
D. Sivaraman
Centre for Laboratory Animal Technology and Research, Sathyabama University, Jeppiaar Nagar, Rajiv Gandhi Salai, Chennai, No. 51, Selva Vinayagar Koil Street, Perambur, 600119, Tamil Nadu, India
LiveDNA: 91.1377
P. Panneerselvam
Department of Pharmaceutical Chemistry,
P. Muralidharan
Department of Pharmacology and Toxicology, C.L. Baid Metha College of Pharmacy, Jyothi Nagar, Rajiv Gandhi Salai, Thoraipakkam, Chennai, 600097, Tamil Nadu, India
International Journal of Pharmacology
Year: 2016 | Volume: 12 | Issue: 2 | Page No.: 52-65







ABSTRACT
Alzheimer’s disease is a progressive neurodegenerative disorder primarily manifesting as a loss of memory, senile dementia, intra neuronal neurofibrillary tangle formation and cerebral parenchyma deposition of the β-amyloid protein in the form of amyloid plaques is the most stereotypic cognitive and neuropathological hallmarks of AD. The present study was designed to determine the ameliorating effect of hydroalcoholic extract of Ipomoea aquatica (HAEIA) on amyloid beta (Aβ) 25-35-induced cognitive deficits and oxidative stress in mice. Animals were treated with HAEIA for periods of 4 weeks dose-dependently (200 and 400 mg kg–1) then received a single Intra Cerebro Ventricular (ICV) injection of Aβ 25-35 (10 μg/mouse). Cognitive behavioral changes in the mice were evaluated using step down inhibitory avoidance, object recognition test, y-maze, hole board, elevated plus maze and water-maze tests. Brian level neurotransmitter such as dopamine, noradrenaline serotonin, glutamate and metabolic enzymes like acetylcholineserase, monoamine oxidase A and B estimated by using spectrofluorimeter technique. Results obtained from the study indicate that HAEIA at the dose of 400 mg kg–1 significantly ameliorated the cognitive and memory deficits caused by ICV injection of Aβ 25-35. The acetyl cholinesterase, monoamine oxidase and glutamate activity in the brain was lower in HAEIA supplemented groups than the Aβ-injected group. The HAEIA significantly improves the level of serotonin, dopamine, nor-adrenaline in treatment group when compare to Aβ-injected group. In conclusion plant like ipomoea auatica often served has a valuable lead for the clinical management of AD with minimal side effects.
PDF Abstract XML References Citation
How to cite this article
D. Sivaraman, P. Panneerselvam and P. Muralidharan, 2016. Memory and Brain Neurotransmitter Restoring Potential of Hydroalcoholic Extract of Ipomoea aquatica Forsk on Amyloid Beta Aβ (25-35) Induced Cognitive Deficits in Alzheimer’s Mice. International Journal of Pharmacology, 12: 52-65.
DOI: 10.3923/ijp.2016.52.65
URL: https://scialert.net/abstract/?doi=ijp.2016.52.65
DOI: 10.3923/ijp.2016.52.65
URL: https://scialert.net/abstract/?doi=ijp.2016.52.65
INTRODUCTION
Alzheimer’s disease is associated with brain shrinkage and localised loss of neurons, mainly in the hippocampus and basal forebrain. The loss of cholinergic neurons in the hippocampus and frontal cortex is a feature of the disease and is thought to underlie the cognitive deficit and loss of short-term memory that occur in AD.
Two microscopic features are characteristic of the disease, namely extracellular amyloid plaques, consisting of amorphous extracellular deposits of β-amyloid protein (known as Aβ) and intraneuronal neurofibrillary tangles, comprising filaments of a phosphorylated form of a microtubule-associated protein (Tau). Both of these deposits are protein aggregates that result from misfolding of native proteins. They appear also in normal brains, although in smaller numbers. The early appearance of amyloid deposits presages the development of AD, although symptoms may not develop for many years. Altered processing of amyloid protein from its precursor is now recognized as the key to the pathogenesis of AD. This conclusion is based on several lines of evidence, particularly the genetic analysis of certain, relatively rare, types of familial AD, in which mutations of the Amyloid Precursor Protein (APP) gene, or of other genes that control amyloid processing, have been discovered. The APP gene resides on chromosome 21, which is duplicated in Down’s syndrome, in which early AD-like dementia occurs in association with overexpression of Amyloid Precursor Protein (APP) (Shah et al., 2013).
It is well known that the major constituents of senile plaques occurring in AD is amyloid β peptide 1-40 and 1-42 amino acid, which is cleaved from the Amyloid Protein Precursor (APP) (Marata et al., 1988; Kang et al., 1987). The proteases that cut out the Aβ sequence are known as secretases.
The Aβ protein is a potent neurotoxin both in vitro and in vivo. It has been substantiated that the 11 amino acid sequence (25-35) of beta amyloid is neurotoxic for primary neurons (Pereira et al., 1999). The Aβ is secreted by normal cells in cultures and are detected as circulating peptide in the plasma and the cerebrospinal fluid of healthy humans and other mammals (Seubert et al., 1992). Formation of Aβ involves cleavage at two different points, including one in the transmembrane domain of APP by β and γ-secretases. Gamma secretase is a clumsy enzyme that lacks precision and cuts APP at different points in the same victinity, generating Aβ fragments of different lengths including Aβ 40 and 42, mutations favour the formation of Aβ 42. Amyloid hypothesis points to cytotoxicity of mature aggregated amyloid fibrils, which are believed to be the toxic form of protein responsible for disturbing the cells calcium ion homeostasis and thus including apatosis (Yankner et al., 1990). The calcium permeable channels in the membranes resulting in excessive calcium influx and cause the induction of neurotoxic cascades (Mark et al., 1997).
Prevalence rates for dementia increase exponentially with advancing age, ranging from 10% in the age group of 60-65 years to 36% in the age group of 90 years (Sharma et al., 1997). This disease may be presenile or senile onset and the occurrence of disease is before or after the age group of 60 years, but now it is challenging the median age of population. The AD was the 7th leading cause of death in 2004 with 65,829 numbers of deaths (Sloane et al., 2002). It has been estimated that nearly 24 million people with dementia worldwide (Hebert et al., 2003).
The leads of Central Nervous System (CNS) active medicinal plants, which have emerged besides Rawolfia serpentina, Mucuna pruriens for Parkinson’s disease, Ocimum santum as an anti-stress agent, Withania somnifera as anxiolytic, Centella asiatica and Bacopa monneria for learning and memory disorders. Bacopa monneria and Ginkgo biloba for Alzheimer’s disease. The study related to Alzheimer’s Disease (AD) is focused towards the traditionally used rejuvenating and neurotonic agents. The recent trends in the pharmacological studies are based on the biochemical and molecular mechanism which leads to the development of CNS active principles from the herbal drugs (Anekonda and Reddy, 2005; Sheikh et al., 2007; Hussian and Manyam, 1997).
