
Qi-Ling Tang
Department of Pharmacy, Yantai Yuhuangding Hospital, No. 20 East of Yuhuangding Road, Zhifu District, Yantai, Shandong 264000, China
Ai-Rong Kang
Department of Respiratory, Yantai Yuhuangding Hospital, Yantai 264000, China
Cong-Xiao Lu
Department of Pharmacy, Yantai Yuhuangding Hospital, No. 20 East of Yuhuangding Road, Zhifu District, Yantai, Shandong 264000, China
International Journal of Pharmacology
Year: 2016 | Volume: 12 | Issue: 2 | Page No.: 116-125







ABSTRACT
The objective of the current study was to evaluate the phytochemical composition and antibacterial effect of methanolic extract of Scutellaria barbata (S. barbata) against a panel of clinically important bacterial pathogens along with deciphering the mode of action of the extract. Liquid chromatography-mass spectrometry was performed to evaluate the phytochemical constituents of the bioactive extract. Disc diffusion assay and microdilution assay were used to test the antibacterial efficacy of the extract. Scanning Electron Microscopy (SEM) was used to evaluate the morphological changes induced by the extract. The 2′, 7′-dichlorofluorescein diacetate (DCFDA) was used to detect the Reactive Oxygen Species (ROS) generation in these six bacterial strains induced by the methanol extract of S. barbata. The extract inhibited cell growth of all tested bacteria dose-dependently with S. typhimurium being the most susceptible pathogen. The percentage of bacterial cells killed within a 40-min contact time between the bacterial cells and the extract was around 24.7% which increased to 44.5% after 60-min time. The extract also induced potent ROS generation as well as intracellular protein leakage in all bacterial cultures with maximum peak in S. typhimurium. The SEM results revealed that the extract induces deleterious morphological changes in the bacterial cell membrane of S. typhimurium indicating that ROS generation can lead to membrane damage in bacterial cells. The present findings indicate that S. barbata has potential antibacterial effect against a variety of bacterial strains and the antibacterial effect is mediated via ROS generation, intracellular protein leakage and rupture of bacterial cell membrane.
PDF Abstract XML References Citation
How to cite this article
Qi-Ling Tang, Ai-Rong Kang and Cong-Xiao Lu, 2016. Phytochemical Analysis, Antibacterial Activity and Mode of Action of the Methanolic Extract of Scutellaria barbata Against Various Clinically Important Bacterial Pathogens. International Journal of Pharmacology, 12: 116-125.
DOI: 10.3923/ijp.2016.116.125
URL: https://scialert.net/abstract/?doi=ijp.2016.116.125
DOI: 10.3923/ijp.2016.116.125
URL: https://scialert.net/abstract/?doi=ijp.2016.116.125
INTRODUCTION
Infectious diseases caused due to bacteria and fungi are the main cause of mortality in tropical and subtropical countries. As per the World Health Organization (WHO) records over 50% deaths are due to infectious diseases in these parts (Kuete et al., 2011). It is believed that due to the indiscriminate use of various antibiotic drugs particularly those of synthetic origin, the pathogenic microbes have developed multidrug resistance which creates serious health issues particularly in developing countries. Many times, this multidrug resistance can result in serious epidemic because no drug can have any effect on the microbes (Weiser et al., 1968). This rapid development of multidrug resistant bacterial and fungal pathogens paves the way for scientists to develop and design novel, newer broad spectrum antimicrobial drugs with less chances of acquiring drug resistance. The less accessibility and high price of new generation antibiotics demands looking for the substances from alternative medicines with proved antimicrobial activity. A large number of medicinal plants have been reported to possess antimicrobial activity especially against drug resistant microbes (Balandrin et al., 1985; Satish et al., 1999; Jones, 1996). Scutellaria barbata D. Don (S. barbata) is broadlyused in traditional Chinese medicine (named Ban-Zhi-Lian in Chinese), which mainly grows throughout Southern China. It contains a large number of alkaloids, flavones, steroids and polysaccharides (Qiao et al., 2011; Liu et al., 2011) due to which it has been found as a potential herb against numerous diseases. (Dai et al., 2013). The biologically active components of Scutellaria barbata are Scutellarin baicalin, luteolin and apigenin. Studies have shown that scutellarin and baicalin were associated with antihypertrophic effects, antioxidant, antitumor, antiviral, inhibit the proteasome and anti-inflammatory activities (Wu et al., 2013; Yang et al., 2011). So, have tried to evaluate this herb against different microbial strains in order to determine its effects. It is estimated that plants have provided the models for 50% of Western drugs (Ncube et al., 2008). Many commercially proven drugs used in modern medicine were initially used in crude form in traditional practices, or for other purposes that suggested potentially useful biological activity. The primary benefits of using plant-derived medicines are that they are relatively safer than synthetic alternatives, offering profound therapeutic benefits and more affordable treatment. The main bioactive chemical compounds present in medicinal plant extracts include phenolics and polyphenols, simple phenols, phenolic acids, quinones, flavones, flavonoids, flavonols, tannins, coumarins, terpenoids, essential oils, lectins, polypeptides, alkaloids etc (Dorman and Deans, 2000).
