
Ming Yang
Department of Osteology, YeDa Hospital, Taishan Road No. 11, Economic and Technological District of Yantai, 264006 Yantai, China
Gaopeng Teng
Department of Pharmacy, YeDa Hospital, 264006 Yantai, China
Guangjian Li
Department of Anesthesiology, YeDa Hospital, 264006 Yantai, China
Tao Huang
School of Pharmacy, Jiangsu University, 212000 Zhenjiang, China
Renguo Xu
Department of Osteology, YeDa Hospital, Taishan Road No. 11, Economic and Technological District of Yantai, 264006 Yantai, China
LiveDNA: 86.24824
International Journal of Pharmacology
Year: 2018 | Volume: 14 | Issue: 8 | Page No.: 1169-1178







ABSTRACT
Background and Objective: Inflammation is a key factor in the pathogenesis of arthritis. In the present study, investigated the potential of osthole, a major natural product, to treat monosodium urate crystal (MSU) induced acute gouty arthritis in rats and explore the mechanisms underlying osthole mediated immunomodulation. Materials and Methods: Rats were oral administrated with osthole at 24 h prior to 3 mg MSU injection, the volume and width of swelling ankle were recorded to evaluate the protection of osthole in vivo. LPS+MSU in Raw 264.7 cells was utilized to assess the effects of osthole on NLRP3 inflammasome in vitro. Results: Osthole could alleviate MSU-induced arthritis through inhibiting the generation of inflammatory factors and NLRP3 inflammasome activation in vivo. Pretreatment with osthole significantly suppressed MSU or LPS induced NF-κB signal along with the transcription of inflammatory factors. In addition, osthole decreased MSU induced oxidative stress and lysosomal damage. Current findings illustrated that osthole significantly suppressed NLRP3 inflammasome in synovial tissue and macrophage cells. The potential mechanism may be based on the attenuation of NF-κB mediated first signal and oxidative stress and lysosome mediated second signal in NLRP3 inflammasome activation. Conclusion: Osthole might be a promising therapeutic agent for alleviating gouty arthritis through inhibiting NLRP3 inflammasome.
PDF Abstract XML References Citation
Received: August 13, 2018;
Accepted: September 05, 2018;
Published: October 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ming Yang, Gaopeng Teng, Guangjian Li, Tao Huang and Renguo Xu, 2018. Effectiveness of Osthole on Uric Acid Crystal-induced Acute Gouty Arthritis Through the Inhibition of NLRP3 Inflammasome. International Journal of Pharmacology, 14: 1169-1178.
DOI: 10.3923/ijp.2018.1169.1178
URL: https://scialert.net/abstract/?doi=ijp.2018.1169.1178
DOI: 10.3923/ijp.2018.1169.1178
URL: https://scialert.net/abstract/?doi=ijp.2018.1169.1178
INTRODUCTION
Gouty arthritis is a common form of inflammatory arthritis due to deposition of monosodium urate crystal (MSU) in and around joints, which is characterized by an acutely red, hot and swollen joint with excruciating pain1. Many researches confirmed that inflammatory response to MSU is mediated principally by macrophages2. Activated macrophages release numerous inflammatory mediators such as TNF-α, IL-1β and IL-6 and inflammatory microenvironment can recruit more inflammatory cells to exacerbate edema and joint injury. IL-1β is a critical inflammatory factor in gouty arthritis. Inflammatory responses is alleviated in MSU-induced mice deficient in IL-1β or the IL-1 receptor3,4. In addition, MSU-induced IL-1β release is mediated by NOD-like receptors containing a PYD 3 (NLRP3) inflammasomes2. Inflammation and pain responses are significantly reduced in MSU-treated NLRP3-deficient mice5. Thus, NLRP3 inflammasome is a potential therapeutic target in MSU-induced arthritis.
Osthole (7-methoxy-8-(3-methyl-2-butenyl)-2H-1-benzopyran-2-one) (FIg. 1a) is a natural coumarin, an active constituent derived from Cnidium monnieri (L.) Cusson, that exhibits wide pharmacological effects, including anti-oxidative6,7, anti-cancer8,9, vascular protection10 and immunomodulatory11 properties. Osthole has also been reported to exert anti-inflammatory property. Osthole inhibits NF-κB signaling and inflammatory mediators to protect kidney failure and carotid artery injury12-14. Notably, osthole reduced NF-κB expression in LPS-stimulated adipocytes may associated with the activation15 of PPARα/γ. Osthole alleviates inflammation by an activated Nrf2-mediated antioxidant pathway in the early stage of focal segmental glomerulosclerosis model16. In addition, osthole has effective anti-inflammatory activity in neutrophils through the suppression17 of PDE4.
However, the effect of osthole on MSU-induced arthritis has not been reported. Considering the anti-inflammatory effect of osthole and the critical role of NLRP3 in arthritis, this present study was designed to explore the effect of osthole on NLRP3 inflammasome in MSU-induced acute gouty arthritis and the possible mechanisms involved.
