
Rifat Ullah Khan
Department of Animal Production, College of Food and Agriculture Sciences, King Saud University, Riyadh 11451, Saudi Arabia
LiveDNA: 92.7542
Shabana Naz
Department of Zoology, Government College University, Faisalabad, Pakistan
Kuldeep Dhama
Division of Pathology, ICAR-Indian Veterinary Institute, Izatnagar, India
LiveDNA: 91.4710
K. Karthik
Division of Bacteriology and Mycology, ICAR-Indian Veterinary Institute, Izatnagar, India
Ruchi Tiwari
Department of Veterinary Microbiology and Immunology, College of Veterinary Sciences, Uttar Pradesh, Pandit Deen Dayal Upadhayay
Pashu Chikitsa Vigyan Vishwa Vidyalaya Evum Go-Anusandhan Sansthan (DUVASU), Mathura 281001, India
LiveDNA: 91.10804
Mutassim M. Abdelrahman
Department of Animal Production, College of Food and Agriculture Sciences, King Saud University, Riyadh 11451, Saudi Arabia
LiveDNA: 966.7438
Ibrahim A. Alhidary
Department of Animal Production, College of Food and Agriculture Sciences, King Saud University, Riyadh 11451, Saudi Arabia
Arshad Zahoor
Department of Animal Health, Faculty of Animal Husbandry and Veterinary Sciences, University of Agriculture, Peshawar, Pakistan
International Journal of Pharmacology
Year: 2016 | Volume: 12 | Issue: 3 | Page No.: 220-231







ABSTRACT
Recently, animal food industry has shown an increasing interest in Direct-Fed Microbials (DFM) to keep the concept of safe food supply at the consumer level. In the absence of suitable growth stimulant after banning the use of antibiotics in animal feed, a positive manipulation of rumen microbes has become the field research for entrepreneurs and research scientists. Direct-fed microbials is a term reserved for naturally occurring live microbes that can be supplemented orally to produce a beneficial health response in the host animal. A number of genera of live microorganisms including; bacteria, yeast and fungi are frequently used in different combination feed as DFM for domestic ruminants. It has been documented that these live culture of microbes improve ruminant productivity, milk production, immunity, digestibility of feed, counter infectious pathogens and protect health. Possible attempts have also been undertaken to justify the mechanism of these microbes. This review tries to summarize the effect of supplementation of DFM on the production, immune response, fermentation pattern and safeguarding health. The discussed concepts and advances concerning to DFM implementation will be useful not only for the researchers, animal owners, feed manufacturers, pharmacists, pharmaceutical companies, stake holders but will also boost the economic gains and profits by promoting the ruminant health and production through feed modification.
PDF Abstract XML References Citation
How to cite this article
Rifat Ullah Khan, Shabana Naz, Kuldeep Dhama, K. Karthik, Ruchi Tiwari, Mutassim M. Abdelrahman, Ibrahim A. Alhidary and Arshad Zahoor, 2016. Direct-Fed Microbial: Beneficial Applications, Modes of Action and
Prospects as a Safe Tool for Enhancing Ruminant Production and
Safeguarding Health. International Journal of Pharmacology, 12: 220-231.
DOI: 10.3923/ijp.2016.220.231
URL: https://scialert.net/abstract/?doi=ijp.2016.220.231
DOI: 10.3923/ijp.2016.220.231
URL: https://scialert.net/abstract/?doi=ijp.2016.220.231
INTRODUCTION
The concept of microbial manipulation was first coined when the consumption of yoghurt by Bulgarian peasants was associated with their longevity (Metchnikoff, 1907). Now it is generally accepted that certain group of viable microbes are beneficial for livestock species. Rumen harbours variety of microbes actively involved in initial and very first step of digestion of green fodder and concentrate diet. Age of animal, feeding pattern and diet formulation influence and determines the microbial environment of rumen in four stomach animals (Dhanasekaran et al., 2008; Apas et al., 2015; Yadav and Shukla, 2015).
The concern over the use of intensive antibiotics has triggered to the newer sense of the importance of DFM in the livestock production. The word probiotics and DFM are used interchangeably but in fact they are not the same. The DFM has been defined by the US Food and Drug Administration Authority as the feed product containing the source of live naturally existing microbes (Brashears et al., 2005). The DFM has been used in livestock industry over the last 20 years to enhance milk production, weight gain and feed efficiency (LeJeune and Wetzel, 2007). They are provided to the animals in the form of bolus or sometime mixed with the feed.
The major cause of rumen microbial study is to improve feed utilization, animal health and production, as well as animal safety by improving immune status which is possible only by controlled fermentation and lowering ruminal diseases. In the past few decades, a number of feed additives have been used for example antibiotics, sodium bicarbonate, slow release urea, methane inhibitors and defaunting agents have been successfully utilized in ruminants nutrition in order to manipulate the microbial ecosystem (Seo et al., 2010). However, due to a number of toxic problems associated with these additives, researchers are looking for safer additives in ruminant nutrition (Barton, 2000). Further, these products have been criticized by the consumers from the safety point of view.
In view of the recent criticism, the use of probiotics has become a suitable alternative that can survive in the rumen of animals (Dhama et al., 2008; Fon and Nsahlai, 2013). When suitable DFM (selected bacteria, yeast or fungi) are supplemented to the ruminants through diet, they help in establishing healthy microflora in the gut by checking the dysbiosis and undesired alterations in the pH. In early stages of neonatal life, gut colonization with DFM microbes set up healthy microbiota in the gastrointestinal tract, which plays crucial role in protecting budding calves against various infections, bacterial diarrhea and pathogenic challenges (Dhama et al., 2008; Varankovich et al., 2015; Yuan et al., 2015). Administration of selected DFM strains in young and adult both types of ruminants augment the health and production performance, improves the immunological aura in the body and compete with the invading microbes without leaving any residual toxic effects in an eco-friendly manner (Ohashi and Ushida, 2009; Yeoman and White, 2014). These multitasking approaches of DFM support and strongly recommend their optimal usage in the livestock industry.
This review discusses the DFM with a focus on their multiple beneficial roles in ruminants such as effects on performance and productivity, mitigation of methane emission, immunomodulatory properties, countering infectious pathogens and protecting health. Information compiled with the concepts and recent advances would encourage more research and validation of practical prospects of DFM supplementation in ruminant feeding and health. It will be highly useful for researchers, animal owners, feed manufacturers, pharmacists, pharmaceutical companies, stakeholders and would help in gaining economic returns by promoting ruminant health and production through feed modification.
