
Dante Aviles- Montes
Centro de Investigacion Biomedica del Sur, Instituto Mexicano del Seguro Social (IMSS), Aregentina No. 1, Col. Centro, Xochitepec, 62790, Morelos, Mexico
Maribel Herrera- Ruiz
Centro de Investigacion Biomedica del Sur, Instituto Mexicano del Seguro Social (IMSS), Aregentina No. 1, Col. Centro, Xochitepec, 62790, Morelos, Mexico
Ruben Roman- Ramos
Doctorado en Ciencias Biologicas y de la Salud, Division de Ciencias Biologicas y de la Salud, Universidad Autonoma Metropolitana (UAM), Unidad Iztapalapa, Av. San Rafael Atlixco No. 186, Col. Vicentina, Delegacion Iztapalapa, Distrito Federal, 09340, Mexico
Enrique Jimenez- Ferrer
Centro de Investigacion Biomedica del Sur, Instituto Mexicano del Seguro Social (IMSS), Aregentina No. 1, Col. Centro, Xochitepec, 62790, Morelos, Mexico
Manases Gonzalez- Cortazar
Centro de Investigacion Biomedica del Sur, Instituto Mexicano del Seguro Social (IMSS), Aregentina No. 1, Col. Centro, Xochitepec, 62790, Morelos, Mexico
Alejandro Zamilpa
Centro de Investigacion Biomedica del Sur, Instituto Mexicano del Seguro Social (IMSS), Aregentina No. 1, Col. Centro, Xochitepec, 62790, Morelos, Mexico
Jaime Tortoriello
Centro de Investigacion Biomedica del Sur, Instituto Mexicano del Seguro Social (IMSS), Aregentina No. 1, Col. Centro, Xochitepec, 62790, Morelos, Mexico
International Journal of Pharmacology
Year: 2015 | Volume: 11 | Issue: 8 | Page No.: 944-955







ABSTRACT
Galphimia glauca is a plant species that has been used in mexican traditional Medicine for the treatment of central nervous system disorders. It was demonstrated that the standardized extract of G. glauca, by means of double-blind, controlled and randomized clinical studies, possesses therapeutic effectiveness in patients with generalized anxiety disorder. In the active extract, nor-secotriterpenes Galphimines, Galphimine-A (G-A) and Galphimine-B (G-B) are prominent for their anxiolytic effect. In the present work, the oral administration of different doses of G-A in ICR male mice submitted to the Elevated Plus Maze (EPM) test was capable of inducing a significant anxiolytic effect, while any of the doses modified the spontaneous motor activity of the mice in the Open Field Test (OFT). The administration of G-A (30 mg kg–1, p.o.) in mice inhibited the anxiogenic effect of picrotoxin (PTX, 2 mg kg–1, i.p.), but not of bicuculline (BCC, 5 mg kg–1, i.p.) and pentylenetetrazole (PTZ, 10 mg kg–1, i.p.) in the EPM. The combination of G-A with these drugs was evaluated by means of in vivo extracellular recordings in basolateral amygdala (BLA). A dose-dependent increase of the firing rate on BLA neurons was induced by the administration (i.v.) of different doses of G-A, while diazepam (DZP) decreased the firing rate. The co-administration of G-A with DZP or flumazenil (FMZ) did not modify the effect produced by these drugs on the firing rate. However, the co-administration of G-A modified the effect produced by PTX. The G-A was able to produce an anxiolytic effect when administered at different doses orally in mice, without inducing an associated sedative effect. Electrophysiological recordings showed that the effect produced by G-A is different of that of DZP and suggest that G-A do not interact directly with the GABAergic system (transmission mechanism for the g-aminobutyric acid-GABA), while it was able to modify the effect produced by PTX, BCC and Phaclofen evidencing a different action mechanism.
PDF Abstract XML References Citation
Received: July 13, 2015;
Accepted: October 26, 2015;
Published: November 21, 2015
How to cite this article
Dante Aviles- Montes, Maribel Herrera- Ruiz, Ruben Roman- Ramos, Enrique Jimenez- Ferrer, Manases Gonzalez- Cortazar, Alejandro Zamilpa and Jaime Tortoriello, 2015. Pharmacological Interaction between Galphimine-A, a Natural Anxiolytic Compound and Gabaergic Drugs. International Journal of Pharmacology, 11: 944-955.