The objective of the present research work was to evaluate the antibacterial activity of the methanolic extract of the root of Scutellaria barbata against five clinically important bacterial strains along with elucidating the mechanism of action of this extract by studying its effect on cell morphology of Gram negative bacteria Salmonella typhimurium using scanning electron microscopy. It is also demonstrated the effect of the extract on the generation of Reactive Oxygen Species (ROS) in S. typhimurium. Phytochemical analysis of the extract using Liquid Chromatography-Mass Spectrometry (LC-MS) was also done to identify bioactive chemical constituents of the extract.
MATERIALS AND METHODS
Solvents: The HPLC-MS grade acetonitrile were purchased from Merck Co., (Darmstadt, GER). High-purity deionized water was obtained from a Milli-Q water purification system (Millipore Bedford, MA, USA). The HPLC-grade formic acid used for preparation of the mobile phase were from Anaoua chemical supply (Houston, USA), methanol used for plant extraction was from ANPEL Scientific Instrument Co. (Shanghai, CHN).
Plant material and preparation of the extract: The roots of S. barbata were collected during June-July 2014 from Jiuzhaigou, Chengdu, China. The plant material was identified by Prof. Xun-Xhao. The roots were thoroughly washed with tap water, shade dried and then chopped into small pieces. Methanol (95%) was used for hot extraction which was carried out for 4 h using a soxhlet extraction apparatus. The extract was then concentrated under reduced pressure in a rotary evaporator at 40°C and was then kept in a refrigerator at 4°C prior to use.
Liquid chromatography-tandem mass spectrometry (LC-ESI-MSMS)/HPLC analysis: The HPLC analysis was carried out by aNexera HPLC system (Shimadzu, Japan) with a double-pump (LC-30AD), column oven (CTO 30A) and Autosampler (SIL-30AC). A Chromolith RP-18e column (4.6 mm ID, 50 mm length) (Merck) was used. The mobile phase was a gradient prepared from solvent A (0.1% aqueous formic acid) and solvent B (acetonitrile) and the conditions used for gradient elution were: 0-3 min, 5-18%B; 3-5 min, 18%B; 5-6 min, 18-45%B; 6-7 min, 45-100%B; 7-8 min, 100% B. The separation was conducted at 25 with a flow rate of 0.8 mL min–1. The injection volume was 2 μL.
The LC-MS equipment (LC-MS QqQ-6410B Agilent Technologies) consisted of a chromatographic system (1260 In nity Agilent Technologies) coupled with an Agilent Triple Quad mass spectrometer tted with an ESI source. The MS conditions were the following: MS range 100-1200 Da, MSn spectra were obtained using both positive and negative modes, nebulizer gas 45 Psi, gas temperature 325°C, capillary voltage 4000 V.
Microorganism strains and culture media: Streptococcus pneumonia (ATCC 2730), Streptococcus pyogenes (ATCC 12344), methicillin-resistant Staphylococcus aureus (MRSA) (ATCC 43300), Bacillus subtilis (ATCC 6633), Pseudomonas aeruginosa (ATCC 25619) and Salmonella typhimurium (ATCC 10535) were used in the current study. The bacterialstrains were procured from the State Key Laboratory of Microbial Resources (SKLMR), the institute of microbiology, Chinese academy of Sciences, China. Bacterial strains were grown on nutrient agar plates at 37°C and maintained on nutrient agar slants. Cell suspension of microorganisms in 0.5% NaCl was adjusted at 0.5 Mcfarland to obtain approximately 105 CFU mL–1.
Preliminary screening for antimicrobial activity: The antimicrobial test was performed by disc diffusion assay as per NCCLS, 1997 (NCCLS., 1997). The nutrient agar plates containing an inoculum size of 106 CFU mL–1 on Saboraud glucose agar plates were used. Previously prepared extract impregnated disc (6 mm in diameter) at the concentrations of 200 μg mL–1 for bacterial strains were placed aseptically on sensitivity plates with appropriate controls. Vancomycin (100 μg mL–1) was used as standard antibacterial. Plates were incubated at 37°C for 30 h. Antibacterial activity wasrecorded by measuring the clean zone of growthinhibition on agar surface around the disc.
Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC): The MIC and MBC tests were carried out by the broth microdilution method (Yu et al., 2004). The extract was dissolved in sterilized physiological saline solution (0.9%) supplementedwith Tween-80 (Sigma) at final concentration of 0.8% (v/v). Serial doubling dilutions of the extract were prepared in a 96-well microtiter plate ranging from 10-0.125%. Each dilution (100 μL) was dispensed into the wells, then inoculated with (100 μL) of the bacterial suspension. The final concentration of each strain was adjusted to 105-106 CFU mL–1. The concentration of the extract at which the bacteria show no visible growth is defined as Minimum Inhibitory Concentration (MIC) while, as the extract concentration at which the bacteria are killed is defined as Minimum Bactericidal Concentration (MBC). All experiments were performed in triplicate.