DIRECT-FED MICROBIALS
The word probiotic is composed of two words: pro means in favour and biotic means life. The term probiotic has been defined as "Those living organisms which are used as feed supplement and produced beneficial impact on the host animal through improving the intestinal microbial balance" (Fuller, 1989). The Food and Drug administration of USA has directed to use the term "direct-fed microbial" instead of probiotic (Miles and Bootwalla, 1991). The definition of DFM is broad in nature and includes variety of organisms like yeast, fungi, bacteria and cell fragments (Oetzel et al., 2007; Elghandour et al., 2014b). The DFM grow in the rumen has a beneficial effect on microbial ecosystem (Seo et al., 2010). Variety of DFM is being used in the ruminants around the world which can be classified into three major classes: bacterial, fungal or combination of both. The most common among them is the bacterial DFM which can further be classified as lactic acid producing bacteria (LAB), second is lactic acid utilizing bacteria (LUB) and other microorganisms (Elghandour et al., 2015). The most common genus used in DFM are Propionibacterium, Streptococcus, Lactobacillus, Bacillus and some other important bacteria such as, Prevotella bryantii and Megasphaera elsdenii (Kung, 2006; Seo et al., 2010). The use of these bacteria allow the producers to decrease the time in adaptation to high concentrate diet and reduce the incidence of acidosis in lactating animals (Kung, 2006; Elghandour et al., 2015).
TYPES OF DFM
Rumen microbes: Generally the population of rumen microbes is composed of different kinds of bacteria, protozoa and fungi, only a small number of these communities qualify for their potential as DFM (Miron et al., 2001). Most of the studies have focused on using such a DFM targeting the metabolism of lactic acid such as, Selenomonas ruminantium, Megasphaera elsdenii and Propionibacterium freudenreichii (Wiryawan and Brooker, 1995; Klieve et al., 2003; Raeth-Knight et al., 2007). Other approach includes the utilization and reduction of lactic acid such as, Prevotella bryantii 25A which consume starch (Chiquette et al., 2008). Another group of DFM includes fibrolytic rumen bacteria namely Ruminococcus albus and Ruminococcus flavefaciens in an effort to enhance fiber digestion (Krause and Otzel, 2006; Chiquette et al., 2007). Logically DFM derived from the rumen may possibly be more capable to adjust into the microbial community since they are origined from the environment from which they have been extracted eliminating the need of daily administration (McAllister et al., 2011).
Lactic acid producing bacteria: Majority of DFM used in cattle production are lactic acid producing bacteria (LAB) including Enterococcus spp., Streptococcus spp., Lactobacillus spp. and Pediococcus spp. (McAllister et al., 2011). This class of DFM has been used in almost all types of livestock. In suckling calves, LAB are administered as bolus while in dairy and beef animals, they are mixed with diet. Furthermore, LAB are also inoculated into the forage before the process of ensiling in order to enhance the preservation, increasing feeding value and aerobic stability of silage (Schmidt et al., 2009).
Lactic acid producing bacteria are more effective since they are environment friendly and can alter the environment through a number of mechanisms. The lactic acid produced by LAB is one of the key compounds that can change the pH of the competitor bacteria, in addition, to production of antimicrobial substance known as bacteriocin (Servin, 2004; McAllister et al., 2011). Other compounds such as mevalonolactone, benzoic acid, diacetyl, reuterin and methylhydantoin are also produced by some of the LAB (Brashears et al., 2005). It has also been suggested that LAB in combination of other bacteria and yeast showed synergistic activities in many of the commercial products (McAllister et al., 2011).
Other bacteria: Some other types of bacteria such as Bifidobacterium spp. and Bacillus spp. have been successfully used as DFM. The endospores produced by Bacillus spp. are environmentally stable and thermo-tolerant which make these bacteria competitively superior in ensuring their survival during storage or pelleting (McAllister et al., 2011). Bacillus spp. are present in rumen in low number and play a minor role in the degradation of cell wall. Strains of Bifidobacterium used as DFM do not originate from the rumen, however, they play key role in starch digestion (Stewart et al., 1997).
Yeast culture: The official definition of yeast culture is a dry product composed of yeast and the media on which it was grown dried in such a manner to preserve the fermenting capacity of the yeast (Yoon and Stern, 1995). The effect of yeast preparation on the rumen fermentation and performance has been well elucidated and has generated considerable scientific interest in the past few decades. It is now well accepted that yeast culture can beneficially modify rumen microflora, digestive and fermentative activities in the rumen.
First published report on the use of the yeast was made public in 1925 describing the use of yeast as a protein source in ruminant ration in lactating cows (Eckles and Williams, 1925). The use of live yeast has been associated with high production of ethanol toxicity; therefore, dead yeast is preferred to be used in animal diet (Yoon and Stern, 1995). The application of yeast increased during 1950’s when, Renz and Koch (1956) reported that inclusion of 50 g day–1 yeast culture increased milk yield.
Saccharomyces cerevisiae is being extensively used as a DFM for the improved performance and milk yield in dairy cattle. Recent analysis has demonstrated that S. cerevisiae in dairy cows improved feed intake, rumen pH and volatile fatty acids, organic digestibility and decreased lactic acid in the rumen (Desnoyers et al., 2009). Saccharomyces cerevisiae are aerobic bacteria which can metabolize lactic acid, alter the population of rumen microbes due to its unique ability to utilize oxygen thereby producing an environment which is more suitable for the growth of anaerobic bacteria in the rumen and increase the fibrolytic bacteria (Jouany et al., 1999; McAllister et al., 2011). In addition to Saccharomyces cerevisiae, Aspergillus oryzae and Aspergillus niger have also been used in ruminant nutrition, however, their role is limited to crude enzyme extract instead of whole cell, primarily targeted to increase fibre digestion in the rumen (McAllister et al., 2011).
For ruminants, yeast culture has been considered as the most promising DFM for nutrient utilization. Previously it was suggested that yeast cannot survive and multiply in the anaerobic culture, however, later evidences suggest that Saccharomyces cerevisiae may multiply and grow in the rumen conferring beneficial impact on cellulysis and productive traits (Dawson and Newman, 1988; Harris and Lobo, 1988; Dutta et al., 2009).
MECHANISM OF ACTION OF DFM
Many factors are involved in understanding the mode of action of DFM such as dose, feeding time, duration and frequency and strains. Moreover, some DFM act via rumen while other influence the gastrointestinal tract (Puniya et al., 2015). Inside rumen the type of DFM such as LUB or LAB influence, the later mainly prevent the acidosis of the rumen in dairy animals through facilitating the rumen microbes which can survive in the presence of lactic acid in the rumen (Yoon and Stern, 1995; Nocek et al., 2002). The former type of bacteria mainly decreases the concentration of lactic acid and keeps the pH at normal level. One such example is Megasphaera elsdenii which utilize the lactic acid in the rumen (Yang et al., 2004; Kung, 2006). Furthermore, this bacteria utilize lactate, glucose and maltose in addition to competing with lactate producing organism (Russell and Baldwin, 1978). Propionibacteria is present in the rumen in high number in animals fed with medium concentrate diet which modify the rumen conditions through conversion of lactate into propionate resulting in the higher production of hepatic glucose (Stein et al., 2006).
The DFM has also been proposed to produce antibacterial compounds such as bacteriocin and hydrogen peroxide which competitively exclude the pathogenic bacteria. Hydogen peroxide can characteristically oxide the sulfhydryl group in hexokinase and glycerol aldehyde -3-phosphate dehydrogenase causing blockage of glycolysis (Carlsson et al., 1983; Dicks and Botes, 2010). The bacteriocin produced by LAB does not allow the binding of substrates to ribonucleotide reductase interfering with the DNA synthesis of target microbes (Dicks and Botes, 2010).