DOI: 10.3923/ijp.2015.944.955
URL: https://scialert.net/abstract/?doi=ijp.2015.944.955
DOI: 10.3923/ijp.2015.944.955
URL: https://scialert.net/abstract/?doi=ijp.2015.944.955
INTRODUCTION
In the search for new drugs that could be useful for the treatment of anxiety, medicinal plants are studied as an important source of novel bioactive molecules, Galphimia glauca (Malpighiaceae) (Tortoriello and Ortega, 1993; Herrera-Ruiz et al., 2006a) is a Mexican medicinal species, from which a characteristic group of nor-secotriterpenes, now named galphimines, has been isolated. Highlighted among these are Galphimine-A (G-A) and Galphimine-B (G-B) (Herrera-Ruiz et al., 2006b), to which the anxiolytic effect of this species has been attributed. Recently, the clinical evaluation of a phytopharmaceutical elaborated with the standardized extract (in its galphimine content) of Galphimia glauca was reported. This product, administered during 4 weeks, demonstrated that it possesses therapeutic effectiveness similar to that exerted by Lorazepam in patients with a diagnosis of Generalized Anxiety Disorder (GAD), however, without evidencing important adverse effects (Herrera-Arellano et al., 2007). Another double-blind, controlled clinical study demonstrated that administration of the phytopharmaceutical for 15 weeks in patients with GAD was safe to use and well tolerated (Herrera-Arellano et al., 2012). Studies in animal in vivo tests have also shown that the extract from G. glauca, standardized in its galphimines content is capable of exerting an anxiolytic effect on the EPM test (Herrera-Ruiz et al., 2006a, b).
In an attempt to know the mechanism of action of G-B, it has been evidenced, by means of electrophysiological recordings, that this nor-secotriterpene is capable of modulating serotoninergic transmission in the dorsal hippocampus of anesthetized rats (Jimenez-Ferrer et al., 2011). It was able to diminish the discharge frequency of dopaminergic neurons of the Ventral Tegmental Area (VTA) by means of a mechanism of action independent from the GABAergic system (Tortoriello et al., 1998; Prieto-Gomez et al., 2003).
Anxiety is a normal emotional response to potential threats or stresses and is associated with a variety of psychological disorders when it becomes excessive (Clement and Chapouthier, 1998; Cannistraro and Rauch, 2003). Anxiety disorders are a heterogeneous group of psychiatric disorders with no clear knowledge of their etiology and pathogenesis. Several familial, biological and genetic risk factors have been invoked for the Obsessive Compulsive Disorder (OCD) or the Panic Disorder (PD), however to date none has shown a main role in their etiology (Cavallini and Bellodi, 2001). Anxiety disorders have a high impact on daily life and cause a great deal of suffering for the individual patient.
The amygdala plays an essential role in mediating emotions such as anxiety (Davis et al., 1994; LeDoux, 2003). The primate amygdala is located in the anteromedial part of the temporal lobe, where it lies ventromedial to the striatum and anterior to the ventral portion of the hippocampal formation. It has a similar position in non-primates, such as rat and cat (McDonald, 1998). The basolateral amygdala (BLA) has been widely studied as one of the critical components in the neural circuitry, mediating anxiety-related states and behaviors (Davis, 1998), it is a cortical-like structure consisting of glutamatergic principal neurons and local GABAergic inhibitory neurons (Pape and Pare, 2010). The role of GABAergic cells is particularly important as their inhibitory action provides constant silencing of the main glutamatergic neurons of the BLA (Gaudreau and Pare, 1996). Suppression of inhibition can increase the firing rate and synchronization of glutamatergic neurons during fear (Pare and Collins, 2000).