Researches done by FDA and other associations have claimed that herbs contain natural composition of various compounds that affects our internal system (brain, heart and mind). In conventional medicine, those formulations are synthesized by chemical associations and are given in different doses to the person. Around 30% of the ingredients that are added in conventional medicine are extracted from natural resources like plants and herbs (Winston, 2005).
Ipomoea aquatic Forsk is a tender, trailing, vascular semiaquatic plant, found on moist soil along the margin of fresh water, ditches, marches and wet rice fields. It is commercially cultivated as an edible green leafy vegetable in Hong kong, Taiwan, China and also in rural part of India (Edie and Ho, 1969). It is runner type plant with numerous small flowers. Only a very few scientific studies have been conducted on its medicinal aspects. These include oral hypoglycemic (Manvar and Desai, 2013) and anti-oxidant activity (Huang et al., 2005). Treatment on liver diseases (Siddiqui and Husain, 1992), constipation (Samuelsson et al., 1992). Ipomoea aquatic is considered a tonic and it is used to treat gastric and intestinal disorders (Watt, 1972; Westphal, 1993).
Various parts of the Ipomoea are used medicinally in Southeastern Asia and reported to be useful for the treatment of high blood pressure (Perry, 1980) as an emetic in the treatment of opium and arsenic poisoning (Chopra et al., 1956). The juice of this plant has reported to be a purgative and anti-helminthic activity (Datta and Banerjee, 1978) and also as a folklore use this plant is used for the treatment of eye diseases (Jain and Verma, 1981).
From brain free redical quenching: The current study was undertaken to evaluate the memory restoring potential of hydroalcoholic extract of Ipomoea aquatica (HAEIA) against amyloid beta induced cognitive dysfunction in mice by suitable in vivo models pertains to memory and learning task.
MATERIALS AND METHODS
Preparation of the plant extract: The fresh leaf of IA was collected and washed with running water. It was shade dried at room temperature and 1 kg of the dried leaf was made in to coarse powder. The powder was passed through a 60 No mesh sieve. Air dried Powdered drug was extracted with mixture of ethanol:water (6:4) (hydro-alcoholic extract) by using soxhlet extraction. Hydroalcoholic extract of Ipomoea aquatica (HAEIA) was filtered, concentrated by rotary vacuum pump to get the solid mass.
Experimental animal: Healthy swiss albino male mice weighing 20-25 g, obtained from C.L. Baid Metha College of Pharmacy was used for the pharmacological studies. The animals were kept under standard conditions maintained at 25±2°C, 12 h light/dark cycle and given standard pellet diet (Hindustan lever, Bangalore) and water provided ad libitum. The animals were acclimatized to the laboratory conditions for a week prior to the experimentation. All the animals were housed in polypropylene cages using paddy husk bedding. Principles of animal handling were strictly adhered to and the handling of animals was made under the supervision of animal ethics committee of the institute. The experimental protocol was approved by Institutional Animal Ethics Committee (IAEC) of Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA). The C.L. Baid Metha College of Pharmacy, Chennai, Tamil nadu, India.
Intra cerebro ventricular injection of Aβ peptide: The administration of a Aβ 25-35 was performed by identifying the bregma pointing the skull, each mouse was injected at bregma with a 50 μL Hamilton micro syringe fitted with a 26-gauge needle that was inserted to a depth of 2.4 mm. In brief, the needle was inserted unilaterally 1 mm to the right of the middle point equidistant from each eye slightly angled towards 45° 1 mm to the right of the middle point equidistant from the eyes perpendicular to the plane of the skull. Mice exhibited normal behavior within 1 min after injection (Laursen and Belknap, 1986). Animals were divided into six groups.
| Group I | : | Animals injected with phosphate buffered saline (10 µL) |
| Group II | : | Animals injected with Aβ peptide (10 µL) by ICV |
| Group III | : | Animals injected by Aβ peptide (10 µL) by ICV and treated with HAEIA 200 mg kg–1 (p.o.) |
| Group IV | : | Animals injected with Aβ peptide (10 µL) by ICV and treated with HAEIA 400 mg kg–1 (p.o.) |
| Group V | : | Animals injected with Aβ peptide (10 µL) by ICV and treated donepezil 5 mg kg–1 (p.o.) |
Amnesia was induced by Intra Cerebro Ventricular (ICV) injection of Aβ peptide (25-35) preparation to II, III, IV and V groups on the 21st day after pretreatment with HAEIA and continues for the period of 07 days. Control animals were injected with only phosphate buffered saline.
Step down inhibitory avoidance: The apparatus consist of 50×25×25 cm acrylic box, whose floor consisted of parallel 1.0 cm apart. A 7.0 cm wide, 2.5 cm high, 25.0 cm long platform occupied the centre floor. In the training session, immediately after stepping down placing their four paws on the grid the animals will receive a 0.4 MA, 2.0 sec scrambled foot shock. In test session no foot shock will be given and step-down latency is used as a measure of retention (to a ceiling of 300 sec) one-trial step-down inhibitory avoidance in mice involves the activation of two separate memory types, a Short Term Memory (STM) system and a Long Term Memory (LTM) system. Therefore, retention tests is carried out 90 min after training to evaluate STM and 7 days after training to evaluate Long Term Memory (LTM) (Itoh et al., 1990).
Water maze task: The experimental apparatus consisted of a circular water tank (diameter 100 cm, height 35 cm), containing water at 28°C to a depth of 15 cm and rendered opaque by adding titanium dioxide. A platform (diameter 4.5 cm, height 14.5 cm) is submerged 1 cm below the water surface and placed at the midpoint of one quadrant. After several trials, the test is conducted after injection of β amyloid peptide. In each training trial, the time required to escape on to the platform is recorded (Doknark et al., 2014).
Exploratory behavior (hole board): Hole board apparatus is used to predict the locomotor activity of the animals, mice were explored twice, with a 24 h interval, to a 40×40×60 cm open field whose brown linoleum floor was divided into 16 equal squares by white lines. After 1 h of administration of test drug, the animals is placed in the rear left square and left to explore it freely for 5 min during which time the number of lines crossing, head dipping and rearing were counted (Dhingra et al., 2006; Saitoh et al., 2006).