Recently, it has been proposed that DFM can modulate the host immune system through stimulating and activation of the dendritic cells, natural killer cells, macrophages, T and B lymphocytes and neutrophils of the intestines (Krehbiel et al., 2003). When the DFM is administered, they are absorbed by the intestinal wall and various immune players such as dendritic cells, macrophages and natural killer cells surround them which result in the stimulation of the immune response (Dicks and Botes, 2010). The LAB such as Lactobacillus casei Shirota and Lactobacillus rhamnosus Lr23 is also involved in the stimulation of macrophages to produce the cytokines (TNF-α) as reported by Matsuguchi et al. (2003).
Many theories have been forwarded in explaining the mechanism through which yeast exerts their beneficial impact on host. Yeast may have buffering effect through reduction of pH (Elghandour et al., 2014a, b). Yeast can also improve the oxygen availability on the surface of freshly ingested food to maintain metabolism (Newbold et al., 1996). Furthermore, DFM such as S. cerevisiae can compete with the starch utilizing bacteria thereby preventing the accumulation of lactate and provides organic acids and vitamins to the cellulytic bacteria and LUB (Chaucheyras et al., 1995; Lynch and Martin, 2002).
The rumen pH is mainly determined by the concentration of lactic acid (Williams et al., 1991). Fermentation of carbohydrate depresses pH leading to reduction in the number of cellulytic bacteria, impair degradation of forage and dry matter intake (Orskov et al., 1978; Thomas and Rook, 1981; Williams, 1989).
BENEFICIAL APPLICATIONS OF DFM
Various beneficial applications of DFM in ruminants viz., effect on ruminant performance, productivity, immunomodulatory activities, protecting from pathogens/ infections, safeguarding health and mitigation of methane emission are discussed in below sections. An overview of modes of actions and beneficial applications of DFM for enhancing ruminant production and protecting health is depicted in Fig. 1.
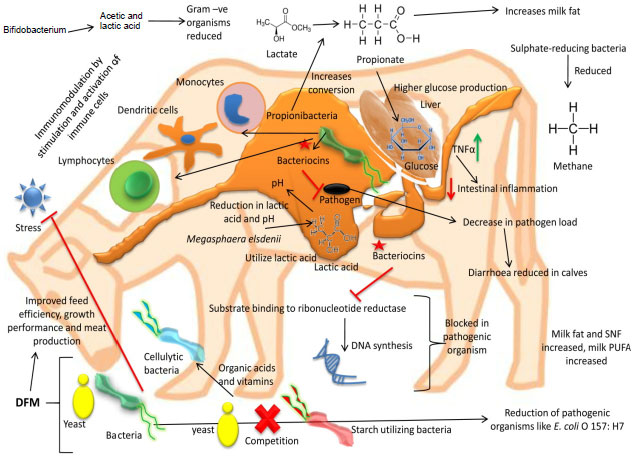 | |
| Fig. 1: | An overview of modes of actions and beneficial applications of DFM for enhancing ruminant production and protecting health |
EFFECT OF DFM ON RUMINANT PERFORMANCE
The purpose of feeding DFM prior to the start of rumination is to establish and maintain stable normal intestinal microbes rather than a production stimulant. In neonates, the microbes of the gastrointestinal tract are very unstable, abrupt dietary or environmental change may cause an alteration in the microbial population of the GIT which may result in diarrhea (Sandine, 1979). Most of the studies dealing with the use of DFM in ruminant production are related with neonatal dairy calves. In young calves, the rapid establishment to adapt to solid feed by stabilizing the rumen and intestinal microbes and the exclusion of enteric pathogen is the primary goal of the use of DFM (Dhama et al., 2008; Puniya et al., 2015).
Experimentally, there are several bacteria which can potentially act as DFM but they have not been commercialized. One such bacteria is M. elsdenii which is used in lactating animals, however, when cows are abruptly shifted from high forage diet to concentrate diet, M. elsdenii is often incapable to prevent lactic acidosis (Puniya et al., 2015). Similarly, E. faecium succeeded less likely in feedlot cattle adapted to high grain diet (Beauchemin et al., 2003).
The several conditions faced by newly produced calves such as weaning, traction and dehorning has effect on microbial environment (Elghandour et al., 2015). These conditions may change the microbiota in the negative directions thereby reducing the performance and weight gain. The administration of DFM is expected to change the microbial population in the positive direction (Krehbiel et al., 2003). Limited data is available on the administration of DFM in stressed calves, however, there is a general consensus that supplementation of DFM to the diet of such calves may improve the health and performance of stressed calves (Elghandour et al., 2015).
Commonly used DFM in young calves are Streptococcus and Lactobacillus species. In contrast to adult ruminants, young ruminants have the ability to digest a significant amount of ration which is associated with the risk of increased proliferation of microorganisms resulting in the increased chance for diarrhea and also weight loss (Elghandour et al., 2015). Here the role of DFM is very important which may obtain positive impact on the GIT function thereby modifying the bacterial concentration in the positive direction (Abu-Tarboush et al., 1996; Kung, 2001). It is the reason of using DFM that in an experiment conducted by Nakanishi et al. (1993) in which Holstein calves supplemented with Lactobacillus acidophilus showed rumination at 30 day compared to untreated calves, indicating the ability of this bacterium to promote ruminal development (Elghandour et al., 2015). Recently, Dicks and Botes (2010) concluded that Bifidobacteria produce acetic and lactic acid at such a ratio (3:2) which may be beneficial for the control of Gram-negative bacteria in the GIT, probably the shift towards the more production of acetate is lethal for the survival of Gram-negative bacteria and other microbes such as moulds and yeast (Gilliland 1989; Elghandour et al., 2015). Growth performance has also been shown to improve when LAB was supplemented in young calves (Adams et al., 2008; Frizzo et al., 2010). Increased reduction in the incidence of diarrhea has been reported by using DFM especially Lactbacillus (Abu-Tarboush et al., 1996).
The DFM have been rarely used in high producing dairy cows, however, its use during early lactation may be the best candidate since cows are in negative energy balance with high fermentable carbohydrate which is associated with lactic acidosis (Kung, 2006). It has been shown that DFM improves performance in dairy cows through high intake of dry matter and enhance the milk yield and milk contents such as protein, blood sugar and insulin concentration (Nocek et al., 2003; Nocek and Kautz, 2006; Oetzel et al., 2007). In beef cattle, to prevent ruminant acidosis caused by high fermentable diet, DF is recommended. The DFM in beef cattle showed improved feed efficiency, growth performance and meat production (Ghorbani et al., 2002; Krehbiel et al., 2003). The supplementation of DFM in the diet of feedlot ruminants has caused improved feed efficiency and daily gain, carcass characteristics and dry matter intake (Huck et al., 1999; Galyean et al., 2000).
The most important role of DFM in feedlot cattle is the reduction of pathogenic bacteria such as, Escherichia coli in the GIT (Elghandour et al., 2015). Studies have shown that different kinds of DFM have reduced the population of E. coli O 157: H7 in feedlot cattle (Ohya et al., 2000).