From the active extract of Galphimia glauca, other galphimines have been isolated and identified, for example, G-E, which is found at a greater concentration in the plant however that does not exhibit activity in anxiety tests. Through a simple chemical reaction, G-E (inactive) is transformed into G-A (active), thus, in this manner it is possible to possess a greater amount of the anxiolytic compound. In the present study, it was evaluated (by means of the EPM test in mice as well as by unitary extracellular recordings in BLA of male rats) the effect produced by different doses of G-A on anxiety and its interaction with potent excitatory substances of the CNS that act on the GABA receptor.
MATERIALS AND METHODS
Plant material: Galphimia glauca was collected from a controlled crop in Morelos state, Mexico. Taxonomic identification was performed by Abigail Aguilar-Contreras, M.Sc., the Herbarium head. A voucher sample was deposited in the Mexican Institute of Social Security Herbarium with code number IMSSM-8646. Plant material was dried under dark conditions at room temperature for 2 weeks. Afterward, in order to obtain particles 3-5 mm in size, the plant material was submitted to an electric grinder mill.
Extract preparation: For obtaining the galphimines, the ground plant material (aerial parts) (7.26 kg) was macerated with n-hexane (16.8 L, 3 times) for its degreasing and later was submitted to extraction with a mixture of ethyl acetate and methanol (50:50, 3 times) (Gonzalez-Cortazar et al., 2005). The extract obtained (310.8 g, 4.27%) was resuspended in solvents with ascending polarity as follows: n-hexane (600 mL, Merck), chloroform (500 mL, Merck), ethyl acetate (500 mL, Merck) and methanol (500 mL, Merck). The solvent was eliminated under conditions of distillation by low pressure in a Heidolph-Laborota 4000 rotary evaporator (Germany), finally obtaining fractions of n-hexane (8 g, 0.01%), ethyl acetate (51.7 g, 0.71%), chloroform (57.4 g, 0.79%) and methanol (189.2 g, 2.6%).
Extraction of galphimine-A (1): The methanolic fraction (150 g), in which the content was mainly the mixture of galphimines was passed through a gravitational chromatographic column (400×90 mm) previously packed with silica gel 60 (600 g, 40-63 μm, Merck). It employed a chloroform-acetone-methanol gradient system as mobile phase, beginning with 100% of the solvent of least polarity and ending with 100% of methanol. This process was monitored by Thin Layer Chromatography (TLC) until obtaining a fraction with the mixture of triterpenes (galphimines). It obtained 45 fractions that, based on the similarity of their content were regrouped into the following eight groups: GgF-A (1-3, 100:0:0), GgF-B (4-19, 100:0:0), GgF-C (20-24, 90:10:0%), GgF-D (25-27, 90:10:0), GgF-E (28-32, 80:20:0), GgF-F (33-36, 70:30:0), GgF-G (37-44, 50:50:0) and GgF-H (45, 0:0:100). The GgF-B fraction (100 g, 66.6%) contains a mixture of galphimines, thus, it was submitted to chemical separation through a gravitational chromatographic column (500 × 50 mm). The solvent system in this case was a gradient composed of n-hexane-ethylacetate-methanol with a gradual increase in polarity, obtaining 48 fractions that were regrouped into the following four groups: GgFB-I (1-8, 70:30:0), GgFB-II (9-11, 70:30:0), GgFB-III (12-16, 50:50:0) and GgFB-IV (17-48, 50:50:10). The GgFB-IV fraction (7.9 g, 7.9%) contained a high amount of Galphimine-E (G-E) and a low concentration of G-A (0.2 g, 1). Due to the high yield of G-E (7.9 g) in this fraction, it was utilized for obtaining G-A by means of a previously reported chemical reaction (Gonzalez-Cortazar et al., 2005) of transformation by hydrolysis. At the end of the reaction, the sample was concentrated at reduced pressure and afterward was taken to complete dryness through lyophilization. The reaction product was obtained (6.55 g, 82.91%), which was separated by gravitational column chromatography (300×20 mm) with silica gel (300 g, 70-230 mesh, Merck). Utilizing a chloroform-acetone-methanol gradient as mobile phase, 50 fractions were obtained, which were regrouped into the following five groups: GARx-1 (1-15, 100:0:0), GARx-2 (16-28, 90:10:0), GARx-3 (29-34, 80:20:0%), GARx-4 (35-49, 70:30:0) and GARx-5 (50, 0:0:100). The GARx-2 fraction was recrystallized with acetone-methanol (7:3 v/v), obtaining crystals (2.8 g, 42.74%) that were identified by NMR of 1H and 13C as Galphimine-A (G-A) (1).