Reactive Oxygen Species (ROS) generation by the extract: The 2’, 7’-dichlorofluorescein diacetate (DCFDA) was used to detect the Reactive Oxygen Species (ROS) generation in these six bacterial strains induced by the methanol extract of S. barbata (Kye et al., 1999). The concentration of the extract used was 65 μg mL–1 and the number of bacterial cellsused was adjusted to 106 CFU mL–1. After all cultures were incubated at 37°C for 4 h, they were centrifuged at 4°C for 20 min at 300×g and then each supernatant was treated with 50 μM DCFDA for 2 h. The ROS formed in the samples were detected using Fluorescence MultiDetection Reader (BIOTEK, USA) at 485 nm of fluorescence excitation wavelength and 528 nm of emission wavelength.
Scanning Electron Microscopy (SEM) evaluation: The SEM was performed as reported already in the literature (Yenugu et al., 2004; Agizzio et al., 2006). Salmonella typhimurium was grown in MH agar at 37°C was put in a saline solution comprising of 0.2% tween-80. Four different concentrations of the extract (0, 5, 25 and 75 μg mL–1) were prepared and added into this suspension and was incubated at room temperature. After 24 h, the bacterial cells were centrifuged at 10,000xg for 10 min. The bacterial cells were then washed with 0.1 mol L–1 tris-acetate buffer (pH 7.1), fixed in tris-acetate buffer containing 1.5% glutaraldehyde and then freeze-dried. Each of the bacterial culture was observed by SEM (Hitachi, Japan) at magnifications of 10,000x. The bacterial cell suspension in saline with no extract treatment served as a negative control.
Determination of the rate of killing of the tested bacterium using the S.B extract: This assay determining the rate of killing of the S. typhimurium was evaluated as per an already reported method with minor modifications (Odenholt et al., 2001). Tubes containing 10 mL of broth to which the extract had been added at concentrations of 1, 2 and 4 times the MIC were inoculated with a suspension of the test strain, giving a final bacterial count of approximately 5x106 CFU mL–1. The tubes were thereafter incubated at 37°C and the killing rate was determined over a period of 2 h. A 1.5 mL volume of each suspension was withdrawn at time intervals and transferred to 5 mL of nutrient broth recovery medium containing 5% “Tween 80” to neutralize the effect of the antimicrobial compound carry-overs from the test organisms. The suspension was then successively diluted and plated for viable counts. The plates were later incubated at 37°C for 48 h. The control plates contained the test microorganisms without the S.B extract. The remaining bacterial colonies were counted and compared with the counts of the culture control.
Determination of protein leakage in S. typhimurium bacterial cells: Twelve hour old bacterial cells were washed twice with saline by centrifugation at 12,000 rpm for 15 min followed by re-suspension in physiological saline. The bacterial cells were then treated with varying concentrations of the S. barbata extract relative to MIC (1x MIC, 2x MIC and 4x MIC) for 2 h. Each bacterial suspension treated with extract was centrifuged at 10,000 rpm and the supernatant obtained was evaluated for protein estimation using the Bradford assay (Bradford, 1976). The concentration of protein was estimated from the established standard curve obtained using Bovine Serum Albumin (BSA).
Statistical analysis: All the experiments were done in triplicate and the results were expressed as Mean±SD. Two-way analysis ANOVA was used for comparisons. Statistical significance was calculated at p<0.05.
RESULTS AND DISCUSSION
LC-ESI-MSMS analysis/HPLC analysis: The phytochemical analysis of the S. barbata root extract was carried out by LC-ESI-MS as well as HPLC-DAD techniques. The extract was run under both positive and negative ESI-MS conditions and it showed several major and minor ionic species. The total ion MS chromatogram is shown in Fig. 1, while as the identified compounds are shown in Fig. 2. Fragmentation of the major peaks was used for the identi cation of compounds. The identi cation of the chemical compounds was also carried out by comparing the molecular ion peaks along with the MS fragmentation pattern with those of the literature. The five chemical constituents identified were (1) Scutellarin, (2) Luteolin, (3) Apigenin, (4) 4’-Hydroxywogonin, (5) Luteolin-7-O-glucuronide (Fig. 2). These phytochemicals have already been reported previously in this medicinal plant (Yao et al., 2011).