Composition of diet and environmental conditions determines the growth response of animals in case of yeast supplementation, however, type of animals may not be affected since daily live weight were equally increased in dairy and beef calves (Hughes, 1988).
Williams et al. (1987) demonstrated higher daily weight gain in lambs fed a diet supplemented with yeast culture. In contrast, Quigley et al. (1992) found that yeast culture affected ruminal metabolites with no influence on dry matter intake and weight gain in calves. Similarly, Mir and Mir (1992) observed that supplemental live-yeast did not positively impact the feed utilization in steers.
MILK PRODUCTION AND COMPOSITION
Modern dairy farms are targeting high milk production utilizing feed composed of high concentrates to meet the metabolic demand of the higher milk production. Such feeding system is associated with metabolic dysfunction like rumen acidosis especially during poor feeding condition and composition.
Higher milk yield with no change in milk composition has been reported due to the supplementation of Lactobacillus acidophilus (Jaquette et al., 1988). Colenbrander et al. (1988) found that treatment of alfalfa with L. acidophilus improved milk production and composition in dairy cows with no effect on dry matter intake. Improved milk response has been reported in dairy cows and goats fed with yeast culture with increased milk fat, however the magnitude of response depends upon the stage of lactation (Williams et al., 1991). Response of milk production may also depends upon production level for example, Hoyos et al. (1987) reported that dairy cows supplemented under high and low level of production with Saccharomyece cerevisiae, S. faecium and L. acidophilus. High production cows showed higher milk yield with no effect on the cows with low production, however, fat was similar in both the groups. Milk production in high producing cows was increased when M. elsdenii NCIMB 41125 was supplemented as compared to the control group (Erasmus et al., 1992; Hagg and Henning, 2007; Aikman et al., 2008). Gomez-Basauri et al. (2001) reported higher milk production (0.73 kg day–1) with less feed consumption (0.42 kg day–1) when cows were doses with lactic acid bacteria including L. casei, L. acidophilus and E. faecium in comparison to the control. On the same line, Boyd et al. (2011) found that using L. acidophilus NP51 and Proponibacterium freudenreichii NP24 may improve milk yield and digestibility in heat stressed cows. It is interesting that using a different strain (L. acidophilus LA747 and P. freudenreichii PF24) in the study of Raeth-Knight et al. (2007) found no effect on digestibility, performance and rumen characteristics. Other studies have shown higher milk yield when lactating cows were supplemented with a combination of fungal culture (Saccharomyces cerevisae) and lactic acid bacteria acidophilus and/or (Lactobacillus plantarum/E. faecium) (Komari et al., 1999; Block et al., 2000). Propionibacteria converts lactic acid into propionic acid, are very important for the ruminants since the propionate represents a major source of energy of the animals and a DFM composed of such bacteria increased milk fat as well as milk yield in dairy cows (Nocek and Kautz, 2006; Oetzel et al., 2007).
In lactating cows, yeast culture significantly improved the dry matter intake and subsequent milk yield (Williams et al., 1991). Similarly, Wohlt et al. (1991) reported higher feed intake and milk yield when yeast culture was supplemented from 30 days pre-partum till week 18 of lactation. Pre-partum and post-partum dry matter intake and milk production have been reported by several authors in dairy animals (Wohlt et al., 1998; Dann et al., 2000; Nocek et al., 2003). Nikkhah et al. (2004) did not find any advantage on the intake of dry matter and milk production in dairy cows, however, milk fat and total solids were increased by the supplementation of live yeast culture.
IMMUNOMODULATION
A number of mechanisms are involved in improving the immune system of the host such as upregulating cell-mediated immune response, augmenting antibody production, enhancing dendritic cell-T interaction, reduction of epithelial cells apoptosis, production of Toll like receptor signaling etc. (McAllister et al., 2011). The DFM stimulates epithelial innate immunity which may suppress intestinal inflammation by increasing the production of TNF-α (Pagnini et al., 2010). The way the DFM affect the production of cytokines and chemokines in addition to T and B cell responses depends upon the types of DFM, the dose and duration of the experiment (McAllister et al., 2011). For example, a DFM consisting of Bifidobacterium thermophilum, L. casei, E. faecium and L. acidophilus heightened the expression of cytokines IL-6 but decreased cytokine IL-10 in chicken (Chichlowski et al., 2007). In another report LAB-based DFM depressed IFN- γ, IL-3 and IL-4 in chicks and also reduced the Salmonella enterica colonies in the intestines of chickens (Haghighi et al., 2008). Another study reported that in response to L. acidophilus DFM, IFN- γ, IFN- α, IL-18, STAT4, STAT2 and MyD88 were upregulated in the cecal tonsils of broilers (Brisbin et al., 2008).
It has also been proposed that DFM mediated immune response is more important in young ruminants since intestinal population is less established on one hand and on the other hand, intestinal tract is more susceptible to colonization of pathogenic microbes (McAllister et al., 2011). For example, Lactococcus lactis DPC 3147 was administered to adult lactating cows which successfully increased IL-1β and IL-8 gene expression but failed to show a response to control mastitis (Beecher et al., 2009).
PROTECTING FROM INFECTIOUS PATHOGENS
Probiotics such as, lactic acid bacteria have been reported to prevent and protect cattle from various pathogens, alleviate stress and boost immunity and have been suggested to be valuable alternative to antibiotics (Gupta and Gupta, 2007; Yasuda et al., 2007; Frizzo et al., 2010; Bayatkouhsar et al., 2013; Uyeno et al., 2015). The DFM Yeast as probiotic has been found effective in decreasing pathogenic effects of infectious bovine rhinotracheitis virus in calves (Cole et al., 1992). Probiotics strains were found to prevent and reduce mastitis and metritis in cattle by reducing adhesion of pathogenic bacteria, producing antimicrobial substances and with other modes of action (Otero et al., 2006; Gulbe et al., 2015). Lactobacillus acidophilus has been reported to provide protection from E. coli O157:H7 infection in cattle owing to their bacteriostatic/bacteriocidal effects (Schamberger et al., 2004; Poppi et al., 2015). Lactobacillus plantarum supplementation in feed reduced Clostridia load in faeces of goats (Maragkoudakis et al., 2010). A probiotic mixture of Bacillus licheniformis and B. subtilis supplementation in feed of sheep resulted in lesser lamb mortality (Kritas et al., 2006). A mixture of L. acidophilus, L. helveticus, L. bulgaricus, L. lactis, Streptococcus thermophilus and Enterococcus faecium given to sheep was found effective in checking faecal shedding of non-O157 Shiga toxin-producing E. coli (STEC) strain, an important food-borne pathogen of humans having public health concerns (Rigobelo et al., 2014). Probiotics could inhibit Listeria monocytogenes and their use has been suggested to prevent and control this important pathogen having public health concerns (Dhama et al., 2015).