MATERIALS AND METHODS
Materials: Osthole was purchased from aladdin (Shanghai, China). Dimethyl sulfoxide (DMSO), isopropanol, ethanol and chloroform were purchased from China national medicine group chemical reagent co., LTD. Calycosin, Lipopolysaccharides (LPS, Escherichia coli, L2630, Serotype O111:B4), 3-(4,5-dimethylthiazol-2-yl)2,5-diphenyltetrazolium bromide (MTT), ammonium persulfate, sodium dodecyl sulphate, Triton X-100 and Tris-base were procured from Sigma-Aldrich (St. Louis, Missouri, USA). DMEM, RPMI-1640 and fetal bovine serum (FBS), TRIzol were purchased from Life Technologies (Grand Island, New York, USA). Reduced glutathione (GSH) assay kit (A006-2), Superoxide Dismutase (SOD) assay kit (WST-1 method) (A001-3) and Malondialdehyde (MDA) assay kit (TBA method) (A003-1) were all obtained from the Institute of Biological Engineering of Nanjing Jiancheng (Nanjing, China). Anti-p65 (#8242), anti-phosphate-p65 (#3033), anti-NLRP3 (#15101), anti-ASC (#67824), anti-caspase-1 (#3866), anti-COX-2 (#12282), β-actin (#5174), HRP conjugated Goat anti-Rabbit IgG (#7074) and RIPA cell lysate(#9806) were purchased from Cell Signaling Technology corporation. Other chemical reagents were ordered from Sangon Biotech (Shanghai, China).
Synthesis of monosodium urate (MSU) crystal: The MSU was prepared as described previously with little modification2. About 100 mg of uric acid was dissolved in 20 mL of distilled water with 60 μL of 10 mol/L NaOH to maintain pH between 7.2 and 7.4 at 70°C. The solution was kept overnight in constant shaking and then kept at room temperature for 24 h. The mixture was transferred in 15 mL tube and stored at 4°C for 4 days. Needle-like crystals were recovered and suspended by vortex overnight. The crystals were collected after washing twice with 100% ethanol and once with acetone followed by centrifugation at 3000 rpm for 3 min at 4°C. Finally, The MSU crystals were suspended in sterile endotoxin-free Phosphate Buffered Saline (PBS).
MSU induced ankle arthritis model: The MSU induced ankle arthritis model was performed as described previously and modified a little2. About 150-180 g male Sprague-dawley rats were divided to four experimental groups (n = 8/group), including normal group, MSU-induced arthritis group, 20 mg kg–1 osthole+MSU treatment group and 40 mg kg–1 osthole+MSU treatment group. All animal experiments were performed according to guidelines and with approval of Animal Experimentation Ethics Committee at YeDa Hospital (No. 20170520). About 3 mg MSU crystals suspended into 50 μL endotoxin-free PBS were injected intra-articularly into the ankle joint of the rats anaesthetized with 2.5% isoflurane. Osthole was orally administered at 20 and 40 mg kg–1 concentration per day at the day before the injection of MSU crystals. After 48 h of MSU crystals injection, the rats were sacrificed for evaluation of inflammation parameters. The ankle swelling were recorded using was measured using a paw volume meter (Chengdu Taimen Company, China) and the ankle caliper diameter of each group were measured at 24 h after MSU injected.
Cell viability: Raw 264.7 macrophage cells were seeded into 96-well culture plate with the density of 5×103 per well and were treated with osthole for 24 h. A 100 μL/well MTT (1 mg mL–1, dissolved in PBS solution) was added into the 96-well plates. After 4 h incubation at 37°C, 100 μL DMSO was also added. One hour later, the absorbance was determined using a multiplate reader at a wavelength of 570 nm.
NLRP3 inflammasome activation assay: NLRP3 inflammasome activation assay was conducted as described previously18. Raw 264.7 macrophage cells were seeded into 24-well culture plate. Osthole were added for 18 h followed by LPS 100 ng mL–1 for 3 h to activate NF-κB signaling pathway. Then, 1 mg mL–1 MSU were added for 3 h to activate NLRP3 inflammasome. The supernatant was collected to assay the concentration of IL-1β and IL-18.
Western blot analysis: Raw 264.7 cells and synovial tissue were collected after undergoing different treatments. Lysis buffer containing protease cocktail inhibitors (Roche) was added. The homogenate was centrifuged at 14000 g for 15 min at 4°C and the protein concentration determined using BCA kit. Whole cell lysate (20 μg) was loaded onto 12.5% SDS-PAGE. Electrophoresis was performed using a stacking gel at 80 V for 20 min and a separating gel at 110 V for 70 min. The proteins were transferred to PVDF membranes (Millipore, MA, USA) using an electro-blotting apparatus (Bio-Rad, CA, USA) at 300 mA for 90 min. The membranes were blocked for 1 h in TBST containing 0.1% Tween-20 and 5% dry milk and then incubated overnight with primary antibodies. After washing for 3 times in TBST, membrane was incubated for 2 h with horseradish peroxidase-conjugated secondary antibodies. The optical densities of the antibody-specific bands were analyzed using a Luminescent Image Analyzer (Alpha, USA).