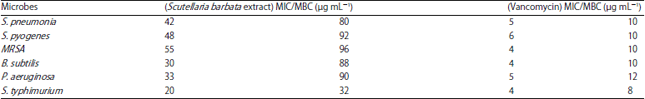
Antibacterial activity of the S. barbata extract: The antibacterial activity of S. barbata extract was evaluated against a panel of six clinically important bacterial pathogens including Streptococcus pneumonia (ATCC 2730), Streptococcus pyogenes (ATCC 12344), methicillin-resistant Staphylococcus aureus (MRSA) (ATCC 43300), Bacillus subtilis ATCC 6633, Pseudomonas aeruginosa (ATCC 25619) and Salmonella typhimurium (ATCC 10535). Both the zones of inhibition as well as MIC/MBC values of the extract were calculated against these microbes using disc diffusion and microdilution assays respectively. The results of the disc diffusion assay (zones of inhibition) are shown in Fig. 3 while, as the results of the microdilution assay (MIC/MBC values) are shown in Table 1. As can be seen from Fig. 3, Salmonella typhimurium exhibited the maximum zone of inhibition at all extract concentrations and as such was most susceptible to the S. barbata extract treatment. Pseudomonas aeruginosa was the next most susceptible bacteria to the extract after S. typhimurium showing higher zones of inhibition at almost all extract doses. Streptococcus pneumonia, Streptococcus pyogenes, methicillin-resistant Staphylococcus aureus bacterial strains were relatively less susceptible to the S. barbata extract. The MIC/MBC values almost also displayed the same trend (Table 1) with S. typhimurium bacterial strain showing the lowest MIC and MBC values indicating that the extract has potent antibacterial effect against this pathogen. MRSA showed the highest value of MIC/MBC indicating that the extract has lesser effect on this bacterial pathogen.
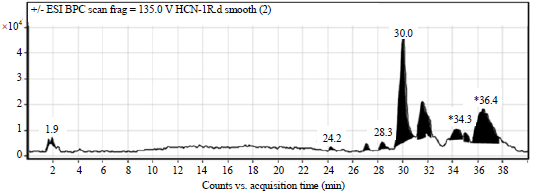 | |
| Fig. 1: | LC-MS chromatogram of the methanol extract of S. barbata. The green numbers show peaks corresponding to compounds, (1) Scutellarin, (2) Luteolin, (3) Apigenin, (4) 4'-Hydroxywogonin and (5) Luteolin-7-O-glucuronide |
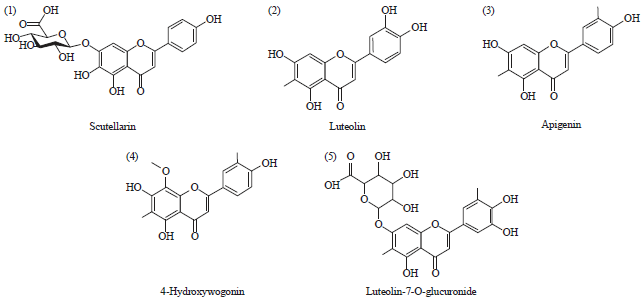 | |
| Fig. 2: | Phytochemical constituents identified in the bioactive extract of Scutellaria barbata using HPLC/LC-MS analytical techniques |
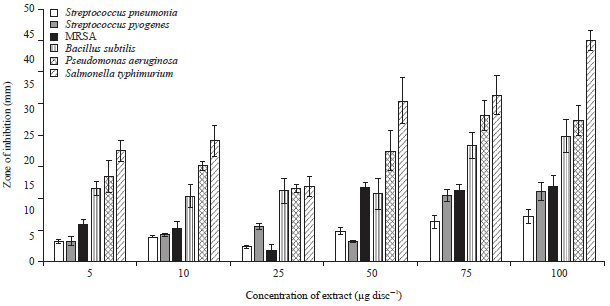 | |
| Fig. 3: | Effect of the different concentrations of the S. barbata extract on the zones of inhibition (antibacterial susceptibility) of the various bacterial strains |
| Table 1: | MIC and MBC values of the S. barbata extract as well as the standard antibacterial drug against a panel of bacterial strains |
 | |
Generation of ROS in bacterial cells treated with different doses of S. barbata extract: There is a direct relation between Reactive Oxygen Species (ROS) generation and bacterialdeath. Various published reports demonstrated that plant extracts could lead to the generation of free radicals within bacterial and fungal cells which finally result in deleterious bacterial membrane damage.
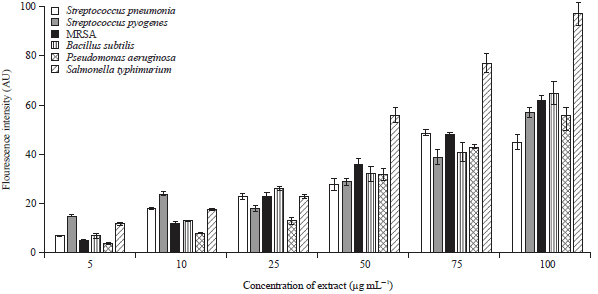 | |
| Fig. 4: | Formation of ROS in S. pneumonia, S. pyogenes, MRSA, B. subtilis, P. aeruginosa and S. typhimurium bacterial cells exposed to different doses of the extract. Bacterial cultures were treated with the extract for 4 h |
In the current study, ROS was measured in six bacterial cultures using DCFDA. After 4 h incubation, elevated levels of ROS were detectedand measured in the S. barbata extract treated bacterial group. As can be seen in Fig. 4, ROS generation was maximum in S. typhimurium at all extract concentrations. This result is consistent with the disc diffusion experiment as well as microdilution assay as both these experiments also showed that S. typhimurium is the most affected bacterial strain. The extract brings about generation of ROS in this bacteria more than other tested bacteria. The extract also induced variable ROS generation in other bacterial strains as can be seen in Fig. 4.