MITIGATION OF METHANE EMISSION
Methane is a global concern and ruminants are though to contribute 12-15% to the total global methane emission. Lactating cows consuming 14.7% of gross energy from dry matter is estimated to produce 419 L of methane/day/cow (Holter and Young, 1992). It has been documented that cattle in developed countries are emitting 55 kg/day/animal methane gas compared to 35 kg/day/animal in developing countries. Methane is an important gas and its contribution towards the global warming is 25 times greater than carbon dioxide (Jeyanathan et al., 2014).
Different strategies have been applied, among them the most important is the idea of DFM application. It was reported that addition of yeast culture reduced methane production in steers by 28% (Williams, 1989). Lynch and Martin (2002) reported a 20% methane reduction after a period of 48 h incubating mixed culture of rumen microorganisms in the presence of alfalfa. Chaucheyras-Durand et al. (2008) discussed three major effects of yeast on the rumen health through stabilizing rumen pH, feed degradation, establishing of microbes in the rumen. In addition to yeast, propionate production is the major biochemical pathway through bacterial DFM (Seo et al., 2010). Propionate-producing bacteria utilize hydrogen and therefore reduce the methane production. It was reported that when a DFM based on M. elsdenii is used, the rumen pattern is shifted in favour of propionate production (Jeyanathan et al., 2014). Other bacteria such as Prevotella ruminicola is also known for propionate formation through acrylate pathway (Wallnofer and Baldwin, 1967). Although propionate producing bacteria have been used to improve animal production. However, little attention has been given to methane production. An encouraging study of Berger et al. (2012) using a mixed culture of Propionibacterium jensenii-Lactobacillus spp. produced little methane production in lactating dairy cows showing the potential to mitigate rumen methane. Nitrate/nitrite-reducing bacteria can also play role in reducing rumen methane to act as H2 sink to CO2 in the rumen.
Sulphate Reducing Bacteria (SRB) have also been used to reduce the methane production in the rumen. The SRB competes with methane producing bacteria for common substrate such as H2, format and acetate. The bacteria of SRB in the rumen consisted of genus Desulfovibrio and Desulfotomaculum. The introduction of sulphate is directly linked with the ability of SRB to compete with methanogenic bacteria. Paul et al. (2011) reported decreased methane production using the newly indentified, Fusobacterium sp., as a DFM.
CONCLUSION AND FUTURE PROSPECTS
Manipulation of gastrointestinal microbial ecosystem to augment animal performance and safeguarding health is one of the prime goals of animal scientists and veterinarians. In light of worldwide ban on the use of antibiotics and pressure to produce higher animal protein, the DFM offer an alternative option. This review has explored the production response with the supplementation of DFM on various aspects. In addition to positive impact of DFM on ruminant production, literature has also reported inconsistency in the response of animals which has been attributed to nature of microbial supplement, dose and duration, age of animal, environmental conditions and combination of these factors. In view of the increasing pressure to produce more and safe animal protein, future DFM research must be directed towards decreasing greenhouse gases and carbon footprint per unit of animal protein produced.
ACKNOWLEDGMENTS
This study was supported by King Saud University, Deanship of Scientific Research, College of Food and Agriculture Sciences, Research centre. All the authors acknowledge respective institutions and universities.
REFERENCES
- Abu-Tarboush, H.M., M.Y. Al-Saiady and A.H.K. El-Din, 1996. Evaluation of diet containing Lactobacilli on performance, Fecal Coliform and Lactobacilli of young dairy calves. Anim. Feed Sci. Technol., 57: 39-49.
CrossRefDirect Link - Adams, M.C., J. Luo, D. Rayward, S. King, R. Gibson and G.H. Moghaddam, 2008. Selection of a novel direct-fed microbial to enhance weight gain in intensively reared calves. Anim. Feed Sci. Technol., 145: 41-52.
CrossRefDirect Link - Yang, W.Z., K.A. Beauchemin, D.D. Vedres, G.R. Ghorbani, D. Colombatto and D.P. Morgavi, 2004. Effects of direct-fed microbial supplementation on ruminal acidosis, digestibility and bacterial protein synthesis in continuous culture. Anim. Feed Sci. Technol., 114: 179-193.
CrossRefDirect Link - Beauchemin, K.A., W.Z. Yang, D.P. Morgavi, G.R. Ghorbani, W. Kautz and J.A. Leedle, 2003. Effects of bacterial direct-fed microbials and yeast on site and extent of digestion, blood chemistry and subclinical ruminal acidosis in feedlot cattle. J. Anim. Sci., 81: 1628-1640.
PubMedDirect Link - Beecher, C., M. Daly, D.P. Berry, K. Klostermann and J. Flynn et al., 2009. Administration of a live culture of Lactococcus lactis DPC 3147 into the bovine mammary gland stimulates the local host immune response, particularly IL-1β and IL-8 gene expression. J. Dairy Res., 76: 340-348.
CrossRefPubMedDirect Link - Chaucheyras-Durand, F., N.D. Walker and A. Bach, 2008. Effects of active dry yeasts on the rumen microbial ecosystem: Past, present and future. Anim. Feed Sci. Technol., 145: 5-26.
CrossRefDirect Link - Chichlowski, M., J. Croom, B.W. McBride, L. Daniel, G. Davis and M.D. Koci, 2007. Direct-fed Microbial primalac and salinomycin modulate whole-body and intestinal oxygen consumption and intestinal mucosal cytokine production in the broiler chick. Poult. Sci., 86: 1100-1106.
CrossRefPubMedDirect Link - Chiquette, J., M.J. Allison and M.A. Rasmussen, 2008. Prevotella bryantii 25A used as a probiotic in early-lactation dairy cows: Effect on ruminal fermentation characteristics, milk production and milk composition. J. Dairy Sci., 91: 3536-3543.
CrossRefDirect Link - Cole, N.A., C.W. Purdy and D.P. Hutcheson, 1992. Influence of yeast culture on feeder calves and lambs. J. Anim. Sci., 70: 1682-1690.
Direct Link - Dann, H.M., J.K. Drackley, G.C. McCoy, M.F. Hutjens and J.E. Garrett, 2000. Effects of yeast culture (Saccharomyces cerevisiae) on prepartum intake and postpartum intake and milk production of jersey cows. J. Dairy Sci., 83: 123-127.
CrossRefDirect Link - Desnoyers, M., S. Giger-Reverdin, G. Bertin, C. Duvaux-Ponter and D. Sauvant, 2009. Meta-analysis of the influence of Saccharomyces cerevisiae supplementation on ruminal parameters and milk production of ruminants. J. Dairy Sci., 92: 1620-1632.
CrossRefDirect Link - Dhama, K., M. Mahendran, S. Tomar and R.S. Chauhan, 2008. Beneficial effects of probiotics and prebiotics in livestock and poultry: The current perspectives. Intas Polivet, 9: 1-12.
Direct Link - Dhanasekaran, D., S. Saha, N. Thajuddin and A. Panneerselvam, 2008. Probiotic effect of Lactobacillus isolates against bacterial pathogens in Claris orientalis. Facta Univ.: Med. Biol., 15: 97-102.
Direct Link - Dicks, L. and M. Botes, 2010. Probiotic lactic acid bacteria in the gastro-intestinal tract: Health benefits, safety and mode of action. Beneficial Microbes, 1: 11-29.