Real-time PCR analysis: Total RNA was isolated from RAW 264.7 macrophage cells after pre-treatment with osthole at 100 or 200 μM for 18 h and then cells were stimulated with 1 mg mL–1 MSU or 100 ng mL–1 LPS for 3 h using the TRIzol reagent according to the manufacturer’s instructions. The 2 μg RNA was reverse transcribed by using the higher capacity cDNA reverse transcription kit (Applied Biosystems, CA, USA) and the mRNA expression was carried out using the DyNAmo SYBR Green 2-step qRT-PCR kit (Finnzymes, F430L). Data collection was performed using ABI PRISM 7000 Sequence Detection System with the SYBR Green PCR Master Mix (Applied Biosystems).The 2‾ΔΔCq method was applied to analyze the relative changes in genes expression. Primer sequences used for quantitative real-time PCR analysis are as follows:
| • | TNF-α-forward CTTCTGTCTACTGAACTTCGGG |
| • | TNF-α-reverse CAGGCTTGTCACTCGAATTTTG |
| • | IL-1β-forward ACGGACCCCAAAAGATGAAG |
| • | IL-1β-reverse TTCTCCACAGCCACAATGAG |
| • | IL-18-forward ATCGGCCTCTATTTGAAGATATGACT |
| • | IL-18-reverse CCTCTAGGCTGGCTATCTTTATACATACT |
| • | IL-6-forward CAAAGCCAGAGTCCTTCAGAG |
| • | IL-6-reverse GTCCTTAGCCACTCCTTCTG |
| • | iNOS-forward GCAAACATCACATTCAGATCCC |
| • | iNOS-reverse TCAGCCTCATGGTAAACACG |
| • | COX2-forward CAAGGGAGTCTGGAACATTG |
| • | COX2-reverse -ACCCAGGTCCTCGCTTATGA |
| • | β-actin-forward ACCTTCTACAATGAGCTGCG |
| • | β-actin -reverse CTGGATGGCTACGTACATGG |
ELISA: ELISA kit (Dakewei, China) was utilized for a quantitative measurement of inflammatory factors, such as IL-1β, IL-18, TNF-α and IL-6, according to the manufacturer’s recommendations.
Determination of oxidative stress status: The macrophages were initially cultured in 6 well plate and pre-treated with 100 and 200 μM osthole and cells were stimulated with 1 mg mL–1 MSU crystals for 24 h. Antioxidant status of RAW 264.7 macrophages was determined by estimating the contents of MDA, SOD, CAT, GPx and GSH.
Determination of lysosomal enzymes: The macrophages were initially cultured in 6 well plate and pretreated with 100 and 200 μM osthole and cells were stimulated with 1 mg mL–1 MSU crystals for 24 h. At the end of incubation, supernatants were collected and cells were washed with PBS and then lysed by adding ice cold lysis buffer. The cell lysate were prepared by centrifugation at 12,000 g for 10 min at 4°C. The cell free supernatant was stored at -80°C until its usage in the assay. The activities of β-Galactosidase, N-acetyl-β-D-Glucosaminidase, Cathepsin D and acid phosphatase were measured according to reference19.
Statistical analysis: The experimental data were expressed as Mean±SD of at least three independent experiments in cell experiments. Statistical analysis was performed using One-way analysis of variance (ANOVA) with the SPSS18.0 statistical software. p-values less than 0.05 were considered statistically significant.
RESULTS
Osthole alleviates MSU-induced gouty arthritis: To evaluate the effect of osthole on MSU-induced gouty arthritis in vivo, the MSU gouty arthritis in SD rats were employed. The structural formula of osthole was given in Fig. 1a. Different concentrations of osthole were administrated into rat on the 1st day, 3 mg MSU was dissolved into PBS and injected into ankle joint. The morphology of ankle at 24 h after MSU injected were exhibited in Fig. 1b. Joint swelling was observed obviously in MSU and osthole pre-treatment suppress the swelling. The ankle swelling were recorded using a paw volume meter (FIg. 1c). The MSU triggered the maximal swelling at 12 h, 20 and 40 mg kg–1 osthole alleviated the volume of swelling. In addition, MSU injection induced a significantly increase in the width of the knee joint. About 30 mg kg–1 osthole decreased MSU-induced swelling (Fig. 1d). In a word, osthole improved MSU-induced gouty arthritis in vivo.
Osthole inhibits MSU-induced NLRP3 inflammasome in gouty arthritis: It has been reported that inflammation is involved in MSU-induced gouty arthritis. Inflammatory factors in serum were measured. As shown in Fig. 2a, 3 mg MSU enhanced IL-1β, IL-18, TNF-α and IL-6 and osthole significantly decreased the contents of IL-1β, IL-18, TNF-α and IL-6. IL-1β and IL-18 is generated by NLRP3 inflammasome, the proteins involved in NLRP3 in synovial tissue were further detected. As described in Fig. 2b, the western blot analysis clearly showed that osthole inhibits NLRP3 inflammasome by inhibiting the MSU-induced NLRP3, ASC, cleaved-caspase-1. Moreover, osthole decreased COX2 expression. Therefore, the protection of osthole in arthritis may be associated with inflammatory inhibition.
Osthole decreased IL-1β and IL-18 secretion in LPS and MSU stimulated Raw264.7 cells: Raw 264.7 macrophagy cells were applied to discover the concrete mechanisms of osthole. The cytotoxicity of osthole to Raw 264.7 cells were measured by MTT assay.