Y-maze task: The Y-maze task is used to measure the spatial working through the spontaneous alternation of behaviour. The maze is made of black painted wood. Each arm is 40 cm long, 3.5 cm high and 10 cm wide and converges at an equal angle. Each mouse is placed at the end of one arm and allowed to move freely through the maze during the training session for 15 min. Mice tend to explore the maze systematically, entering each arm in turn. The ability to alternate requires that the mice know which arm they have already visited. The observations are for 5 min. The series of arm entries, including possible returns into the same arm, are recorded visually. Alternation is defined as the number of successive entries into the three arms, on overlapping triplet sets. The percentage of alternation is calculated as the ratio of actual alternations, defined as the total number of arm entries minus two and multiplied by 100 (Hiramatsu and Inoue, 2000).
Object recognition test: The apparatus comprises of a wooden box (70×60×30 cm) with a grid floor that could be easily cleaned with hydrogen peroxide after each trail. The objects to be discriminated were placed at diagonally opposite corners of the box and were in two different shapes: pyramid of 8 cm side and cylinder of 8 cm height. On day 0, animals were allowed to explore the box without any object for 2 min. On first trail (T1), two identical objects were presented in two opposite corners of the box and the time taken by each mouse to complete 20 sec exploration was measured. Exploration meant directing the nose at a distance less than 2 cm to an object and/or touching with the nose. During the second trail (T2, 90 min after T1), a new object replaced one of the objects present in T1 and mice were left in the box for 5 min. The time spent for exploring New (N) and Familiar (F) objects were recorded separately. Care was taken to avoid place preference and olfactory stimuli by randomly changing the role (F or N) and the position of the two objects during T2 and cleaning them carefully (Yoo et al., 2014).
Elevated plus maze: The apparatus consists of two open arms (35×6 cm) and two enclosed arms (35×6×15 cm). The arm was connected together with a central square of 5×5 cm. The maze is elevated to a height of 100 cm and the procedure is conducted inside a light and sound attenuated room. Mice were placed individually at the end of an open arm of Elevated Plus Maze (EPM) facing away from the central platform and the time it took to move from the end of open arm to either of the closed arms Transfer Latency (TL) was recorded (Bera et al., 2011). Transfer Latency (TL) was taken as the time taken by mouse to move into one of the covered arm with all its four legs was gently pushed into one of the two covered arms and the TL was assigned as 90 sec. The mouse was allowed to explore the maze for 10 sec and then returned to its home cage. Memory retention was examined 24 h after the first day trial. The inflexion ratio was calculated by the equation (Joshi and Parle, 2006a):
where, L0 is the initial Transfer Latency (TL) in sec on first time, L1 is the Transfer Latency (TL) in sec on 2nd time.
Estimation of acetylcholineserase enzyme: Acetylcholineserase (AChE) enzyme activity was estimated by Elman method.
Reagents:
| • | Phosphate buffer (0.1 M), solution A: 5.22 g of K2HPO4 and 4.68 g of NaH2PO4 are dissolved in 150 mL of distilled water, solution B prepared by 6.2 g NaOH is dissolved in 150 mL of distilled water. Solution B is added to solution A to get the desired pH (pH 8.0 or 7.0) and then finally the volume is made up to 300 mL with distilled water |
| • | The DTNB reagent prepared by 39.6 mg of DTNB with 15 mg NaHCO3 is dissolved in 10 mL of 0.1 M phosphate buffer (pH 7.0) |
| • | Acetylthiocholine (ATC) prepared by 21.67 mg of acetylthiocholine is dissolved in 1 mL of distilled water |
Procedure: The mice were decapitated; brains are removed quickly and placed in ice-cold saline. Frontal cortex, hippocampus and septum are quickly dissected out on a petri dish chilled on crushed ice. The tissues are weighed and homogenized in 0.1 M phosphate buffer (pH 8). The 0.4 mL aliquot of the homogenate is added to a cuvette containing 2.6 mL phosphate buffer (0.1 M, pH 8) and 100 μL of DTNB. The contents of the cuvette are mixed thoroughly by bubbling air and absorbance is measured at 412 nm in a spectrophotometer. When absorbance reaches a stable value, it is recorded as the basal reading. Twenty microliter of substrate acetylthiocholine is added and change in absorbance is recorded. Change in the absorbance per minute is thus determined (Joshi and Parle, 2006b; Palchykova et al., 2006; Ellman et al., 1961).
Estimation of monoamine oxidase A and B: The MAO-A and B activity was estimated by the method of Charles and McEwen (Weinstock et al., 2003).
Reagents: Sucrose (0.25 M), tris buffer (0.1 M), EDTA (0.02 M), serotonin (4 mM), sodium phosphate buffer (100 mM, pH 7.4), hydrochloric acid (1 M), butyl acetate/cyclohexane and benzyl amine.
Preparation of sample: Mouse brain mitochondrial fraction are prepared by cutting the brain sample in to small pieces and rinsed in 0.25 M sucrose, 0.1 M tris, 0.02 M EDTA (pH 7.41) to remove blood. The pieces were homogenized for 45 sec in a potter homogenizer with 400 mL of the same medium. The homogenate was centrifuged at 800 rpm for 10 min and the pellets were discarded. The supernatant was then centrifuged at 12,000 rpm for 20 min in the same medium. The precipitate was washed twice more with 100 mL of sucrose tris EDTA and again suspended in 50 mL of the medium.
Procedure for estimation of MAO-A: Two hundred and fifty microliter of the homogenate was added to 250 μL of serotonin and 250 μL of buffer. The reaction tube was placed at 37°C for 20 min and the reaction was arrested by the addition of 200 μL of 1 M HCl. The reaction product was extracted with 5 mL of butyl acetate. The organic phase was separated and measured at 280 nm using a spectrophotometer. Blank samples were prepared by adding 1 M HCl (200 μL) prior to reaction and the reaction was carried out.
MAO-A is expressed in nmoles per milligram protein
Procedure for estimation of MAO-B: Two hundred and fifty microliter of the homogenate was added to 250 μL of benzyl amine and 250 μL of buffer. The reaction tube was placed at 37°C for 20 min and the reaction was arrested by the addition of 200 μL of 1 M HCl. The reaction product was extracted with 5 mL of cyclohexane. The organic phase was separated and measured at 242 nm using a spectrophotometer. Blank samples were prepared by adding 1 M HCl (200 μL) prior to reaction and the reaction was carried out.
The MAO-B activity is expressed in nmol mg–1 protein.