CrossRefPubMedDirect Link - Elghandour, M.M.Y., A.Z.M. Salem, J.S.M. Castaneda, L.M. Camacho, A.E. Kholif and J.C.V. Chagoya, 2015. Direct-fed microbes: A tool for improving the utilization of low quality roughages in ruminants. J. Integr. Agric., 14: 526-533.
CrossRefDirect Link - Elghandour, M.M.Y., J.C.V. Chagoyan, A.Z.M. Salem, A.E. Kholif, J.S.M. Castaneda, L.M. Camacho and G. Buendia, 2014. In vitro fermentative capacity of equine fecal inocula of 9 fibrous forages in the presence of different doses of Saccharomyces cerevisiae. J. Equine Vet. Sci., 34: 619-625.
CrossRefDirect Link - Elghandour, M.M.Y., J.C.V. Chagoyan, A.Z.M. Salem, A.E. Kholif, J.S.M. Castaneda, L.M. Camacho and M.A. Cerrillo-Soto, 2014. Effects of Saccharomyces cerevisiae at direct addition or pre-incubation on in vitro gas production kinetics and degradability of four fibrous feeds. Ital. J. Anim. Sci., 13: 295-301.
CrossRefDirect Link - Erasmus, L.J., P.M. Botha and A. Kistner, 1992. Effect of yeast culture supplement on production, rumen fermentation and duodenal nitrogen flow in dairy cows. J. Dairy Sci., 75: 3056-3065.
CrossRefDirect Link - Frizzo, L.S., L.P. Sotto, M.V. Zbrun, E. Bertozzi, G. Sequeira, R.R. Armesto and M.R. Rosmini, 2010. Lactic acid bacteria to improve growth performance in young calves fed milk replacer and spray-dried whey powder. Anim. Feed Sci. Technol., 157: 159-167.
CrossRefDirect Link - Fuller, R., 1989. Probiotics in man and animals. J. Applied Bacteriol., 66: 365-378.
CrossRefPubMedDirect Link - Ghorbani, G.R., D.P. Morgavi, K.A. Beauchemin and J.A.Z. Leedle, 2002. Effects of bacterial direct-fed microbials on ruminal fermentation, blood variables and the microbial populations of feedlot cattle. J. Anim. Sci., 80: 1977-1985.
PubMedDirect Link - Gilliland, S.E., 1989. Acidophilus milk products: A review of potential benefits to consumers. J. Diary Sci., 72: 2483-2494.
CrossRefDirect Link - Holter, J.B. and A.J. Young, 1992. Methane prediction in dry and lactating Holstein cows. J. Dairy Sci., 75: 2165-2175.
CrossRefPubMedDirect Link - Klieve, A.V., D. Hennessy, D. Ouwerkerk, R.J. Forster, R.I. Mackie and G.T. Attwood, 2003. Establishing populations of Megasphaera elsdenii YE 34 and Butyrivibrio fibrisolvens YE 44 in the rumen of cattle fed high grain diets. J. Applied Microbiol., 95: 621-630.
CrossRefPubMedDirect Link - Krause, M.K. and G.R. Otzel, 2006. Understanding and preventing subacute ruminal acidosis in dairy herds: A review. Anim. Feed Sci. Technol., 126: 215-236.
CrossRefDirect Link - Krehbiel, C.R., S.R. Rust, G. Zhang and S.E. Gilliland, 2003. Bacterial direct-fed Microbials in ruminant diets: Performance response and mode of action. J. Anim. Sci., 81: E120-E132.
Direct Link - Kritas, S.K., A. Govaris, G. Christodoulopoulos and A.R. Burriel, 2006. Effect of Bacillus licheniformis and Bacillus subtilis supplementation of ewe's feed on sheep milk production and young lamb mortality. J. Vet. Med. Ser. A, 53: 170-173.
CrossRefDirect Link - LeJeune, J.T. and A.N. Wetzel, 2007. Preharvest control of Escherichia coli O157 in cattle. J. Anim. Sci., 85: E73-E80.
PubMedDirect Link - Lynch, H.A. and S.A. Martin, 2002. Effects of Saccharomyces cerevisiae culture and Saccharomyces cerevisiae live cells on in vitro mixed ruminal microorganism fermentation. J. Dairy Sci., 85: 2603-2608.
CrossRefDirect Link - Miron, J., D. Ben-Ghedalia and M. Morrison, 2001. Invited review: Adhesion mechanisms of rumen cellulolytic bacteria. J. Dairy Sci., 84: 1294-1309.
CrossRefDirect Link - Newbold, C.J., R.J. Wallace and F.M. McIntosh, 1996. Mode of action of the yeast Saccharomyces cerevisiae as a feed additive for ruminants. Br. J. Nutr., 76: 249-261.
CrossRefPubMedDirect Link - Nocek, J.E., W.P. Kautz, J.A.Z. Leedle and E. Block, 2003. Direct-fed microbial supplementation on the performance of dairy cattle during the transition period. J. Dairy Sci., 86: 331-335.
CrossRefPubMedDirect Link - Nocek, J.E. and W.P. Kautz, 2006. Direct-fed microbial supplementation on ruminal digestion, health, and performance of pre-and postpartum dairy cattle. J. Dairy Sci., 89: 260-266.
CrossRefPubMedDirect Link - Oetzel, G.R., K.M. Emery, W.P. Kautz and J.E. Nocek, 2007. Direct-fed microbial supplementation and health and performance of pre- and postpartum dairy cattle: A field trial. J. Dairy Sci., 90: 2058-2068.
CrossRefDirect Link - Ohashi, Y. and U. Ushida, 2009. Health-beneficial effects of probiotics: Its mode of action. Anim. Sci. J., 80: 361-371.
CrossRefDirect Link - Puniya, A.K., A.Z.M. Salem, S. Kumar, S.S. Dagar and G.W. Griffith et al., 2015. Role of live microbial feed supplements with reference to anaerobic fungi in ruminant productivity: A review. J. Integr. Agric., 14: 550-560.
CrossRefDirect Link - Quigley, III J.D., L.B. Wallis, H.H. Dowlen and R.N. Heitmann, 1992. Sodium bicarbonate and yeast culture effects on ruminal fermentation, growth and intake in dairy calves. J. Dairy Sci., 75: 3531-3538.
CrossRefPubMedDirect Link - Raeth-Knight, M.L., J.G. Linn and H.G. Jung, 2007. Effect of direct-fed microbials on performance, diet digestibility and rumen characteristics of holstein dairy cows. J. Dairy Sci., 90: 1802-1809.
CrossRefDirect Link - Schamberger, G.P., R.L. Phillips, J.L. Jacobs and F. Diez-Gonzalez, 2004. Reduction of Escherichia coli O157:H7 populations in cattle by addition of colicin E7-producing E. coli to feed. Applied Environ. Microbiol., 70: 6053-6060.
Direct Link - Schmidt, R.J., W. Hu, J.A. Mills and L. Kung, Jr., 2009. The development of lactic acid bacteria and Lactobacillus buchneri and their effects on the fermentation of alfalfa silage. J. Dairy Sci., 92: 5005-5010.
