stress condition-foot-shock stimulation) on actions of ATP and adenosine on contractions and EPC of frog sartorius muscle.
MATERIALS AND METHODS
General procedures: Experiments were carried out on 48 Rana ridibunda frogs at the temperature of 20-24°C from September until March. Ringer solution composition was (in mM): NaCl 113.0, KCl 2.5, CaCl2 1.8 and NaHCO3 to bring pH up to 7.3-7.4.
Electrical foot-shock stimulation (EFSS): One hour before the experiment, each animal was placed in a separate home cage to wait the exposure to a foot-shock stimulation. That was done in a stimulation cage with 30×30 cm metal floor connected to electric stimulator with 50 cm height transparent plastic walls. A series of three foot-shock stimulation (0.7 mA for 3 sec with a 15 min inter-shock interval) were administered. After the last series, the frog was left in the shock device for 30 min and then returned to a home cage. Frogs were killed 2 h after the last stimulation and used to isolate sartorius muscles. Control animals were also placed into the stimulation cage but no electric foot-shock was provided.
Contractile responses measurement: The isolated muscles were hanged vertically in organ baths filled with the 10 mL of the Ringer solution. An initial tension of 1.0 g was applied to the muscles and 30 min was given to the muscles to equilibrate to the experimental conditions.
Stimulation of the muscles was provided by a pair of platinum rings 2.5 mm in diameter and 15 mm apart through which the muscles were placed. Electrical stimulation applied using a Digitimer Multi Stim D330 module (UK) by rectangular electrical impulses (1 Hz, 0.5 ms, 10 V) and stimulation time was 30 sec. Muscle responses were registered in isometric condition by a force displacement transducer (Linton FSG-01, UK), acquired by a digital data acquisition system (Biopack MP100WSW, USA) and recorded by a personal computer.
Initially electrical stimulation to the muscles was applied every 10 min till responses become stable. These initial responses were taken as initial controls (100%) relative to which all the experimental contractions measured.
The stability of contraction was tested in preliminary experiments where muscles were electrically stimulated 5 times with 10-30 min gaps in-between, during which no drugs or agents were used. Experiments showed that there were no significant changes in muscle contractility during this period.
EPC measurement: A muscle was divided and cut across to eliminate any mechanical responses. Muscle band was fixed with a point to a silicone holder on the lower of a Lucite camera with a 2.5 mL of Ringer solution. The EPC were induced with a frequency of 1/30 Hz with one strong stimulation of the nerve. Synaptic currents were written through the voltage clamp technique by two electrodes with a holding potential of -40 mV. Intracellular electrodes were filled with 2.5 M KCl and were given the resistance of 3-5 MΩ. All drugs were addressed through solution providing tubes with a speed of a 2 mL min–1. Values were normalized according to initial EPC that were assumed to be a 100%.
Cortisone level measurement: Blood was taken by a syringe directly from the frog’s heart. The concentration of cortisol in the serum was measured using LIAISON Kit (Biomedica Gruppe, Austria).
Substances used: Adenosine 5’-triphosphate (ATP) and adenosine were supplied by Sigma.
Analysis of results: Student's one sample two tailed test to compare medians to a control 100% value was used. Student's unpaired test (two tailed) to compare cortisone concentrations was used. Values were calculated as Mean±Standard mean error (n) where n was a muscle sample for mechanical contraction or synapses for EPC.
Ethics: Frogs lived in an aquarium (100 L), partly filled with a water (15°C) that was replaced daily. Animals were killed by beheading, after that the spinal brain was broken down.
The Ethical committee of Kazan State Medical University approved this study (September, 2012 protocol).
RESULTS
Contractile responses: In control experiments ATP and adenosine (both at a concentration of 100 μM) significantly inhibited the contractions of frog sartorius muscles evoked by electrical field stimulation which were 77.6±2.3% (n = 24) and 81.3±2.4% (n = 23) of their initial values, correspondingly (Fig. 1).
In frogs exposed to EFSS inhibitory action of ATP attenuated and was 86.7±6.0% (n = 7), that figure was not significantly different from the initial value. In contrast, after EFSS inhibitory action of adenosine was not significantly affected being 67.5±9.2% (n = 4) still being significantly different from initial value (Fig. 1).
End-plate current (EPC): Motor nerve excitation (0.03 Hz) induced EPC with the amplitude of 199±12 nA (n = 11) on muscles with a holding potential of -40 mV.