Estimation of dopamine
Preparation of tissue extracts
Reagents:
| • | HCl-Butanol solution: (0.85 mL of 37% hydrochloric acid in 1 L n-butanol) |
| • | Heptane |
| • | 0.1 M HCl: (0.85 mL conc. HCl upto 100 mL H20) |
Procedure: On the day of experiment mice were sacrificed, whole brain was dissected out and the sub cortical region (including the striatum) was separated. Weighed quantity of tissue is homogenized in 5 mL HCl–butanol for about 1 min. The sample was then centrifuged for 10 min at 2000 rpm. An aliquot supernatant phase (1 mL) was removed and added to centrifuge tube containing 2.5 mL heptane and 0.31 mL HCl of 0.1 M. After 10 min of vigorous shaking, the tube was centrifuged under the same conditions as above in order to separate the two phases and the overlaying organic phase was discarded. The aqueous phase (0.2 mL) was then taken either for 5-HT, NA and DA assay. All steps were carried out at 0°C (Schlumpf et al., 1974).
Estimation of noradrenaline
Reagents:
| • | 0.4 M HCl: 3.4 mL conc. HCl up to 100 mL H20 |
| • | Sodium acetate buffer (pH 6.9): 2.88 mL of 1 M acetic acid (5.7 mL of glacial acetic acid upto 100 mL with distilled water) +27.33 mL of 0.3 M sodium acetate (4.08 g of sodium acetate 100 mL with distilled water) and volume is made up to 100 mL with distilled water). The pH is adjusted with sodium hydroxide solution |
| • | 5 M NaOH: 20 g of sodium hydroxide pellets dissolved in distilled water and volume is made up to 100 mL with distilled water) |
| • | 0.1 M Iodine solution (in ethanol): 4 g of pot. Iodide +2.6 g of iodine dissolved in ethanol volume is made up to 100 mL) |
| • | Na2SO3 solution (0.5 g Na2SO3 in 2 mL H2O+18 mL 5 M NaOH) |
| • | 10 M Acetic acid: 57 mL of glacial acetic acid dissolved in distilled water up to 100 mL |
Procedure: To the 0.2 mL of aqueous phase, 0.05 mL 0.4 M HCl and 0.1 mL of EDTA/sodium acetate buffer (pH 6. 9) were added, followed by 0.1 mL iodine solution (0.1 M in ethanol) for oxidation. The reaction was stopped after 2 min by addition of 0.1 mL Na2SO3 solution. The 0.1 mL acetic acid is added after 1.5 min. The solution was then heated to 100°C for 6 min when the sample again reached room temperature, excitation and emission spectra were read from the spectrofluorimeter. The readings were taken at 330-375 nm for dopamine and 395-485 nm for nor-adrenaline (Pal, 2009).
Estimation of serotonin
Reagents: O-phthaldialdehyde (OPT) reagent: (20 mg in 100 mL conc. HCl).
Procedure: To 0.2 mL aqueous extract 0.25 mL of OPT reagent were added. The fluorophore will be developed by heating to 100°C for 10 min. After the samples reached equilibrium with the ambient temperature, readings were taken at 360-470 nm in the spectrofluorimeter. Tissue blanks for Dopamine and nor-adrenaline were prepared by adding the reagents of the oxidation step in reversed order (sodium sulphite before iodine). For serotonin tissue blank, 0.25 mL cont. the HCI without OPT was added. Internal standard: (500 μg mL–1) each of noradrenaline, dopamine and serotonin are prepared in distilled water:HCl-butanol in 1:2 ratio (Balamurugan et al., 2009).
Estimation of glutamate
Reagents:
| • | Glycine-hydrazine buffer |
| • | Adenosine-5'-diphosphate |
| • | Nicotinamide adenine dinucleotide |
| • | Glutamate dehydrogenase |
| • | Perchloric acid |
| • | Phosphate solution |
| • | Deproteinization |
Weighed quantity of brain portion was homogenized with perchloric acid and centrifuge for 10 min at 3000 rpm. Adjust supernatant fluid to pH 9 with phosphate solution. Allow to stand 10 min, in an ice bath and then filter through a small, fluted filter paper. Allow to warm to room temperature, dilute and take 1.0 mL for the assay.
Assay system: Wavelenghth: 340 nm, light path: 1 cm, final volume: 3.35 mL, room temperature. For each series of measurements prepare reagent blank containing water instead of sample.
Pipette successively into cuvettes the following solutions:
| • | Glycine-hydrazine buffer |
| • | Sample |
| • | ADP solution |
| • | NAD solution |
| • | Mix, read extinction E1 at 340 nm |
The add glutamate dehydrogenase solution and mix well, allow the sample and blank to stand for 45 min and measured extinction E2 at 340 nm. Calculated the difference between E1 and E2 for sample and blank (Kepe et al., 2006; Sowerby and Ottaway, 1966):
ΔE sample-ΔE blank = ΔE glutamate is used for the calculations
Estimation of total protein: The amount of total protein was estimated using standard kits employing methods of Lowry using folin phenol reagent (Lowery et al., 1951).
Statistical analysis: All values are expressed as mean SEM. Data were analyzed by one way ANOVA followed by Dunnett’s test and other data were evaluated using Graph Pad PRISM software. A p-value<0.05 was considered significantly different. Comparisons were made between: (a) Group I (control) vs. group II (negative control), (b) Group II (negative control) vs group III (HAEIA 200 mg kg–1), IV (HAEIA 400 mg kg–1) and V donepezil (5 mg kg–1). Symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001.
RESULTS
Effect of HAEIA on step down inhibitory avoidance: The Step Down Latency (SDL) time of group II animals were found to be reduced on comparison with group I animals significantly. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed statistical significance increase in SDL. The increase in SDL indicates increase in Short Term Memory (STM) and Long Term Memory (LTM). The results are tabulated in Table 1 and illustrated in Fig. 1.
Effect of HAEIA on water maze task: The escape latency of group II animals were increased significantly on comparison with group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant decrease in escape latency onto the hidden platform on comparison with the group II animals.
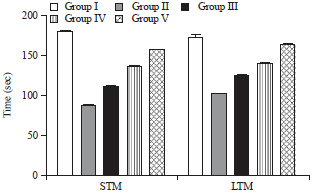 | |
| Fig. 1: | Effect of HAEIA on step down inhibitory avoidance test |
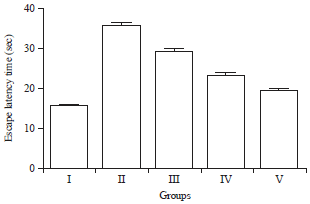 | |
| Fig. 2: | Effect of HAEIA on escape latency in water maze |
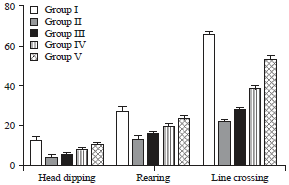 | |
| Fig. 3: | Effect of HAEIA on open field exploratory behaviour (Hole board) |
The decrease in escape latency indicates increase in memory retention. The results are tabulated in Table 2 and illustrated in Fig. 2.