CrossRefDirect Link - Servin, A.L., 2004. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol. Rev., 28: 405-440.
CrossRefPubMedDirect Link - Stein, D.R., D.T. Allen, E.B. Perry, J.C. Bruner and K.W. Gates et al., 2006. Effects of feeding propionibacteria to dairy cows on milk yield, milk components and reproduction. J. Dairy Sci., 89: 111-125.
CrossRefPubMedDirect Link - Wohlt, J.E., A.D. Finkelstein and C.H. Chung, 1991. Yeast culture to improve intake, nutrient digestibility and performance by dairy cattle during early lactation. J. Dairy Sci., 74: 1395-1400.
CrossRefDirect Link - Wohlt, J.E., T.T. Corcione and P.K. Zajac, 1998. Effect of yeast on feed intake and performance of cows fed diets based on corn silage during early lactation. J. Dairy Sci., 81: 1345-1352.
CrossRefPubMedDirect Link - Apas, A.L., M.E. Arena, S. Colombo and S.N. Gonzalez, 2015. Probiotic administration modifies the milk fatty acid profile, intestinal morphology and intestinal fatty acid profile of goats. J. Dairy Sci., 98: 47-54.
CrossRefDirect Link - Bayatkouhsar, J., A.M. Tahmasebi, A.A. Naserian, R.R. Mokarram and R. Valizadeh, 2013. Effects of supplementation of lactic acid bacteria on growth performance, blood metabolites and fecal coliform and lactobacilli of young dairy calves. Anim. Feed Sci. Technol., 186: 1-11.
CrossRefDirect Link - Boyd, J., J.W. West and J.K. Bernard, 2011. Effects of the addition of direct-fed microbials and glycerol to the diet of lactating dairy cows on milk yield and apparent efficiency of yield. J. Dairy Sci., 94: 4616-4622.
CrossRefDirect Link - Brashears, M.M., A. Amezquita and D. Jaroni, 2005. Lactic acid bacteria and their uses in animal feeding to improve food safety. Adv. Food Nutr. Res., 50: 1-31.
CrossRefDirect Link - Brisbin, J.T., H. Zhou, J. Gong, P. Sabour and M.R. Akbari et al., 2008. Gene expression profiling of chicken lymphoid cells after treatment with Lactobacillus acidophilus cellular components. Dev. Comp. Immunol., 32: 563-574.
CrossRefDirect Link - Carlsson, J., Y. Iwami and T. Yamada, 1983. Hydrogen peroxide excretion by oral streptococci and effect of lactoperoxidase-thiocyanate-hydrogen peroxide. Infect. Immunity, 40: 70-80.
Direct Link - Chiquette, J., G. Talbot, F. Markwell, N. Nili and R.J. Forster, 2007. Repeated ruminal dosing of Ruminococcus flavefaciens NJ along with a probiotic mixture in forage or concentrate-fed dairy cows: Effect on ruminal fermentation, cellulolytic populations and in sacco digestibility. Can. J. Anim. Sci., 87: 237-249.
CrossRefDirect Link - Dhama, K., K. Karthik, R. Tiwari, M.Z. Shabbir, S. Barbuddhe, S.V.S. Malik and R.K. Singh, 2015. Listeriosis in animals, its public health significance (food-borne zoonosis) and advances in diagnosis and control: A comprehensive review. Vet. Q., 35: 211-235.
CrossRefDirect Link - Dutta, T.K., S.S. Kundu and M. Kumar, 2009. Potential of direct-fed-microbials on lactation performance in ruminants-a critical review. Livestock Res. Rural Dev., Vol. 21.
Direct Link - Eckles, C.H. and V.M. Williams, 1925. Yeast as a supplementary feed for lactating cows. J. Dairy Sci., 8: 89-93.
CrossRefDirect Link - Fon, F.N. and I.V. Nsahlai, 2013. Effect of direct-fed microbial consortia on ruminal fermentation of maize stover in sheep. Small Rumin. Res., 111: 71-75.
CrossRefDirect Link - Galyean, M.L., G.A. Nunnery, P.J. Defoor, G.B. Salyer and C.H. Parsons, 2000. Effects of live cultures of Lactobacillus acidophilus (strains 45 and 51) and Propionibacterium freudenreichii PF-24 on performance and carcass characteristics of finishing beef steers. Burnett Center Progress Report No. 8, November, 2000, Lubbock, Texas, USA.
- Gulbe, G., A. Valdovska, V. Saulite and J. Jermolajevs, 2015. In vitro assessment for antimicrobial activity of Lactobacillus helveticus and its natural glycopeptides against mastitis causing pathogens in dairy cattle. Open Biotechnol. J., 9: 61-66.
CrossRefDirect Link - Gupta, S. and R.K. Gupta, 2007. Therapeutic efficacy of probiotics during indigestion in cattle. Intas Polivet, 8: 205-207.
Direct Link - Haghighi, H.R., M.F. Abdul-Careem, R.A. Dara, J.R. Chambers and S. Shariff, 2008. Cytokine gene expression in chicken cecal tonsils following treatment with probiotics and Salmonella infection. Vet. Microbiol., 126: 225-233.
CrossRefDirect Link - Jeyanathan, J., C. Martin and D.P. Morgavi, 2014. The use of direct-fed microbials for mitigation of ruminant methane emissions: A review. Animal, 8: 250-261.
CrossRefPubMedDirect Link - Jouany, J.P., F. Mathieu, J. Senaud, J. Bohatier, G. Bertin and M. Mercier, 1999. Influence of protozoa and fungal additives on ruminal pH and redox potential. S. Afr. J. Anim. Sci., 29: 65-66.
Direct Link - Kung, Jr. L., 2006. Direct-Fed Microbial and Enzyme Feed Additives. In: Direct-Fed Microbial, Enzyme and Forage Additive Compendium, Lundeen, T. (Ed.). Miller Publishing, Minnetonka, MN., USA.
Direct Link - Maragkoudakis, P.A., K.C. Mountzouris, C. Rosu, G. Zoumpopoulou and K. Papadimitriou et al., 2010. Feed supplementation of Lactobacillusplantarum PCA 236 modulates gut microbiota and milk fatty acid composition in dairy goats-a preliminary study. Int. J. Food Microbiol., 141: S109-S116.
CrossRefPubMedDirect Link - Matsuguchi, T., A. Takagi, T. Matsuzaki, M. Nagaoka, K. Ishikawa, T. Yokokura and Y. Yoshikai, 2003. Lipoteichoic acids from Lactobacillus strains elicit strong tumor necrosis factor alpha-inducing activities in macrophages through toll-like receptor 2. Clin. Diagn. Lab. Immunol., 10: 259-266.
CrossRefPubMedDirect Link - McAllister, T.A., K.A. Beauchemin, A.Y. Alazzeh, J. Baah, R.M. Teather and K. Stanford, 2011. Review: The use of direct fed microbials to mitigate pathogens and enhance production in cattle. Can. J. Anim. Sci., 91: 193-211.