In sartorius muscles of control frogs ATP (100 μM) significantly reduced amplitude of EPC in a reversible way to 66.1±1.1% (n = 28) of its initial value. The similar degree of depression (70.4±1.3% (n = 27) was observed after adenosine (100 μM) application (Fig. 2).
In frogs after EFSS, inhibitory action of ATP almost totally abolished amplitude of EPC being 94.1±6% (n = 11) which was statistically identical to initial value. In similar conditions, adenosine (100 μM) decreased the amplitude of EPC to 72.3±5% (n = 6) (Fig. 2).
Cortisol level: The concentration of cortisol in blood serum obtained from foot-shocked frogs was 1.33±0.05 mkg dL–1 (n = 20), while for blood obtained from control frogs, that were not exposed to foot-shock stimulation, it was 0.64±0.06 mkg dL–1 (n = 28) and Student's unpaired test showed a significant difference between these values (p<0.001).
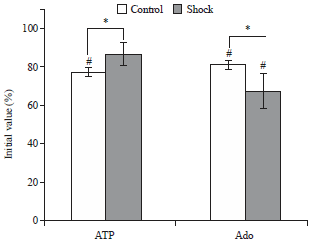 | |
| Fig. 1: | Amplitude of contractions of frog sartorius muscles elicited by electrical field stimulation in the presence of ATP (100 µM) or adenosine (Ado, 100 µM) in control conditions and in frogs exposed to electrical foot-shock stimulation (Shock). Figures are calculated as a percentage to initial values before applications of ATP or adenosine (100%). Values were mean, vertical bars were S.E.M., n = 7-24 (*p<0.05 compared with initial response, #p<0.05 compared with control) |
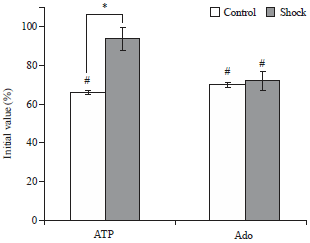 | |
| Fig. 2: | End plate currents of frog sartorius muscles elicited by electrical field stimulation in the presence of ATP (100 µM) or adenosine (Ado, 100 µM) in control conditions and in frogs exposed to electrical foot-shock stimulation (Shock). Figures are calculated initial values (%) before applications of ATP or adenosine (100%). Values are mean, vertical bars are S.E.M., n = 7-24 (*p<0.05 compared with initial response, #p<0.05 compared with control) |
DISCUSSION
This study demonstrated that inhibitory effect of ATP on contractions and EPC of frog sartorius muscles was reduced if frogs were preliminary put on acute stress condition like electric foot-shock. Interestingly, inhibitory actions of adenosine were not affected at similar experimental conditions. Since the increase of cortisol level in the blood of foot-shocked frogs was also revealed, it is suggested that these results are once more support the view about glucocorticoids-ATP antagonism, which has been shown earlier7.
The possible mechanisms of antagonism of glucocorticoids with ATP are not well understood currently. One possibility is involvement in intracellular second messenger production since at the frog neuromuscular junction ATP inhibited the transmitter release via presynaptic P2Y receptors coupled to multiple intracellular cascades8.
Another possible mechanism of ATP-glucocorticoids interactions can be involvement of Ca2+ movements. It is known that at the frog neuromuscular junction ATP inhibits Ca2+ entry into the nerve terminal9 and a similar effect was found for glucocorticoids at C2C12 skeletal muscle cells4. It has also been shown that glucocorticoids inhibit ATP-induced Ca2+ influx in HT4 neuroblastoma cells10 and in endothelial cells11. The involvement of second messengers and changes of cell calcium level is considered currently as the more realistic mechanisms of the cortisol-ATP antagonism.
The interaction of glucocorticoids with ATP might have some physiological and pharmacological significance during inflammation. It is known that ATP released from damaged cells during stress and inflammation has a pro-inflammatory activity through a combination of actions: Release of histamine from mast cells, provoking production of prostaglandins and the production and release of cytokines from immune cells12. It is accepted now that anti-inflammatory action of glucocorticoids is not only due to the well-defined genomic way but also due to some less known non-genomic mechanisms13. It is suggested that antagonism with ATP could be one of such possible non-genomic mechanisms of acute anti-inflammatory action of glucocorticoids.