Effect of HAEIA on exploratory behavior (Hole board): The group II animals exhibited a significant decrease in head dipping, rearing and line crossing on comparison with the group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed statistical significance increase in open field habituation like head dipping, rearing and line crossing on comparison with the group II animals. The results are tabulated in Table 3 and illustrated in Fig. 3.
Effect of HAEIA on Y-maze task: The percentage of alteration was reduced in group II when compared with group I animals significantly. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant increase in the alteration on comparison with group II animals. The increase in percentage alteration in Y-maze indicates increase of spatial working memory. The results are tabulated in Table 4 and illustrated in Fig. 4.
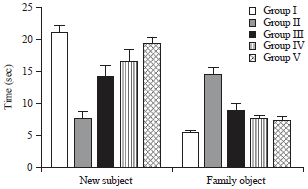
Effect of HAEIA on object recognition test: Exploration time toward newer objects was statistically decreased in group II animals compared with group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant increase in exploration time in comparison with group II animals.
| Table 1: | Step down inhibitory avoidance test |
 | |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test, STM: Short term memory, LTM: Long term memory | |
| Table 2: | Water maze task |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
| Table 3: | Open field exploratory behavior (Hole board) |
 | |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
| Table 4: | Y-maze task |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
| Table 5: | Object recognition study |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
| Table 6: | Elevated plus maze |
| Values are expressed as Mean±SEM n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
| Table 7: | Estimation of acetylcholinesterase enzyme |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
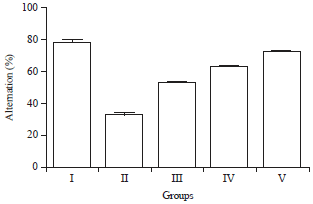 | |
| Fig. 4: | Effect of HAEIA on Y-maze task |
 | |
| Fig. 5: | Effect of HAEIA on object recognition study |
Exploration time toward familiar objects was statistically increased in group II animals compared with group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant decrease in exploration time in comparison with group II animals. The results are tabulated in Table 5 and illustrated in Fig. 5.
 | |
| Fig. 6: | Effect of HAEIA on elevated plus maze |
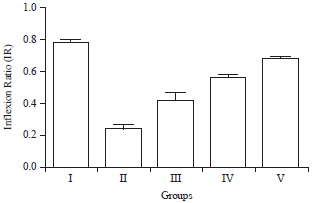
Effect of HAEIA on elevated plus maze: The Inflexion Ratio (IR) of the group II animals were decreased on comparison with the group I animals significantly. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant increase in IR in comparison with group II animals. The results are tabulated in Table 6 and illustrated in Fig. 6.
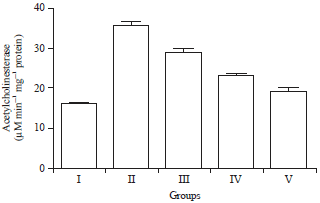
Effect of HAEIA on estimation of acetylcholineserase enzyme: Group II animals exhibited a significant increase the level of AchE on comparison with group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant reduction in the AchE enzyme level on comparison with group II animals. The results are tabulated in Table 7 and illustrated in Fig. 7.
| Table 8: | Estimation of monoamine oxidase-A |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
| Table 9: | Estimation of monoamine oxidase-B |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
| Table 10: | Estimation of dopamine |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
 | |
| Fig. 7: | Effect of HAEIA on acetylcholinesterase enzyme |
 | |
| Fig. 8: | Effect of HAEIA on estimation of monoamine oxidase-A |
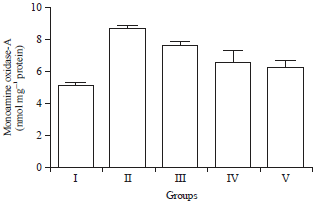
Effect of HAEIA on estimation of monoamine oxidase A and B: The level of MAO-A and MAO-B in brain of group II animals were increased significantly when compared with the control group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant reduction in the enzyme MAO-A and MAO-B level on comparison with group II animals. The results are tabulated in Table 8 and 9 and illustrated in Fig. 8 and 9.
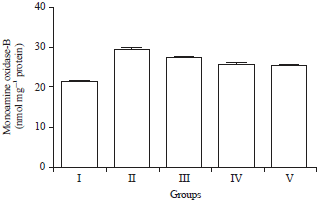 | |
| Fig. 9: | Effect of HAEIA on estimation of monoamine oxidase-B |
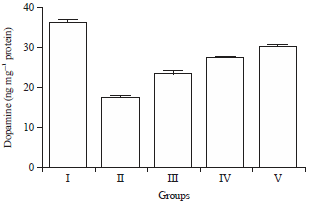 | |
| Fig. 10: | Effect of HAEIA on dopamine |
Effect of HAEIA on estimation of dopamine: The brain dopamine level in group II animals were reduced significantly on comparison with group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant increase in the level of dopamine on comparison with group II animals. The results are tabulated in Table 10 and illustrated in Fig. 10.
| Table 11: | Estimation of noradrenaline |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
| Table 12: | Estimation of serotonin |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
| Table 13: | Estimation of glutamate |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
| Table 14: | Estimation of total protein |
| Values are expressed as Mean±SEM, n = 6, symbols represent statistical significance: *p<0.05, **p<0.01, ***p<0.001, one-way ANOVA followed by Dunnett’s test | |
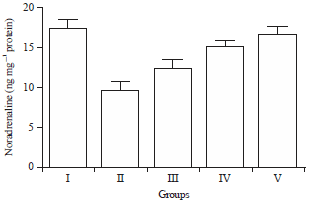 | |
| Fig. 11: | Effect of HAEIA on noradrenaline |
Effect of HAEIA on estimation of noradrenaline: The brain nor-adrenaline level in group II animals were reduced significantly on comparison with group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant increase in the level of nor-adrenaline on comparison with group II animals. The results are tabulated in Table 11 and illustrated in Fig. 11.
Effect of HAEIA on estimation of serotonin: The brain serotonin and nor-adrenaline level in group II animals were reduced significantly on comparison with group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant increase in the level of serotonin on comparison with group II animals. The results are tabulated in Table 12 and illustrated in Fig. 12.