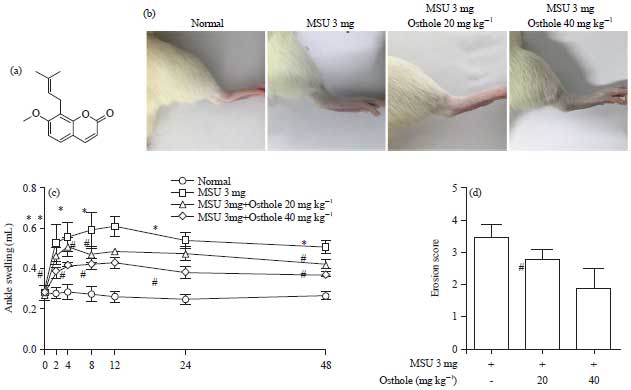 | |
| Fig. 1(a-d): | Osthole alleviates MSU-induced gouty arthritis, (a) Structure of osthole, (b) Ankle joints swelling in each group. Normal was rats without any treatment. MSU group was 3 mg MSU-induced arthritis rats. Osthole groups were oral administration with osthole at a dose of 20 and 40 mg kg–1, respectively, (c) Volume of ankle swelling in each group and (d) Ankle caliper diameter of each group were measured at 24 h after MSU stimulated |
| *p<0.01: Compared with normal group, #p<0.05: Compared with MSU group, values are the Mean±SD | |
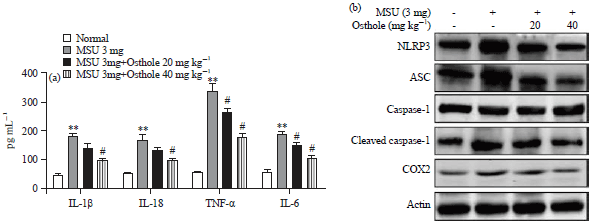 | |
| Fig. 2(a-b): | Effect of osthole on NLRP3 inflammasome in MSU-induced arthritis, (a) Concentrations of IL-1β, IL-18, TNF-α and IL-6 in serum in each group and (b) Rats were oral administrated with 20 and 40 mg kg–1 osthole, the ankle of rat was administrated with 3 mg MSU dissolved into PBS. After 48 h, rats were sacrificed and the expression of NLRP3, ASC, pro-caspase-1, cleaved-caspase-1 and COX-2 in synovial tissue were detected by western blot in different groups. β-actin acted as an internal reference |
| **p<0.01: compared with normal group, #p<0.05: compared with MSU group, values are the Mean±SD | |
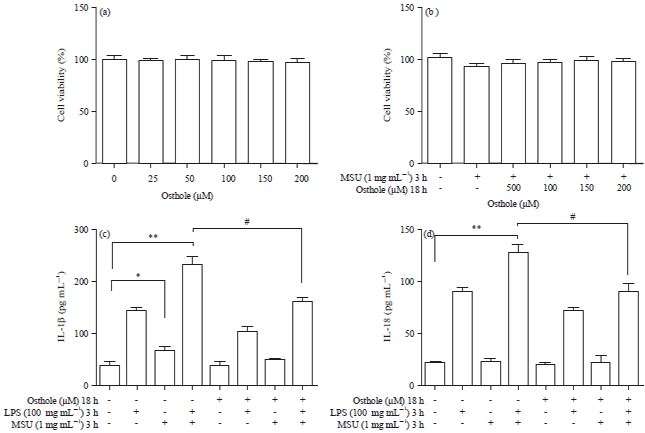 | |
| Fig. 3(a-d): | Osthole decreased IL-1β and IL-18 secretion in LPS and MSU stimulated Raw264.7 cells, (a) Raw 264.7 macrophage cells were treated with different concentrations of osthole for 24 h, MTT assay was performed as described in materials and methods, (b) Raw 264.7 macrophage cells were treated with different concentrations of osthole and MSU (1 mg mL–1) for 24 h. MTT assay was performed and (c-d) Raw 264.7 macrophage cells were treated with 200 µM osthole for 18 h and 100 ng mL–1 LPS for 3 h followed by MSU (1 mg mL–1) for 3 h. Supernatant were collected to measure the level of IL-1β and IL-18 |
| *p<0.05, **p<0.01: compared with control group, #p<0.05: compared with LPS+MSU group, values are the Mean±SD | |
As described in Fig. 3a, osthole treatment for 24 h did not effect cell viability, which indicated that concentration of osthole below 200 μM was not toxic to Raw 264.7. What’s more, cell viability was also not significantly changed by MSU at the concentration of 1 mg mL–1. The combination of MSU and osthole has no effect on cell survival (Fig. 3b). Classic NLRP3 inflammasome model was performed to detect the effect of osthole on MSU stimulated NLRP3 inflammasome. Raw 264.7 cells were treated with osthole (200 μM) for 18 h prior to the stimulation of LPS for 3 h followed by MSU for 3 h. The IL-1β and IL-18 in supernatant were examined. As shown in Fig. 3c and d, IL-1β and IL-18 were significantly increased after LPS and MSU treatment. Osthole can significantly down regulate IL-1β and IL-18 level. The results showed that osthole inhibit LPS and MSU stimulated NLRP3 inflammasome in Raw264.7 macrophage cells.
Osthole decreased inflammatory factors and NF-κB signal in MSU and LPS stimulated Raw264.7 cells: Due to MSU stimulation alone can promote the generation of IL-1β and osthole can suppress MSU stimulated IL-1β,the effect of osthole on NF-κB signal was further investigated. The MSU is not the activator of TLR4, Raw264.7 cells were added MSU for 24 h, the transcription level of IL-1β, IL-18, TNF-α, IL-6, iNOS and COX-2 were detected using real time PCR. As shown in Fig. 4a, MSU alone can promote inflammatory factors transcription and 200 μM osthole can significantly decrease the mRNA level of IL-1β, IL-18, TNF-α, IL-6, iNOS and COX-2. The effect of osthole on LPS stimulated inflammation was further evaluated. Raw 264.7 cells were treated with osthole for 18 h followed by 100 ng mL–1 LPS for 6 h. As depicted in Fig. 4b, LPS treatment produced higher levels of inflammatory factors than MSU treatment alone. osthole can significantly decrease LPS induced inflammatory factors. In addition, the NF-κB signal was further detected. The MSU treatment for 24 h can activate p-p65, an indicator of activated NF-κB signal and osthole down regulated p-p65 (Fig. 4c) as an activator of NF-κB signal, LPS treatment for 6 h can significantly induce p-p65 and osthole can also suppress LPS stimulated p-p65 (Fig. 4d). Thus, present results demonstrated that osthole may suppress NF-κB signal in MSU treatment alone and LPS treatment alone.