NMR spectra analysis: The spectra of NMR1H and 13C of G-A, dissolved in CD3OD, were obtained in NMR Avance DMX500® Bruker 500 MHz equipment. The chemical displacements were reported in Parts per Million (ppm) and Tetramethylsilane (TMS) was employed as internal reference.
Animals: Male albino ICR mice weighing around 35 g were used (Harlan, Mexico City) for the Elevated plus Maze (EPM) test. Male Sprague-Dawley rats (Harlan, Mexico City) weighing 250-350 g were used for extracellular recordings. All animals were maintained under standard laboratory conditions at 25°C with 12 h light/12 h dark cycles, with lights turned on at 07:00 a.m. and with free access to water and standard food pellets (Harlan rodent diet). The animals were allowed at least 3 weeks to adapt to the laboratory environment prior to initiating the experiments. All experimental procedures were carried out according to a protocol approved by the Institutional Research Committee in compliance with the Official Mexican Regulation that dates from 1999 (NOM-062-ZOO-1999). Minimal number of animals (n = 8) and duration of observation required to obtain consistent data were employed. The experimental protocol was approved by the Local Ethics Committee and was assigned control number R-2010-1701-23.
Drugs: In the experiments, it utilized the following substances: Tween 20 (0.5%) was used as negative control group, G-A (7.5, 15, 30 and 45 mg kg–1, dissolved in Tween 20, 0.5%), GABAergic system modulators, Diazepam (DZP, 1.0 mg kg–1, Pfizer) was employed as an anxiolytic and sedative drug and it is an allosteric modulator of the GABA-A receptor, bicuculline methiodide (BCC, 5 mg kg–1), as GABA-A receptor antagonist, picrotoxin (PTX, 2 mg kg–1), as GABA-A receptor antagonist, binds to the GABA receptor-linked chloride (Cl) channel and is an anxiety inducer and pentylenetetrazole (PTZ, 10 mg kg–1) with non-specific action is a CNS stimulant and induces anxiety. All GABAergic drugs were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Urethane was used as anesthetic drug for the electrophysiological recordings.
Pharmacological design: For in vivo behavioral tests, groups of 8 mice were organized. In order to build dose response curves, different doses (1.0, 1.5, 3.25, 7.5, 15, 30 and 45 mg kg–1) of G-A were administered orally 1 h prior to the initiation of each test (EPM, OFT). For co-administration experiments, all animals were pretreated with 30 mg kg–1 of G-A orally (p.o.) and 30 min later with one of the different GABAergic drugs by intraperitoneal via (i.p.). The doses utilized for each case were as follows: BCC 5 mg kg–1, PTX 2.0 mg kg–1 and PTZ 10 mg kg–1. Finally, 30 min later, each mouse was evaluated in the EPM or OFT tests. The DZP (1.0 mg kg–1 i.p.) and Tween 20 solution (p.o.) were used as a positive (anxiolytic) and negative control group, respectively. For electrophysiological recordings, treatments were administered with increasing and cumulative doses intravenously (i.v.). With the aim to obtain dose-response curves, the following different doses were used: 0.125, 0.25, 0.75, 1.5, 3.0 and 6.0 mg kg–1 for PHC, BCC and PTX; 0.125, 0.25, 0.5, 1.0 and 10 mg kg–1 for DZP and 0.125, 0.75, 1.25, 2.5, 5.0 and 10 mg kg–1 for PTZ. In order to explore the interaction of G-A with different drugs, G-A was administered at 0.125 mg kg–1 and later, the cumulative doses of GABAergic drugs.