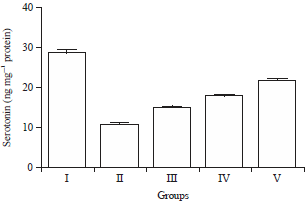 | |
| Fig. 12: | Effect of HAEIA on serotonin |
Effect of HAEIA on estimation of glutamate: The glutamate in the brain of group II animals were increased significantly on comparison with group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant reduction in the level of glutamate on comparison with group II animals. The results are tabulated in Table 13 and illustrated in Fig. 13.
Effect of HAEIA on estimation of total protein: There was significant increase in the level of total protein in group II animals on comparison with group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant decrease in the level of total protein and protein carbonyl content on comparison with group II animals. The results are tabulated in Table 14 and illustrated in Fig. 14.
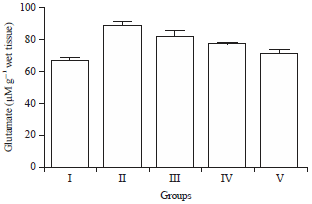 | |
| Fig. 13: | Effect of HAEIA on glutamate |
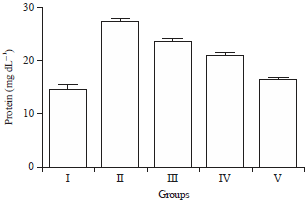 | |
| Fig. 14: | Effect of HAEIA on total protein |
DISCUSSION
Cognitive impairment is a major problem, which eventually develops in AD. It contributes to the patients functional disability and loss of skills. The pathological hallmark of Alzheimer’s disease is deposition of amyloid-beta protein in the brain. The crucial role of the central cholinergic system for memory and learning has been confirmed in a wide range of behavioural studies (Morris, 1984; Koppen et al., 1996). The Morris water maze is one of the most popular paradigms for testing learning and memory and was therefore chosen for the present study and the result of the in vivo study shows that treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant decrease in escape latency onto the hidden platform on comparison with the group II animals.
It was observed that treatment with HAEIA significantly ameliorated cognitive deficits in Aβ 25-35 injected mice. Step down inhibitory avoidance task is a classic task of memory with a strong aversive component. HAEIA significantly improves both the STM and LTM at the dose level of 200 and 400 mg kg–1.
Intra cerebro ventricular injection of Aβ 25-35 induced impairment of memory was assessed by passive avoidance, object recognition test in which object recognition task is one of the best task for evaluating the aversion of animal towards new things which impact on learning memory. It was found that animal treated with beta amyloid (25-35) shown statistically decreased exploration time toward newer objects when compared with group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant increase in exploration time.
Increased percentage of alterations observed in Y maze task of animals treated with HAEIA at 200 and 400 mg kg–1 similar type of results obtained in open field exploratory behavior, animal treated with HAEIA shows increase line crossing when compare to group II animals. Further in elevated plus maze task there was significant decrease in IR of group II animals when compare to beta amyloid injected group whereas treatment with HAEIA and standard drug shown significant increase in IR.
There are two types of cholinesterase, AChE and butyrylcholinesterase (BuChE). The AChE is found primarily in the blood and neural synapses. The AChE hydrolyzes acetylcholine (ACh) more quickly and BuChE hydrolyzes butyrylcholine (BuCh) more quickly. Increased level of AChE level further up regulates the processing and deposition of beta amyloid in AD. Presence of amyloid protein triggers the event of microglial activation and there by promotes the neuroinflammation leads to degeneration of cholinergic and adrenergic neurons. Loss of neurons and decreased neuronal count found in AD projects decreased level of acetylcholine, serotonin, dopamine and noradrenaline.
The Aβ 25-35 has the potential to induce oxidative stress in the brain cholinergic hypo function, elevation of AChE and MAO (Casanova et al., 1985; Takehashi et al., 2002). Moreover, it has been reported that it induces the production of hydrogen peroxide and lipid peroxide in hippocampal neurons of the rat brain (Yatin et al., 2000). In the present study, it was found that a significant increase in the level of AChE and MAO in mice brain after a single injection with Aβ 25-35. The AChE activity has been shown to be increased around Aβ plaques in Alzheimer’s brain. The calcium influx followed by oxidative stress is involved in the increase in activity of AChE induced by Aβ 25-35 peptide, decreasing cell membrane order and ultimately leading to the exposure of more active enzyme. The observation that Aβ peptide increases AChE activity indicates that it can be possible to ameliorate cholinergic function, by inhibiting Aβ induced increase in AChE activity. The AChE activity in the brain was increased in mice treated with Aβ 25-35 when compared with the normal. It has been postulated that the reduction in Ach release may lead to enhanced secretion of amyloid protein, resulting in further progression of the Alzheimer’s disease (Auld et al., 1998).
The MAO-A and MAO-B in the brain have been implicated in the etiology of Alzheimer’s disease. Elevations in MAO-A in Alzheimer’s neurons have been linked to increase in neurotoxic metabolites and neuronal loss. Free radical generation by Aβ or other noxious stimuli could contribute to an imbalance between the production of nitric oxide and oxygen radicals and precipitate in oxidative stress.
Recent reviews have examined the role of the noradrenergic system in emotional memory and the influence of norepinephrine on fear circuitry and the function of norepinephrine system in general. Learning to recognize important clues in our environment with emotional salience, such as danger or altruistic social interactions, is an essential survival mechanism (Van Stegeren, 2008).
Results of brain neurotransmitter estimation reveals that group II animal injected with beta amyloid showed significant increase in the level of AChE and MAO, increase in the level of these hallmark enzymes leads to decrease in the level of essential neurotransmitters like acetylcholine, dopamine and serotonin.
Serotonergic system plays a significant role in learning and memory, in particular by interacting with the cholinergic, glutaminergic, dopaminergic system. Its action is mediated via specific receptors located in crucial brain structures involved in these functions, primarily the septo-hippocampal complex and the nucleus basalis magnocellularis frontal cortex (Rodrigues et al., 2009; Smythies, 2005). Decrease in the level of serotonin may attribute to the delay in the cognitive process may further sever the condition of AD. The result of the neurotransmitter and metabolic enzyme projects that treatment with HAEIA at both the dose level of 200 and 400 mg kg–1 significantly increase the level neurotransmitter such as dopamine, serotonin and nor adrenaline this may due to the fact that HAEIA has a tendency to block the enzyme MAO and AChE there by it attenuates the memory defects causes due to amyloid deposition in AD. Further it is also evident that decreased level of AchE may contribute to the increase in acetylcholine neurotransmitter which is very vital for the process of memory and learning behavior.
The 5-hydroxytryptamine (5-HT) is a monoamine neurotransmitter. Biochemically derived from tryptophan, serotonin is primarily found in the gastrointestinal tract (GI tract), platelets and the Central Nervous System (CNS) of animals, including humans (Buhot et al., 2000).