CrossRefDirect Link - Ohya, T., T. Marubashi and H. Ito, 2000. Significance of fecal volatile fatty acids in shedding of Escherichia coli O157 from calves: Experimental infection and preliminary use of a probiotic product. J. Vet. Med. Sci., 62: 1151-1155.
CrossRefPubMedDirect Link - Orskov, E.R., H.S. Soliman and A. MacDearmid, 1978. Intake of hay by cattle given supplements of barley subjected to various forms of physical treatment or treatment with alkali. J. Agric. Sci., 90: 611-615.
CrossRefDirect Link - Otero, M.C., L. Morelli and M.E. Nader-Macias, 2006. Probiotic properties of vaginal lactic acid bacteria to prevent metritis in cattle. Lett. Applied Microbiol., 43: 91-97.
CrossRefDirect Link - Pagnini, C., R. Saeed, G. Bamias, K.O. Arseneau, T.T. Pizarro and F. Cominelli, 2010. Probiotics promote gut health through stimulation of epithelial innate immunity. Proc. Natl. Acad. Sci. USA., 107: 454-459.
CrossRefDirect Link - Paul, S.S., S.M. Deb and D. Singh, 2011. Isolation and characterization of novel sulphate-reducing Fusobacterium sp. and their effects on in vitro methane emission and digestion of wheat straw by rumen fluid from Indian riverine buffaloes. Anim. Feed Sci. Technol., 166-167: 132-140.
CrossRefDirect Link - Poppi, L.B., J.D. Rivaldi, T.S. Coutinho, C.S. Astolfi-Ferreira, A.J.P. Ferreira and I.M. Mancilha, 2015. Effect of Lactobacillus sp. isolates supernatant on Escherichia coli O157:H7 enhances the role of organic acids production as a factor for pathogen control. Pesquisa Veterinaria Brasileira, 35: 353-359.
CrossRefDirect Link - Renz, F. and A. Koch, 1956. Milk production with the active yeast concentrate Astoral. Zuchtungskunde, 28: 298-301.
Direct Link - Rigobelo, E.E.C., N. Karapetkov, S.A. Maesta, F.A. Avila and D. McIntosh, 2014. Use of probiotics to reduce faecal shedding of Shiga toxin-producing Escherichia coli in sheep. Beneficial Microbes, 6: 53-60.
CrossRefDirect Link - Russell, J.B. and R.L. Baldwin, 1978. Substrate preferences in rumen bacteria: Evidence of catabolite regulatory mechanisms. Applied Environ. Microbiol., 36: 319-329.
PubMedDirect Link - Sandine, W.E., 1979. Roles of Lactobacillus in the intestinal tract. J. Food Protect., 42: 259-262.
Direct Link - Seo, J.K., S.W. Kim, M.H. Kim, S.D. Upadhaya, D.K. Kam and J.K. Ha, 2010. Direct-fed microbials for ruminant animals. Asian-Aust. J. Anim. Sci., 23: 1657-1667.
CrossRefDirect Link - Uyeno, Y., S. Shigemori and T. Shimosato, 2015. Effect of probiotics/prebiotics on cattle health and productivity. Microbes Environ., 30: 126-132.
CrossRefDirect Link - Varankovich, N.V., M.T. Nickerson and D.R. Korber, 2015. Probiotic-based strategies for therapeutic and prophylactic use against multiple gastrointestinal diseases. Front Microbiol., Vol. 6.
CrossRefDirect Link - Wallnofer, P. and R.L. Baldwin, 1967. Pathway of propionate formation in Bacteroides ruminicola. J. Bacteriol., 93: 504-505.
Direct Link - Williams, P.E., C.A. Tait, G.M. Innes and C.J. Newbold, 1991. Effects of the inclusion of yeast culture (Saccharomyces cerevisiae plus growth medium) in the diet of dairy cows on milk yield and forage degradation and fermentation patterns in the rumen of steers. J. Anim. Sci., 69: 3016-3026.
PubMedDirect Link - Yadav, R. and P. Shukla, 2015. An overview of advanced technologies for selection of probiotics and their expediency: A review. Crit. Rev. Food Sci. Nutr., (In Press).
CrossRefDirect Link - Yasuda, K., S. Hashikawa, H. Sakamoto, Y. Tomita, S. Shibata and T. Fukata, 2007. A new synbiotic consisting of Lactobacillus casei subsp. casei and dextran improves milk production in Holstein dairy cows. J. Vet. Med. Sci., 69: 205-208.
CrossRefDirect Link - Yeoman, C.J. and B.A. White, 2014. Gastrointestinal tract microbiota and probiotics in production animals. Annu. Rev. Anim. Biosci., 2: 469-486.
CrossRefDirect Link - Yoon, I.K. and M.D. Stern, 1995. Influence of direct-fed microbials on ruminal microbial fermentation and performance of ruminants-A review. Asian-Aust. J. Anim. Sci., 8: 533-555.
CrossRefDirect Link - Williams, P.E.V., 1989. The Mode of Action of Yeast Culture in Ruminant Diets: A Review of the Effect on Rumen Fermentation Patterns. In: Biotechnology in the Feed Industry: Proceedings of Alltech's Fourth Annual Symposium, Lyons, T.P. (Ed.). Alltech Technical Publications, Nicholasville, USA., pp: 65-84.
- Yuan, C.L., Z.Y. Yu, Y.T. Lin and L.H. Wang, 2015. Effects of probiotics on the calves and its affecting factors. China Dairy Cattle, 8: 9-12.
Direct Link - Aikman, P.C., P.H. Henning, A.K. Jones, S. Potteron and J. Siviter et al., 2008. Effect of administration of Megasphaera elsdenii NCIMB 41125 lactate utilizing bacteria in early lactation on the production, health and rumen environment of highly productive dairy cows fed a high concentrate diet. KK Animal Nutrition Internal Report, KKAN, Umbogintwini, South Africa.
- Huck, G.L., K.K. Kreikemeier and K.K. Bolsen, 1999. Effect of reconstituting field-dried and early-harvested sorghum grain on the ensiling characteristics of the grain and on growth performance and carcass merit of feedlot heifers. J. Anim. Sci., 77: 1074-1081.
PubMedDirect Link - Chaucheyras, F., G. Fonty, G. Bertin and P. Gouet, 1995. Effects of live Saccharomyces cerevisiae cells on zoospore germination, growth and cellulolytic activity of the rumen anaerobic fungus, Neocallimastix frontalis MCH3. Curr. Microbiol., 31: 201-205.
CrossRefDirect Link - Nocek, J.E., W.P. Kautz, J.A.Z. Leedle and J.G. Allman, 2002. Ruminal supplementation of direct-fed microbials on diurnal pH variation and in situ digestion in dairy cattle. J. Dairy Sci., 85: 429-433.
CrossRefDirect Link - Wiryawan, K.G. and J.D. Brooker, 1995. Probiotic control of lactate accumulation in acutely grain-fed sheep. Aust. J. Agric. Res., 46: 1555-1568.
CrossRefDirect Link - Barton, M.D., 2000. Antibiotic use in animal feed and its impact on human health. Nutr. Res. Rev., 13: 279-299.
CrossRefDirect Link