Ostholec effectively ameliorates MSU-induced oxidative stress and decreases the release of lysosomal acid hydrolases in RAW 264.7 macrophage cells: In order to assess the effect of osthole on oxidative stress in MSU-stimulated macrophages, cell treated with 100 and 200 μM osthole and MSU (1 mg mL–1) for 24 h. The contents of MDA, SOD, CAT, GPx and GSH were measured.
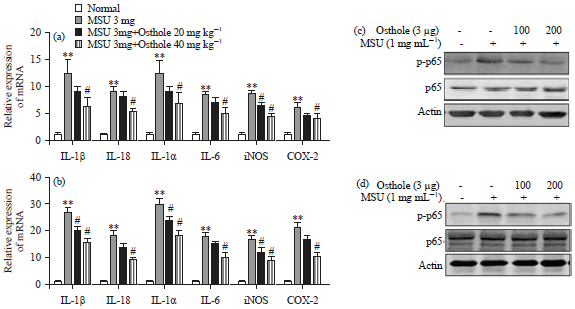 | |
| Fig. 4(a-d): | Osthole decreased inflammatory factors and NF-κB signal in MSU and LPS stimulated Raw 264.7 cells, (a) Raw 264.7 macrophage cells were treated with osthole (100, 200 µM) and MSU (1 mg mL–1) for 24 h. The relative mRNA expression of IL-1β, IL-18, TNF-α, IL-6, iNOS and COX-2 were detected using real time PCR, (b) Raw 264.7 macrophage cells were treated with osthole (100, 200 µM) for 18 h and LPS (100 ng mL–1) for 6 h. The relative mRNA expression of IL-1β, IL-18, TNF-α, IL-6, iNOS and COX-2 were detected using real time PCR, (c) Effect of osthole on the expression of p-p65 and p65 in RAW 264.7 cells induced by 1 mg mL–1 MSU for 24 h. Expression of β-actin acted as an internal reference and (d) Effect of osthole on the expression of p-p65 and p65 in RAW 264.7 cells induced by 100 ng mL–1 LPS for 6 h. Expression of β-actin acted as an internal reference |
| **p<0.01: compared with control group, #p<0.05: compared with MSU or LPS group, values are the Mean±SD | |
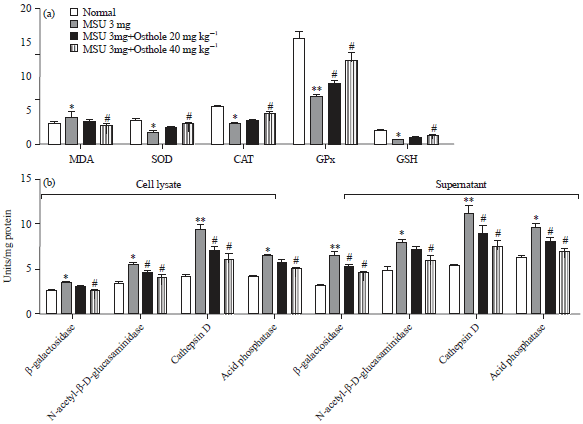 | |
Fig. 5(a-b): | Effect of osthole on MSU-induced oxidative stress and release of lysosomal acid hydrolases in RAW 264.7 macrophage cells, (a) Raw 264.7 macrophage cells were treated with osthole and MSU (1 mg mL–1) for 24 h. The content of MDA, SOD, CAT, GPx and GSH were measured and (b) Raw 264.7 macrophage cells were treated with osthole and MSU (1 mg mL–1) for 24 h, the cellular content and the secreted activities of β-Galactosidase, N-acetyl-β-D-Glucosaminidase, Cathepsin D and Acid phosphatase were further detected |
| *p<0.05, **p<0.01: compared with normal group, #p<0.05: compared with MSU group. Values are the Mean±SD | |
As shown in Fig. 5a, MDA, the byproduct of lipid oxidation, was significantly increased in MSU stimulated Raw 264.7 cells, the antioxidative enzyme including SOD, CAT and GPx were reduced in MSU treated cells. As an endogenous antioxidant, GSH also down regulated in MSU treatment group. Although, 100 μM ostholeh as a tendency to ameliorate oxidative stress, 200 μM osthole significantly decreased MDA content, enhanced the level of SOD, CAT, GPx and GSH against MSU stimulation. To investigate the effect of osthole on lysosomal enzyme activities, cell free supernatant and cell lysate were applied to assess lysosomal enzyme activity. As depicted in Fig. 5b, the increased lysosomal enzyme activities in cell and supernatant were observed in MSU treated Raw 264.7 cells and osthole can significantly decrease the level of lysosomal enzyme activities in cell and supernatant.
DISCUSSION
Nonsteroidal anti-inflammatory drugs (NSAIDs) is the first-line systemic treatment in gouty arthritis. While the side effects of NSAIDs may cause acute renal failure and gastrointestinal bleeding. As part of supplements and alternative medicine, traditional Chinese medicine have also been used to treat gout.