It is also evident that amyloid deposition in the brains of AD will trigger the release of glutamate an excitatory amino acid may also have neurotoxic properties which can give rise to several forms of neurodegeneration. Glutamate neurotoxicity is commonly referred to as excitotoxicity, a term that connotes the role of an excitatory mechanism and of excitatory receptors in the toxic process. Rodents treated with glutamate causes sustain acute damage to the arcuate nucleus of the hypothalamus (Zorumski and Olney, 1993).
The result of the current study shows that glutamate level in the brain of group II animals were increased significantly on comparison with group I animals. Treatment with HAEIA (200 and 400 mg kg–1) to the group III, IV and standard drug donepezil (5 mg kg–1) to the group V showed significant reduction in the level of glutamate. This results clearly demonstrates that neuro protective effect of HAEIA.
CONCLUSION
In conclusion pharmacological screening of HAEIA against amyloid induced cognitive dysfunction in mice reveals more significant evidence based data. Treatment with low and high dose of the drug was effective in improving the short term and long term memory including exploratory and learning memory in animals. This is further supported by the performance of animal belongs to treatment group in various memory and learning related task like water maze, object recognition study, step down inhibitory avoidance, Y-maze and plus maze provided during in vivo studies.
Data’s collected from the in vitro neurotransmitters and metabolic enzyme estimation further strengthen the efficacy of the drug. It was observed that treatment with HAEIA significantly improves the level of serotonin, dopamine, nor-adrenaline. Similarly there was a significant decrease in the level of AChE, MAO (A and B) enzyme, glutamate and total protein in treatment group animals.
Result of the present study evident that plant Ipomoea aquatica halt the progression and may be used for the effective treatment and clinical management of Alzheimer’s disease. Further studies need to be carried out in clinical level to ascertain the safety and efficacy of this drug in humans.
REFERENCES
- Saitoh, A., M. Yamada, M. Yamada, S. Kobayashi, N. Hirose, K. Honda and J. Kamei, 2006. ROCK inhibition produces anxiety-related behaviors in mice. Psychopharmacology, 188: 1-11.
CrossRefPubMedDirect Link - Koppen, A., J. Klein, B.H. Schmidt, F.J. van der Staay and K. Loffelholz, 1996. Effects of nicotinamide on central cholinergic transmission and on spatial learning in rats. Pharmacol. Biochem. Behav., 53: 783-790.
CrossRefDirect Link - Auld, D.S., S. Kar and R. Quirion, 1998. β-Amyloid peptides as direct cholinergic neuromodulators: A missing link? Trends Neurosci., 21: 43-49.
CrossRefDirect Link - Siddiqui, M.B. and W. Husain, 1992. Some aquatic and marshy land medicinal plants from Hardoi district, Uttar Pradesh. Fitoterapia, 63: 245-248.
Direct Link - Balamurugan, G., M. Dharan and S. Rajan, 2009. Anti epileptic activity of poly herbal extract from Indian medicinal plants. J. Scient. Res., 1: 153-159.
Direct Link - Bera, K.M., R.S. Ghule, N.S. Kapdiya, M.B. Shah and U.M. Upadhyay, 2011. Brain tonic activity of Shankhavali Churna. Res. J. Pharm. Biol. Chem. Sci., 2: 791-797.
Direct Link - Buhot, M.C., S. Martin and L. Segu, 2000. Role of serotonin in memory impairment. Ann. Med., 32: 210-221.
CrossRefDirect Link - Casanova, M.F., L.C. Walker, P.J. Whitehouse and D.L. Price, 1985. Abnormalities of the nucleus basalis in Down's syndrome. Ann Neurol., 18: 310-313.
CrossRefDirect Link - Zorumski, C.F. and J.W. Olney, 1993. Excitotoxic neuronal damage and neuropsychiatric disorders. Pharmacol. Therapeut., 59: 145-162.
CrossRefDirect Link - Yoo, D.Y., J.H. Choi, W. Kim, H.Y. Jung and S.M. Nam et al., 2014. Cynomorium songaricum extract enhances novel object recognition, cell proliferation and neuroblast differentiation in the mice via improving hippocampal environment. BMC Complement. Altern. Med., Vol. 14.
CrossRefDirect Link - Datta, S.C. and A.K. Banerjee, 1978. Useful weeds of west Bengal rice fields. Econ. Bot., 32: 297-310.
CrossRefDirect Link - Pal, D.K., 2009. Determination of brain biogenic amines in Cynodon dactylon Pers. and Cyperus rotundus L. Treated mice. Int. J. Pharmacy Pharmaceut. Sci., 1: 190-197.
Direct Link - Dhingra, D., M. Parle and S.K. Kulkarni, 2006. β-Alanine protects mice from memory deficits induced by ageing, scopolamine, diazepam and ethanol. Indian J. Pharma. Sci., 68: 216-221.
CrossRefDirect Link - Doknark, S., S. Mingmalairak, A. Vattanajun, B. Tantisira and M.H. Tantisira, 2014. Study of ameliorating effects of ethanolic extract of Centella asiatica on learning and memory deficit in animal models. J. Med. Assoc. Thailand, 97: S68-S76.
PubMedDirect Link - Edie, H.H. and B.W.C. Ho, 1969. Ipomoea aquatica as a vegetable crop in Hong Kong. Econ. Bot., 23: 32-36.
CrossRefDirect Link - Ellman, G.L., K.D. Courtney, V. Andres Jr. and R.M. Featherstone, 1961. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol., 7: 88-90.
CrossRefDirect Link - Joshi, H. and M. Parle, 2006. Zingiber officinale: Evaulation of its nootropic effect in mice. Afr. J. Trad. Complement. Altern. Med., 3: 64-74.
Direct Link - Huang, D.J., H.J. Chen, C.D. Lin and Y.H. Lin, 2005. Antioxidant and antiproliferative activities of water spinach (Ipomoea aquatica Forsk) constituents. Botanical Bull. Acad. Sinica, 46: 99-106.
Direct Link - Hebert, L.E., P.A. Scherr, J.L. Bienias, D.A. Bennett and D.A. Evans, 2003. Alzheimer disease in the US population: Prevalence estimates using the 2000 census. Arch. Neurol., 60: 1119-1122.
CrossRefPubMedDirect Link - Hussian, G. and B.V. Manyam, 1997. Mucuna pruriens proves more effective than L-DOPA in Parkinson's disease animal model. Phytother. Res., 11: 419-423.