Morphological evaluation of S. typhimurium by scanning electron microscopy after extract treatment: The SEM results indicated that the S. barbata extract targets cell membrane of S. typhimurium bacteria. As can be seen in Fig. 5a-d, S. barbata extract induced potential morphological changes in S. typhimurium bacteria. As compared to the negative control (Fig. 5a, untreated bacterial cells) which showed normal cellular morphology, the S. barbata extract treated bacteria (Fig. 5b-d) showed lot of cellular damage especially at 75 μg mL–1 concentration of extract (D). The membrane damage followed dose-dependent relation with the extract. This membrane damage in S. typhimurium is consistent with the Reactive Oxygen Species generation by the extract. The ROS generation can eventually lead to membrane damage which was demonstrated by SEM in this study.
Extent and rate of killing of S. typhimurium by the S.B extract: The extent and rate of killing of the S. typhimurium by the S.B extract at a concentration of 1x MIC, 2x MIC and 4x MIC is shown in Fig. 6. The percentage of bacterial cells killed within a 40 min contact time between the bacterial cells and the extract was around 24.7% which increased to 44.5% after 60 min time. After 80 min, about 95% of the cells were killed. This behaviour was also seen when the concentration of the S.B extract was doubled, that is 2x MIC. The concentration of the extract was again increased to 4x MIC and the percentage of bacterial cells killed after 40 min contact time was 64.2%. Thus the S.B extract showed a dose-dependent and time-dependent bactericidal effects on S. typhimurium bacterial cells.
Scutellaria barbata extract induced protein leakage in S. typhimurium microbes: Further, the bactericidal effects of the S. barbata extract were evaluated by observing its effect on protein leakage after bacterial cell membrane rupture. At a dose of 1x MIC and 40 min contact time of the bacterial cells with the extract, around 34.2 μg mL–1 of protein leaked out of the S. typhimurium cells which increased to 38.1, 42.4, 56.2 and 58.1 μg mL–1 after 60, 80, 100 and 120 min time periods respectively (Fig. 7). The same pattern of protein leakage was seen when the S. barbata extract concentration was increased to 2x MIC and then 4x MIC.
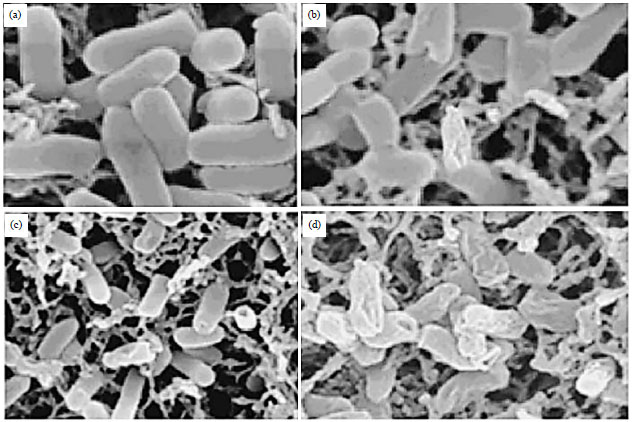 | |
| Fig. 5(a-d): | Electron micrographs of S. typhimurium exposed to different concentrations of the extract. The images were taken at (a) Magnification of 10,000x and S. typhimurium 12 h in saline, (b-d) Scanning electron micrographs of S. typhimurium treated for 12 h with 5, 25 and 75 μg mL–1 of the extract |
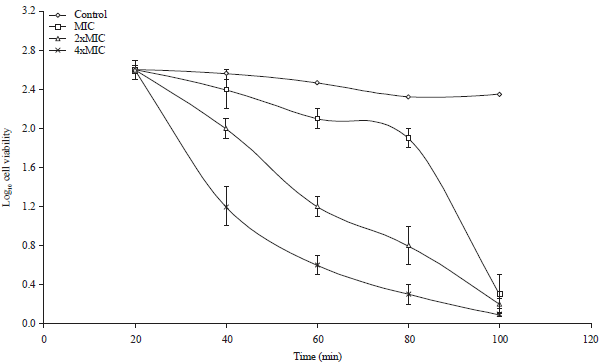 | |
| Fig. 6: | The extent and the rate of killing of S. typhimurium by the S.B extract at concentrations of 1x MIC, 2x MIC, 4x MIC and the control. Each point represents the log10 of the mean survival of bacterial cells at a particular time interval in the presence of the extract |
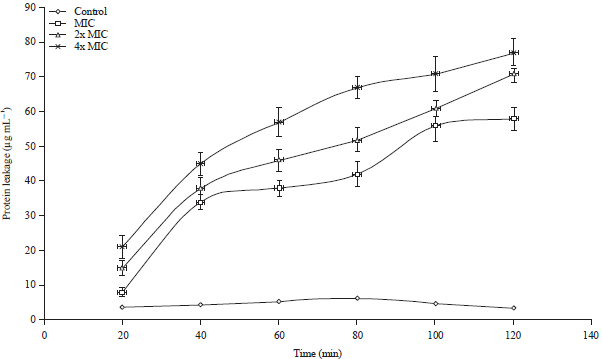 | |
| Fig. 7: | The effect of S. barbata extract on protein leakage from S. typhimurium bacterial cells at concentrations of 1x MIC, 2x MIC, 4x MIC and control. Each point characterizes the amount of protein leaked (μg mL–1) from the bacterial cells at a specific time interval in the presence of the extract |
The protein leakage was much significant at 4x MIC concentration having values of 45.1, 57.3, 67.5, 71.3 and 77.3 μg mL–1 after 40, 60, 80, 100 and 120 min contact of the bacterial cells with the extract, respectively.