The present results suggest that osthole can alleviate MSU-induced arthritis and improve ankle swelling. The inflammatory factors in serum were decreased and osthole decreased NLRP3, ASC, cleaved caspase-1 and COX-2 expression in synovial tissue. Due to the NLRP3 inflammasome is always activated in macrophage under inflammatory circumstance, the cell viability of osthole in MSU-stimulated Raw264.7 cells were further detected. The classic NLRP3 inflammasome using LPS and MSU were further measured and osthole can decrease NLRP3 mediated IL-1 and IL-18. In order to assess whether osthole effect the first signal or the second signal in NLRP3 inflammasome activation. Inflammatory factors such as IL-1β, IL-18, TNF-α, IL-6, iNOS and COX-2 mRNA and phosphated p65 in first signal were measured under LPS and MSU circumstance respectively. For second signal, oxidative stress markers including MDA, SOD, CAT, GPx and GSH levels were also assessed under MSU stimuli. Considering MSU damaged lysosome, the contents of lysosomal acid hydrolases were further detected. The total enzyme activities between cell lysate and supernatant was increased in MSU stimulated Raw 264.7 and osthole down regulated lysosomal enzymes in cell lysate and supernatant. In a word, osthole can block signal 1 and signal 2 simultaneously in NLRP3 inflammasome to attenuate arthritis.
When the body suffered microbial infection (pathogen associated molecular patterns, PAMPs) or damage signals stimulation (danger-associated molecular patterns, DAMPs), it will start the congenital immunity via pattern recognition receptors on the surface or induction system to promote the type I interferons (IFNα and IFNβ) or inflammatory cytokines such as IL-1β, IL-18 and IL-33 release. The classic NLRP3 inflammasome activation is activated by a combination of two signals. The first signal activates TLR4 (toll like receptor 4) signaling pathway and promotes the generation of precursors such as IL-1 and IL-18 mediated by the nuclear transcription factor (NF-κB). The second signal promotes the assembly of NLRP3/ASC/pro-caspase-1 and pro-caspase-1 was cut into an activated form by itself, the activated caspase-1 cut IL-1 and IL-18 precursors to secret into the extracellular matrix20. The Non-classical NLRP3 inflammasome activation is TLR4-independence. The inflammatory death of the cell, namely pyroptosis, is mediated by caspase-11, caspase-4 or caspase-5 to release and identify pathogenic microorganisms21,22. Thus, the markers of NLRP3 and the activation of NF-κB was measured in vivo and in vitro.
There are some endogenous physiological substances can regulate NLRP3 inflammation. Dopamine23, Omega-3 fatty acids24, β-Hydroxybutyric acid25 and nitric oxide26 inhibited the activation of NLRP3 inflammasome. The serum amyloid A (Aβ)27 uric acid crystallization2, saturated fatty acid28 and cholesterolcrystals29 promotes the NLRP3 inflammasome activation. Mitochondria are the main sources of ROS and are involved in the regulation of NLRP3 inflammasome. However, it's controversial for ROS in NLRP3 inflammasome. On the one hand, it is believed that ROS mediated NLRP3 inflammasome activation30,31. On the other, ROS just promotes NLRP3 prinming32. In addition, MSU crystal-induced arthritis is involved in oxidative stress33. The MSU crystals induce the released Reactive Oxidative Stress (ROS) and reactive nitrogen species (RNS) to alter the cellular oxidative state of the endoplasmic reticulum34. Natural enzyme system exists in the body to remove oxygen free radical, such as superoxide dismutase (SOD), Nitric Oxide Synthase (NOS), catalase (CAT) and glutathione peroxidase (GSH-Px). The SOD is a specific super oxygen anion free radical scavenger, CAT and GSH-Px clear the precursor of oxygen free radicals and lipid peroxide. The SOD indirectly reflect the clearance of oxygen free radicals. As the product of lipid peroxidation, malondialdehyde (MDA) levels can represent lipid peroxide level, which can reflect the generation of free radicals and can also reflect the extent of body injury. Therefore, it detected the response of osthole to oxidative stress.
In addition, the stimulation of silica, alum or β-amyloid, the phagocytic cells are unstable and lysosomal protease B is released into the cytoplasm, which can induce NLRP3 activation. In addition, the crystal or special molecules in endosome may directly damage the lysosome membrane, which lead to engulfed particles diffuse into the cytoplasm and interact with proteins involved in NLRP3 activation35,36. The use of inhibitors of cathepsins can block the activation of NLRP3 inflammasome and the maturity of pro-IL-1β37. Burst release of Ca2+ from lysosome can activate Ca2+-CAMK II-TAK1-JNK pathway, which promotes the ASC oligomerization mediated NLRP3 inflammasome activation38. Thus, lysosomal function was further measured.
Current findings highlight that osthole inhibit NLRP3 inflammasome in arthritis and provide a new mechanism for its anti-inflammation. However, further studies need to be conducted to clarify the detailed inhibition of osthole in lysosomal maturation. These results collectively suggest that osthole can be an effective candidate compound for the treatment of arthritis.
CONCLUSION
This study demonstrated that osthole exerts protective effect on MSU-induced arthritis through the inhibition of NLRP3 inflammasome. This is mainly caused by the inhibition of phosphated NF-κB, the inhibition of oxidative stress and lysosomal protection. Current findings enrich the current knowledge on osthole and osthole can be the potential natural resource to prevent inflammation in MSU-induced arthritis.
SIGNIFICANCE STATEMENT
This study discovers that osthole inhibits MSU-induced inflammation through inhibiting the activation of NLRP3 inflammasome. This study will help the researches uncover the immunoregulation of osthole on arthritis and apply osthole to treat arthritis in future.
ACKNOWLEDGMENT
The authors would like to thank the YeDa Hospital for financing this project.
REFERENCES
- So, A.K. and F. Martinon, 2017. Inflammation in gout: Mechanisms and therapeutic targets. Nat. Rev. Rheumatol., 13: 639-647.