CrossRefDirect Link - Itoh, J., T. Nabeshima and T. Kameyama, 1990. Utility of an elevated plus-maze for the evaluation of memory in mice: Effects of nootropics, scopolamine and electroconvulsive shock. Psychopharmacology, 101: 27-33.
PubMedDirect Link - Jain, S.P. and D.M. Verma, 1981. Medicinal plants in folklore of northeast Haryana. Natl. Sci. Acad. India Newslett., 4: 269-271.
Direct Link - Joshi, H. and M. Parle, 2006. Nardostachys jatamansi improves learning and memory in mice. J. Med. Food, 9: 113-118.
CrossRefPubMedDirect Link - Sowerby, J.M. and J.H. Ottaway, 1966. The enzymic estimation of glutamate and glutamine. Biochem. J., 99: 246-252.
Direct Link - Kang, J., H.G. Lemaire, A. Unterbeck, J.M. Salbaum and C.L. Masters et al., 1987. The precursor of Alzheimer's disease amyloid A4 protein resembles a cell-surface receptor. Nature, 325: 733-736.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Schlumpf, M., W. Lichtensteiger, H. Langemann, P.G. Waser and F. Hefti, 1974. A fluorometric micromethod for the simultaneous determination of serotonin, noradrenaline and dopamine in milligram amounts of brain tissue. Biochem. Pharmacol., 23: 2437-2446.
CrossRefPubMedDirect Link - Mark, R.J., Z. Pang, J.W. Geddes, K. Uchida and M.P. Mattson, 1997. Amyloid β-peptide impairs glucose transport in hippocampal and cortical neurons: Involvement of membrane lipid peroxidation. J. Neurosci., 17: 1046-1054.
Direct Link - Weinstock, M., E. Gorodetsky, T. Poltyrev, A. Gross, Y. Sagi and M. Youdim, 2003. A novel cholinesterase and brain-selective monoamine oxidase inhibitor for the treatment of dementia comorbid with depression and Parkinson's disease. Prog. Neuro-Psychopharmacol. Biol. Psychiatry, 27: 555-561.
CrossRefDirect Link - Hiramatsu, M. and K. Inoue, 2000. Improvement by low doses of nociceptin on scopolamine-induced impairment of learning and/or memory. Eur. J. Pharmacol., 395: 149-156.
CrossRefPubMedDirect Link - Shah, M.H., H.D. Shah and V.P. Chaudhari, 2013. Future pharmacological armamentaria in management of alzheimer disease. Natl. J. Community Med., 4: 109-116.
Direct Link - Manvar, M.N. and T.R. Desai, 2013. Phytochemical and pharmacological profile of Ipomoea aquatica. Indian J. Med. Sci., 67: 49-60.
CrossRefDirect Link - Morris, R., 1984. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods, 11: 47-60.
CrossRefPubMedDirect Link - Sheikh, N., A. Ahmad, K.B. Siripurapu, V.K. Kuchibhotla, S. Singh and G. Palit, 2007. Effect of Bacopa monniera on stress induced changes in plasma corticosterone and brain monoamines in rats. J. Ethnopharmacol., 111: 671-676.
CrossRefDirect Link - Pereira, C., M.S. Santos and C. Oliveira, 1999. Involvement of oxidative stress on the impairment of energy metabolism induced by Aβ Peptides on PC12 cells: Protection by antioxidants. Neurobiol. Dis., 6: 209-219.
CrossRefDirect Link - Yankner, B.A., L.K. Duffy and D.A. Kirschner, 1990. Neurotrophic and neurotoxic effects of amyloid beta protein: Reversal by tachykinin neuropeptides. Science, 250: 279-282.
CrossRefDirect Link - Rodrigues, S.M., J.E. LeDoux and R.M. Sapolsky, 2009. The influence of stress hormones on fear circuitry. Ann. Rev. Neurosci., 32: 289-313.
CrossRefDirect Link - Samuelsson, G., M.H. Farah, P. Claeson, M. Hagos and M. Thulin et al, 1992. Inventory of plants used in traditional medicine in Somalia II. Plants of the families Combretaceae to Labiatae. J. Ethnopharmacol., 37: 47-70.
PubMedDirect Link - Seubert, P., C. Vigo-Pelfrey, F. Esch, M. Lee and H. Dovey et al., 1992. Isolation and quantification of soluble Alzheimer's β-peptide from biological fluids. Nature, 359: 325-327.
CrossRefPubMedDirect Link - Sharma, A., V. Parikh and M. Singh, 1997. Pharmacological basis of drug therapy of Alzheimer's disease. Indian J. Exp. Biol., 35: 1146-1155.
PubMedDirect Link - Sloane, P.D., S. Zimmerman, C. Suchindran, P. Reed, L. Wang, M. Boustani and S. Sudha, 2002. The public health impact of Alzheimer's disease, 2000-2050: Potential implication of treatment advances. Ann. Rev. Public Health, 23: 213-231.
CrossRefDirect Link - Smythies, J., 2005. Section III. The norepinephrine system. Int. Rev. Neurobiol., 64: 173-211.
CrossRefDirect Link - Laursen, S.E. and J.K. Belknap, 1986. Intracerebroventricular injections in mice: Some methodological refinements. J. Pharmacol. Methods, 16: 355-357.
CrossRefDirect Link - Palchykova, S., R. Winsky-Sommerer, P. Meerlo, R. Durr and I. Tobler, 2006. Sleep deprivation impairs object recognition in mice. Neurobiol. Learn. Memory, 85: 263-271.
CrossRefDirect Link - Takehashi, M., S. Tanaka, E. Masliah and K. Ueda, 2002. Association of monoamine oxidase A gene polymorphism with Alzheimer's disease and Lewy body variant. Neurosci. Lett., 327: 79-82.
CrossRefDirect Link - Anekonda, T.S. and P.H. Reddy, 2005. Can herbs provide a new generation of drugs for treating Alzheimer's disease? Brain Res. Rev., 50: 361-376.
CrossRefPubMedDirect Link - Kepe, V., J.R. Barrio, S.C. Huang, L. Ercoil and P. Siddarth et al., 2006. Serotonin 1A receptors in the living brain of Alzheimer's disease patients. PNAS, 103: 702-707.
CrossRefDirect Link - Van Stegeren, A.H., 2008. The role of the noradrenergic system in emotional memory. Acta Psychologica, 127: 532-541.
CrossRefDirect Link - Yatin, S.M., S. Varadarajan and D.A. Butterfield, 2000. Vitamin E prevents Alzheimer's amyloid beta-peptide (1-42)-induced neuronal protein oxidation and reactive oxygen species production. J. Alzheimers Dis., 2: 123-131.
PubMedDirect Link