CONCLUSION
This study verified that foot-shock stimulated frogs have higher blood cortisol levels and ATP, in contrast to adenosine did not considerably modify contractions and EPC of sartorius muscles isolated from these animals.
SIGNIFICANCE STATEMENT
This study showed that ATP, unlike in control conditions, did not considerably inhibit contractions and EPC of sartorius muscles isolated from foot-shock stimulated frogs. This effect is at least partially might be due to higher blood cortisol levels in foot-shock stimulated frogs than in controls.
ACKNOWLEDGMENT
This study was partly supported by the Russian Foundation for Basic Research grant No. 16-04-00101 and was partly performed in the framework of the Russian Government Program of Competitive Growth of Kazan Federal University.
REFERENCES
- Grishin, S.N. and A.U. Ziganshin, 2013. Modulatory role of purines in neuromuscular transmission. Biochem. (Moscow) Suppl. Ser. A: Membr. Cell Biol., 7: 183-191.
CrossRefDirect Link - Grishin, S.N., A.Y. Teplov, A.V. Galkin, A.M. Devyataev and A.L. Zefirov et al., 2006. Different effects of ATP on the contractile activity of mice diaphragmatic and skeletal muscles. Neurochem. Int., 49: 756-763.
CrossRefPubMedDirect Link - Giniatullin, A.R., S.N. Grishin and R.A. Giniatullin, 2000. Effect of hydrocortisone on purine actions in the nerve-muscle preparation. Rossiiskii Fiziologicheskii Zhurnal Imeni IM Sechenova, 86: 1293-1299.
Direct Link - Passaquin, A.C., P. Lhote and U.T. Ruegg, 1998. Calcium influx inhibition by steroids and analogs in C2C12 skeletal muscle cells. Br. J. Pharmacol., 124: 1751-1759.
CrossRefDirect Link - Chaban, V.V., E.A. Mayer, H.S. Ennes and P.E. Micevych, 2003. Estradiol inhibits ATP-induced intracellular calcium concentration increase in dorsal root ganglia neurons. Neuroscience, 118: 941-948.
CrossRefDirect Link - Ziganshin, A.U., R.R. Kamaliev, S.N. Grishin, L.E. Ziganshina, A.L. Zefirov and G. Burnstock, 2005. The influence of hypothermia on P2 receptor-mediated responses of frog skeletal muscle. Eur. J. Pharmacol., 509: 187-193.
CrossRefDirect Link - Ziganshin, A.U., R.R. Kamaliev, S.N. Grishin, B.A. Ziganshin and G. Burnstock, 2009. Interaction of hydrocortisone with ATP and adenosine on nerve-mediated contractions of frog skeletal muscle. Eur. J. Pharmacol., 607: 54-59.
CrossRefDirect Link - Sokolova, E., S. Grishin, A. Shakirzyanova, M. Talantova and R. Giniatullin, 2003. Distinct receptors and different transduction mechanisms for ATP and adenosine at the frog motor nerve endings. Eur. J. Neurosci., 18: 1254-1264.
CrossRefDirect Link - Grishin, S., A. Shakirzyanova, A. Giniatullin, R. Afzalov and R. Giniatullin, 2005. Mechanisms of ATP action on motor nerve terminals at the frog neuromuscular junction. Eur. J. Neurosci., 21: 1271-1279.
CrossRefDirect Link - Han, J.Z., W. Lin and Y.Z. Chen, 2005. Inhibition of ATP-induced calcium influx in HT4 cells by glucocorticoids: Involvement of protein kinase A. Acta Pharmacol. Sin., 26: 199-204.
CrossRefDirect Link - Rogers, K.M., C.A. Bonar, J.L. Estrella and S. Yang, 2002. Inhibitory effect of glucocorticoid on coronary artery endothelial function. Am. J. Physiol.-Heart Circulat. Physiol., 283: H1922-H1928.
CrossRefDirect Link - Di Virgilio, F., P. Chiozzi, S. Falzoni, D. Ferrari, J.M. Sanz, V. Venketaraman and O.R. Baricordi, 1998. Cytolytic P2X purinoceptors. Cell Death Different., 5: 191-199.
CrossRefDirect Link - Goulding, N.J., 2004. The molecular complexity of glucocorticoid actions in inflammation-a four-ring circus. Curr. Opin. Pharmacol., 4: 629-636.
CrossRefDirect Link