DISCUSSION
There are approximately 20,000 plant species being used as ethno-medicines all over the world.However there has been an enormous increase in the multidrug resistant strains of clinically relevant bacterial pathogens which is posing a great challenge to the scientific community. This increase in multidrug resistance is believed to arise due to the indiscriminate use of antibiotics (Srividya et al., 2010; Martin and Yost, 2011; Bibi et al., 2011). Among the available treatments maximum drugs used to treat infectious diseases happen to be secondary metabolites isolated from natural resources. Importantly, these therapeutic secondary metabolites also work as prototypes or lead compounds for the derivation of novel antibiotics. One important feature of plant natural compounds is their hydrophobic nature which render them capable of partitioning the lipid bilayer of the membrane and thus cause disruption of the cell membrane and make them permeable making it vulnerable to the compound effects. Many plant derived compounds including phenols, quinones, terpenoids, flavones have been reported to possess antibacterial activity. Flavonoids and phenols exhibit effective antibacterial activity owing to their ability to form complexes with extra cellular and soluble proteins and to complex with bacterial cell walls leading to the death of the bacteria (Kuete, 2010; Cox et al., 2000; Carson et al., 2002; Cowan, 1999). More specifically flavonoids have been extensively studied and most of the studies have established their role as potential anti-microbial agents (Daglia, 2012; Cushnie and Lamb, 2005; Zhang et al., 2011).
Scutellaria barbata D. Don (Lamiaceae) is a perennial herb which is distributed throughout southern China. This herb is known in traditional Chinese Medicine as Ban-Zhi-Lian. It has been explored for the possible treatment of inflammation, tumor and gynecological diseases in China (Dai et al., 2008). Phytochemically, S. barbata is known to contain alkaloids, polyphenols and flavones (Jiangsu New Medical College, 1977; Qian, 1987). Scutellaria barbata is widely used as an antitumor agent in China. Ethanol extract of S. barbata has been reported to induce apoptosis in the lung cancer cells (A549) while as methylene chloride fraction of the plant has been shown to induce apoptosis in human leukemia cells through the mitochondrial signaling pathway (Yin et al., 2004; Cha et al., 2004). Flavonoids (apigenin and luteolin) were isolated from a 50% ethanolic extract of the plant as active constituents against methicillin-resistant Staphylococcus aureus. The E-1-(40-Hydroxyphenyl)-but-1-en-3-one was isolated from methanol extract of the plant and displayed strong cytotoxicity in K562 human leukaemia cell lines (Sato et al., 2000; Ducki et al., 1996).
S. barbata also contains essential oil with hexahydrofarnesylacetone, 3, 7, 11, 15-tetramethyl-2-hexadecen-1-ol, menthol and 1-octen-3-ol as main chemical constituents. The essential oil has also been reported to possess antibacterial activity against a wide-spectrum of bacterial strains including Staphylococcus aureus, Escherichia coli, Pseudomonas aeruginosa, methicillin-sensitive and methicillin-resistant Staphylococcus aureus, Staphylococcus epidermidis, Staphylococcus heamolyticus, Staphylococcus simulans, Enterococcus faecalis among others (Yu et al., 2004). Keeping in consideration the broad spectrum effects of S. barbata, we have evaluated the antibacterial activity and mode of action of the methanolic extract of the root of S. barbata along with phytochemical analysis of the extract to identify main chemical constituents. Antibacterial efficacy of the extract was measured by studying the zone of inhibition and MIC/MBC values of the extract against different bacterial strains using Disc diffusion and microdilution assays. Scanning Electron Microscopy (SEM) was employed to evaluate the morphological changes induced by the extract in cellular membrane of the bacteria. The extract inhibited growth inhibition of all tested bacteria with S. typhimurium being the most susceptible pathogen towards the extract followed by P. aeruginosa. The extract showed dose-dependent growth inhibitory effects. In addition it was found to induce potent ROS generation in all bacterial cultures with maximum peak in S. typhimurium. The SEM results revealed that the methanolic extract of S. barbata induces deleterious morphological changes in the bacterial cell membrane of S. typhimurium indicating that ROS generation can lead to membrane damage in bacterial cells. The percentage of bacterial cells killed within a 40 min contact time between the bacterial cells and the extract was around 24.7% which increased to 44.5% after 60 min time. After 80 min, about 95% of the cells were killed. This behaviour was evident when the extract concentrations was doubled, that is 2x MIC. Further the intracellular protein leakage was found among the the S. typhimurium bacterial cells when exposed to different doses of the extract indicating bacterial cell wall rupture by the extract and the biocidal effects of the extract on the cells, as evident by the killing rate of test results. Based on the previous studied on S. barbata regarding the potential of this herb as anti-inflammatory, anti-tumor, diuretic and gynecological diseases, it is evident that the herb can a potential lead against different bacterial strains that are clinically important.