Elevated Plus Maze (EPM) test: The EPM test is the most frequently used model for assessment of the anxiolytic activity of a new substance, including herbal remedies (Zhang, 2004) in rodents (Pellow et al., 1985; Lister, 1987). The apparatus was constructed of Plexiglas and consisted of two open arms (30×5 cm) and two closed arms (30×5 cm) with 25 cm walls. The arms extended from a central platform (5×5cm) and the maze was elevated 38.5 cm from the floor of the room. Each animal was placed in the center of the maze, facing one of the closed arms. Number of entries and time on closed and open arms were recorded during a 5 min test. Entry onto an arm was defined as the animal placing all four paws on the arm. All tests were taped with a video camera. After each test, the maze was carefully cleaned with wet tissue paper (10% ethanol solution). Conventional spatial-temporal measurements comprised number of entries (all four paws on open or closed arms and expressed as percentage of total entries on open arms (EOA%) and time on open or closed arms (expressed as percentage of time on open arms (TOA%)).
Open Field Test (OFT): The OFT was employed to evaluate the exploratory activity of the animals (Archer, 1973). Ambulatory behavior was assessed in an Open-Field Test (OFT). The apparatus consisted of transparent acrylic walls and a black floor box measuring 30×30×15 cm in height. The floor of the arena was divided into nine squares equal in area. At the start of each trial, a mouse was placed in the central square and was allowed to explore the arena freely. Number of squares crossed (with all four paws) and number of rearings (R) were counted in a 5 min session. The equipment was cleaned with a solution of 10% ethanol between tests in order to mask animal clues.
In situ extracellular recording in basolateral amygdala (BLA): Rats were anesthetized (i.p.) with urethane (1.5 g kg–1) and were subsequently placed in a stereotaxic device (Stoelting, IL, USA). Body temperature was maintained at 37°C throughout the experiment and prior to recording, a femoral vein was catheterized for i.v. administration of drugs. A midline skin incision was made on the head and burr holes were drilled into the skull overlying the basolateral amygdala (BLA, the recording site). Coordinates for this area were determined utilizing the stereotaxic atlas of (Paxinos and Watson, 1986). The coordinates used for amygdala centered on 4.8-5.5 mm lateral from midline, 2.5-3.8 mm caudal from bregma. The recording electrodes (single-barrel glass) were constructed from borosilicate glass capillary tubing (2 mm outer diameter, World Precision Instruments, Inc., Sarasota, Fl, USA) by using a horizontal flaming Brown micropipette puller (Sutter Instrument Co., CA, USA). The electrode was then filled with 2 M NaCl solution containing Fast green by employing a non-metallic microfill syringe needle (World Precision Instruments, Inc.). Recording began no earlier than 30 min after surgery. Signals from the recording electrode were amplified by a headstage connected to the preamplifier prior to being fed into a window discriminator/amplifier (1000X gain, 100-4000-Hz band pass, Grass PS, Series AC Pre-Amplifier) with low cutoff of 200 Hz and a high cutoff of 16 kHz. Signals were displayed on an oscilloscope (Tektronix, Inc., Beaverton, OR, USA) for real time monitoring, as well as on a custom-designed digital acquisition system (Axon). At the beginning of each experimental trial, we conducted a 5 min baseline recording, after which the appropriate treatments were administered. For the G-A dose-response curve, increasing and cumulative doses were administered; the effect on the neuronal firing frequency of each dose was recorded for 3 min. At the end of recording, a positive current through the recording electrode was applied in order to stain the recording area. Finally, the animals were killed by decapitation and the brain was obtained to verify the recording area. Records concerning the basolateral amygdala (BLA) were considered for pharmacological analysis.
Statistical analysis: Statistical analysis was performed with an SPSS ver. 11.0 software program and based on Analysis of variance (ANOVA) followed by the post-hoc Dunnett test for behavioral tests) and a Student t test was used to analyze the results of the electrophysiological recordings. A significant difference was established with respect to the control group when the p-value was <0.05.