CrossRefDirect Link - Martinon, F., V. Petrilli, A. Mayor, A. Tardivel and J. Tschopp, 2006. Gout-associated uric acid crystals activate the NALP3 inflammasome. Nature, 440: 237-241.
CrossRefPubMedDirect Link - Torres, R., L. Macdonald, S.D. Croll, J. Reinhardt and A. Dore et al., 2009. Hyperalgesia, synovitis and multiple biomarkers of inflammation are suppressed by interleukin 1 inhibition in a novel animal model of gouty arthritis. Ann. Rheum. Dis., 68: 1602-1608.
CrossRefDirect Link - Reber, L.L., T. Marichal, J. Sokolove, P. Starkl and N. Gaudenzio et al., 2014. Contribution of mast cell-derived interleukin‐1β to uric acid crystal-induced acute arthritis in mice. Arthritis Rheumatol., 66: 2881-2891.
CrossRefDirect Link - Amaral, F.A., V.V. Costa, L.D. Tavares, D. Sachs and F.M. Coelho et al., 2012. NLRP3 inflammasome-mediated neutrophil recruitment and hypernociception depend on leukotriene B4 in a murine model of gout. Arthritis Rheumatism, 64: 474-484.
CrossRefDirect Link - Yan, Y.H., S.H. Li, H.Y. Li, Y. Lin and J.X. Yang, 2016. Osthole protects bone marrow-derived neural stem cells from oxidative damage through PI3K/Akt-1 pathway. Neurochem. Res., 42: 398-405.
CrossRefPubMedDirect Link - Zhang, J., J. Xue, H. Wang, Y. Zhang and M. Xie, 2011. Osthole improves alcohol‐induced fatty liver in mice by reduction of hepatic oxidative stress. Phytother. Res., 25: 638-643.
CrossRefDirect Link - Liu, P.Y., D.C. Chang, Y.S. Lo, Y.T. His and C.C. Lin et al., 2018. Osthole induces human nasopharyngeal cancer cells apoptosis through Fas-Fas ligand and mitochondrial pathway. Environ. Toxicol., 33: 446-453.
CrossRefDirect Link - Wu, C., Z. Sun, B. Guo, Y. Ye, X. Han, Y. Qin and S. Liu, 2017. Osthole inhibits bone metastasis of breast cancer. Oncotarget, 8: 58480-58493.
CrossRefPubMedDirect Link - Li, Y., Y. Li, F. Shi, L. Wang, L. Li and D. Yang, 2018. Osthole attenuates right ventricular remodeling via decreased myocardial apoptosis and inflammation in monocrotaline-induced rats. Eur. J. Pharmacol., 818: 525-533.
CrossRefDirect Link - Chiang, C.Y., C.C. Lee, C.K. Fan, H.M. Huang, B.L. Chiang and Y.L. Lee, 2017. Osthole treatment ameliorates Th2-mediated allergic asthma and exerts immunomodulatory effects on dendritic cell maturation and function. Cell. Mol. Immunol., 14: 935-947.
CrossRefDirect Link - Huang, T. and Z. Dong, 2017. Osthole protects against inflammation in a rat model of chronic kidney failure via suppression of nuclear factor-κB, transforming growth factor-β1 and activation of phosphoinositide 3-kinase/protein kinase B/nuclear factor (erythroid-derived 2)-like 2 signaling. Mol. Med. Rep., 16: 4915-4921.
CrossRefDirect Link - Yu, C., P. Li, D. Qi, L. Wang and H.L. Qu et al., 2017. Osthole protects sepsis-induced acute kidney injury via down-regulating NF-κB signal pathway. Oncotarget, 8: 4796-4813.
CrossRefPubMedDirect Link - Li, Y.Q., J.Y. Wang, Z.Q. Qian, Y.L. Li, W.N. Li, Y. Gao and D.L. Yang, 2017. Osthole inhibits intimal hyperplasia by regulating the NF-κB and TGF-β1/Smad2 signalling pathways in the rat carotid artery after balloon injury. Eur. J. Pharmacol., 811: 232-239.
CrossRefDirect Link - Wang, X.L., X. Shang, Y. Cui, X. Zhao, Y. Zhang and M.L. Xie, 2015. Osthole inhibits inflammatory cytokine release through PPAR α/γ-mediated mechanisms in LPS-stimulated 3T3-L1 adipocytes. Immunopharmacol. Immunotoxicol., 37: 185-192.
CrossRefDirect Link - Yang, S.M., Y.L. Chan, K.F. Hua, J.M. Chang and H.L. Chen et al., 2014. Osthole improves an accelerated focal segmental glomerulosclerosis model in the early stage by activating the Nrf2 antioxidant pathway and subsequently inhibiting NF-κB-mediated COX-2 expression and apoptosis. Free Radical Biol. Med., 73: 260-269.
CrossRefDirect Link - Tsai, Y.F., H.P. Yu, P.J. Chung, Y.L. Leu, L.M. Kuo, C.Y. Chen and T.L. Hwang, 2015. Osthol attenuates neutrophilic oxidative stress and hemorrhagic shock-induced lung injury via inhibition of phosphodiesterase 4. Free Radical Biol. Med., 89: 387-400.
CrossRefDirect Link - Heneka, M.T., M.P. Kummer, A. Stutz, A. Delekate and S. Schwartz et al., 2013. NLRP3 is activated in Alzheimer’s disease and contributes to pathology in APP/PS1 mice. Nature, 493: 674-678.