CONCLUSION
In conclusion, this study demonstrates that the Scutellaria barbata methanol extract is a potent antimicrobial agent capable of inhibiting the growth of various clinically important microbes. The present study demonstrates that the antibacterial effect of the extract is mediated through membrane damage induced due to ROS generation by the extract followed by the leakage of protein molecules. The extract could be used as a safe and natural alternative to antibiotics with less incidence of showing multidrug resistance.
REFERENCES
- Agizzio, A.P., M. da Cunha, A.O. Carvalho, M.A. Oliveira, S.F.F. Ribeiro and V.M. Gomes, 2006. The antifungal properties of a 2S albumin-homologous protein from passion fruit seeds involve plasma membrane permeabilization and ultrastructural alterations in yeast cells. Plant Sci., 171: 515-522.
CrossRefDirect Link - Balandrin, M.F., J.A. Klocke, E.S. Wurtele and W.H. Bollinger, 1985. Natural plant chemicals: Sources of industrial and medicinal materials. Science, 228: 1154-1160.
CrossRefPubMedDirect Link - Bibi, Y., S. Nisa, F.M. Chaudhary and M. Zia, 2011. Antibacterial activity of some selected medicinal plants of Pakistan. BMC Complementary Altern. Med., Vol. 11.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Carson, C.F., B.J. Mee and T.V. Riley, 2002. Mechanism of action of Melaleuca alternifolia (tea tree) oil on Staphylococcus aureus determined by time-kill, lysis, leakage and salt tolerance assays and electron microscopy. Antimicrob. Agents Chemother., 46: 1914-1920.
CrossRefDirect Link - Cha, Y.Y., E.O. Lee, H.J. Lee, Y.D. Park and S.G. Ko et al., 2004. Methylene chloride fraction of Scutellaria barbata induces apoptosis in human U937 leukemia cells via the mitochondrial signaling pathway. Clinica Chimica Acta, 348: 41-48.
CrossRefDirect Link - Cowan, M.M., 1999. Plant products as antimicrobial agents. Clin. Microbiol. Rev., 12: 564-582.
CrossRefPubMedDirect Link - Cox, S.D., C.M. Mann, J.L. Markham, H.C. Bell, J.E. Gustafson, J.R. Warmington and S.G. Wyllie, 2000. The mode of antimicrobial action of the essential oil of Melaleuca alternifolia (tea tree oil). J. Applied Microbiol., 88: 170-175.
CrossRefPubMedDirect Link - Cushnie, T.P.T. and A.J. Lamb, 2005. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents, 26: 343-356.
CrossRefPubMedDirect Link - Daglia, M., 2012. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol., 23: 174-181.
CrossRefPubMedDirect Link - Dai, Z.J., X.J. Wang, Z.F. Li, Z.Z. Ji and H.T. Ren et al., 2008. Scutellaria barbate extract induces apoptosis of hepatoma H22 cells via the mitochondrial pathway involving caspase-3. World J. Gastroenterol., 14: 7321-7328.
CrossRefDirect Link - Dai, Z.J., B.F. Wang, W.F. Lu, Z.D. Wang and X.B. Ma et al., 2013. Total flavonoids of Scutellaria barbata inhibit invasion of hepatocarcinoma via MMP/TIMP in vitro. Molecules, 18: 934-950.
CrossRefDirect Link - Dorman, H.J.D. and S.G. Deans, 2000. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol., 88: 308-316.
CrossRefDirect Link - Ducki, S., J.A. Hadfield, N.J. Lawrence, C.Y. Liu, A.T. McGown and X. Zhang, 1996. Isolation of E-1-(4'-hydroxyphenyl)-but-1-en-2-one from Scutellaria barbata. Planta Medica, 62: 185-186.
CrossRefPubMedDirect Link - Jones, F.A., 1996. Herbs-useful plants. Their role in history and today. Eur. J. Gastroenterol. Hepatol., 8: 1227-1231.