CrossRefDirect Link - Dhanasekar, C., S. Kalaiselvan and M. Rasool, 2015. Morin, a bioflavonoid suppresses monosodium urate crystal-induced inflammatory immune response in RAW 264.7 macrophages through the inhibition of inflammatory mediators, intracellular ROS levels and NF-κB activation. PloS One, Vol. 10, No. 12.
CrossRefDirect Link - Latz, E., T.S. Xiao and A. Stutz, 2013. Activation and regulation of the inflammasomes. Nat. Rev. Immunol., 13: 397-411.
CrossRefDirect Link - Shi, J., Y. Zhao, K. Wang, X. Shi and Y. Wang et al., 2015. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature, 526: 660-665.
CrossRefDirect Link - Vigano, E., C.E. Diamond, R. Spreafico, A. Balachander, R.M. Sobota and A. Mortellaro, 2015. Human caspase-4 and caspase-5 regulate the one-step non-canonical inflammasome activation in monocytes. Nat. Commun., Vol. 6.
CrossRefDirect Link - Yan, Y., W. Jiang, L. Liu, X. Wang, C. Ding, Z. Tian and R. Zhou, 2015. Dopamine controls systemic inflammation through inhibition of NLRP3 inflammasome. Cell, 160: 62-73.
CrossRefDirect Link - Yan, Y., W. Jiang, T. Spinetti, A. Tardivel and R. Castillo et al., 2013. Omega-3 fatty acids prevent inflammation and metabolic disorder through inhibition of NLRP3 inflammasome activation. Immunity, 38: 1154-1163.
CrossRefDirect Link - Youm, Y.H., K.Y. Nguyen, R.W. Grant, E.L. Goldberg and M. Bodogai et al., 2015. The ketone metabolite β-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat. Med., 21: 263-269.
CrossRefDirect Link - Mao, K., S. Chen, M. Chen, Y. Ma and Y. Wang et al., 2013. Nitric oxide suppresses NLRP3 inflammasome activation and protects against LPS-induced septic shock. Cell Res., 23: 201-212.
CrossRefDirect Link - Yu, N., S. Liu, X. Yi, S. Zhang and Y. Ding, 2015. Serum amyloid A induces interleukin‐1β secretion from keratinocytes via the NACHT, LRR and PYD domains‐containing protein 3 inflammasome. Clin. Exp. Immunol., 179: 344-353.
CrossRefDirect Link - Wen, H., D. Gris, Y. Lei, S. Jha and L. Zhang et al., 2011. Fatty acid-induced NLRP3-ASC inflammasome activation interferes with insulin signaling. Nat. Immunol., 12: 408-415.
CrossRefDirect Link - Duewell, P., H. Kono, K.J. Rayner, C.M. Sirois and G. Vladimer et al., 2010. NLRP3 inflammasomes are required for atherogenesis and activated by cholesterol crystals. Nature, 464: 1357-1361.
CrossRefDirect Link - Zhou, R., A.S. Yazdi, P. Menu and J. Tschopp, 2011. A role for mitochondria in NLRP3 inflammasome activation. Nature, 469: 221-225.
CrossRefDirect Link - Nakahira, K., J.A. Haspel, V.A. Rathinam, S.J. Lee and T. Dolinay et al., 2011. Autophagy proteins regulate innate immune responses by inhibiting the release of mitochondrial DNA mediated by the NALP3 inflammasome. Nat. Immunol., 12: 222-230.
CrossRefPubMedDirect Link - Bauernfeind, F., E. Bartok, A. Rieger, L. Franchi, G. Nunez and V. Hornung, 2011. Cutting edge: Reactive oxygen species inhibitors block priming, but not activation, of the NLRP3 inflammasome. J. Immunol., 187: 613-617.
CrossRefPubMedDirect Link - Zamudio-Cuevas, Y., C. Hernandez-Diaz, C. Pineda, A.M. Reginato, J.F. Cerna-Cortes, L. Ventura-Rios and A. Lopez-Reyes, 2015. Molecular basis of oxidative stress in gouty arthropathy. Clin. Rheumatol., 34: 1667-1672.
CrossRefDirect Link - Zamudio-Cuevas, Y., K. Martinez-Flores, J. Fernandez-Torres, Y.A. Loissell-Baltazar and D. Medina-Luna et al., 2016. Monosodium urate crystals induce oxidative stress in human synoviocytes. Arthritis Res. Ther., Vol. 18, No. 1.
CrossRefDirect Link - Lamkanfi, M. and V.M. Dixit, 2012. Inflammasomes and their roles in health and disease. Annu. Rev. Cell Dev. Biol., 28: 137-161.
CrossRefDirect Link - Cassel, S.L., S.C. Eisenbarth, S.S. Iyer, J.J. Sadler and O.R. Colegio et al., 2008. The Nalp3 inflammasome is essential for the development of silicosis. Proc. Natl. Acad. Sci. USA., 105: 9035-9040.
CrossRefDirect Link - Orlowski, G.M., J.D. Colbert, S. Sharma, M. Bogyo, S.A. Robertson and K.L. Rock, 2015. Multiple cathepsins promote pro-IL-1β synthesis and NLRP3-mediated IL-1β activation. J. Immunol., 195: 1685-1697.
CrossRefPubMedDirect Link - Okada, M., A. Matsuzawa, A. Yoshimura and H. Ichijo, 2014. Lysosome rupture-activated TAK1-JNK pathway regulates NLRP3 inflammasome activation. J. Biol. Chem., 289: 32926-32936.
CrossRefPubMedDirect Link