PubMedDirect Link - Kuete, V., P.Y. Ango, G.W. Fotso, G.D. Kapche and J.P. Dzoyem et al., 2011. Antimicrobial activities of the methanol extract and compounds from Artocarpus communis (Moraceae). BMC Complementary Altern. Med., Vol. 11.
CrossRef - Kuete, V., 2010. Potential of Cameroonian plants and derived products against microbial infections: A review. Planta Medica, 76: 1479-1491.
CrossRefPubMedDirect Link - Liu, G., H. Yang, M. Zhang, S. Li, B. Chen and S. Yao, 2011. Novel isolation of phytochemical compositions by phase transition extraction with acetonitrile. J. Sep. Sci., 34: 347-353.
CrossRefDirect Link - Martin, S.J. and R.J. Yost, 2011. Infectious diseases in the critically ill patients. J. Pharm. Pract., 24: 35-43.
CrossRefDirect Link - Ncube, N.S., A.J. Afolayan and A.I. Okoh, 2008. Assessment techniques of antimicrobial properties of natural compounds of plant origin: Current methods and future trends. Afr. J. Biotechnol., 7: 1797-1806.
CrossRefDirect Link - Odenholt, I., E. Lowdin and O. Cars, 2001. Pharmacodynamics of telithromycin in vitro against respiratory tract pathogens. Antimicrob. Agents Chemother., 45: 23-29.
CrossRefPubMedDirect Link - Qiao, S., R. Shi, M. Liu, C. Liu and W. Yang et al., 2011. Simultaneous quantification of flavonoids and phenolic acids in Herba Scutellariae barbatae and its confused plants by high performance liquid chromatography-tandem mass spectrometry. Food Chem., 129: 1297-1304.
CrossRefDirect Link - Satish, S., K.A. Raveesha and G.R. Janardhana, 1999. Antibacterial activity of plant extracts on phytopathogenic Xanthomonas campestris pathovars. Lett. Applied Microbiol., 28: 145-147.
CrossRefDirect Link - Sato, Y., S. Suzaki, T. Nishikawa, M. Kihara, H. Shibata and T. Higuti, 2000. Phytochemical flavones isolated from Scutellaria barbata and antibacterial activity against methicillin-resistant Staphylococcus aureus. J. Ethnopharmacol., 72: 483-488.
CrossRefDirect Link - Srividya, A.R., S.P. Dhanabal, V.K. Misra and G. Suja, 2010. Antioxidant and antimicrobial activity of Alpinia officinarum. Indian J. Pharmaceut. Sci., 72: 145-148.
CrossRefPubMedDirect Link - Weiser, R., A.W. Asscher and J. Wimpenny, 1968. In vitro reversal of antibiotic resistance by ethylenediamine tetraacetic acid. Nature, 219: 1365-1366.
CrossRefDirect Link - Wu, Y.X., E. Sato, W. Kimura and N. Miura, 2013. Baicalin and scutellarin are proteasome inhibitors that specifically target chymotrypsin-like catalytic activity. Phytother. Res., 27: 1362-1367.
CrossRefPubMedDirect Link - Yang, P., X.Z. Kong, C.M. Cheng, C.Y. Li, X.M. Yang and X.E. Zhao, 2011. Synthesis and biological evaluation of 8-substituted and deglucuronidated scutellarin and baicalin analogues as antioxidant responsive element activators. Sci. China Chem., 54: 1565-1575.
CrossRefDirect Link - Yao, H., S. Li, J. Hu, Y. Chen and L. Huang et al., 2011. Chromatographic fingerprint and quantitative analysis of seven bioactive compounds of Scutellaria barbata. Planta Medica, 77: 388-393.
CrossRefPubMedDirect Link - Yenugu, S., K.G. Hamil, Y. Radhakrishnan, F.S. French and S.H. Hall, 2004. The androgen-regulated epididymal sperm-binding protein, human β-defensin 118 (DEFB118) (formerly ESC42), is an antimicrobial β-defensin. Endocrinology, 145: 3165-3173.
CrossRefPubMedDirect Link - Yin, X., J. Zhou, C. Jie, D. Xing and Y. Zhang, 2004. Anticancer activity and mechanism of Scutellaria barbata extract on human lung cancer cell line A549. Life Sci., 75: 2233-2244.
CrossRefDirect Link - Yu, J., J. Lei, H. Yu, X. Cai and G. Zou, 2004. Chemical composition and antimicrobial activity of the essential oil of Scutellaria barbata. Phytochemistry, 65: 881-884.
CrossRefPubMedDirect Link - Zhang, L., A.S. Ravipati, S.R. Koyyalamudi, S.C. Jeong and N. Reddy et al., 2011. Antioxidant and anti-inflammatory activities of selected medicinal plants containing phenolic and flavonoid compounds. J. Agric. Food Chem., 59: 12361-12367.
CrossRefPubMedDirect Link