RESULTS AND DISCUSSION
Isolation and identification of Galphimine-A (1): The G-A (1) was obtained in its natural form and by G-E transformation, which also was purified from the Galphimia glauca extract. Corroboration of the structure of G-A was carried out by comparison of the spectroscopic data of the Nuclear Magnetic Resonance (NMR) of 1H and 13C described in the literature (Gonzalez-Cortazar et al., 2005) in addition to comparison with a standard sample of G-A isolated from the plant (Fig. 1).
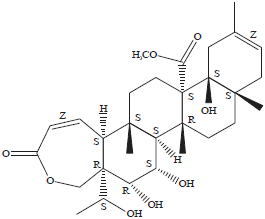 | |
| Fig. 1: | Corroboration of structure of Galphimine-A |
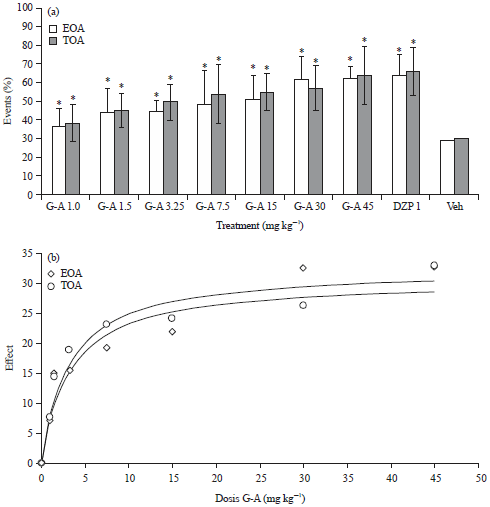 | |
| Fig. 2(a-b): | Effect produced by the oral administration of different doses of Galphimine-A (G-A) (a) On the percentage of entries to open arms (EOA%) and Time spent on open arms (TOA%) of mice in the EPM test and (b) Dose-response curve of Δ of activity of G-A respect to vehicle (G-AEOA-VehEOA). One-way analysis of variance (ANOVA) post-hoc Dunnett test (n = 8, Mean±Standard Deviation (SD)), *p<0.05 (in comparison with the Vehicle (Veh)) |
In vivo behavioral tests
Elevated Plus Maze (EPM) test: Mice that received different doses of G-A by oral pathway (1.0, 1.5, 3.25, 7.5, 15, 30 and 45 mg kg–1) presented a significant (*p<0.05) increment in the percentage of Entries on Open Arms (EOA%) and percentage of time spent on open arms (TOA%) in the EPM test, when compared with the control group (Fig. 2). This effect was dose-dependent and observed even at the lowest dose, the values of EOA and TOA for 1.0 mg kg–1 were 36.48 and 38.25%, while for 45 mg kg–1 were 60.09 and 63.55%, respectively. The Effective dose 50 (ED50) of G-A for the percentage of TOA was 0.44 mg kg–1 and maximal Effect (Emax) was 55.84%, while for percentage of EOA, the ED50 was 0.51 mg kg–1 with Emax = 55.98%. The DZP at 1.0 mg kg–1 induced a significant increment in both parameters (*p<0.05).
In Fig. 3 are shown the results of the administration of GABAergic drugs alone and in combination with G-A. Intraperitoneal (i.p.) administration of PTX (2 mg kg–1) or PTZ (10 mg kg–1) induced significant diminution of percentage of EOA and TOA in comparison with the Vehicle (Veh) (*p<0.05), although BICC (5 mg kg–1) also induced a decrement of these parameters, however, this effect was not different to Veh (p>0.05). The combination of G-A (30 mg kg–1) with BICC or PTZ did not induce any difference respect to the drugs alone and the co-administration of G-A with PTX provoked an increment of percentage of EOA and TOA in comparison with PTX alone (p<0.05) and it was also similar to Veh, although different to the G-A group (p<0.05).
Open Field Test (OFT): The animals treated with DZP showed a diminution in total number of crossings (TC) and rearings (R) during the OFT test, an effect that was significantly different from that of the negative control group (*p<0.05, Veh). Different doses of G-A did not provoke any change of these parameters, in comparison with Veh control group (p>0.05). The GABAergic antagonists PTX and PTZ produced a significant decrement (*p<0.05) of these parameters, however BCC did not induce any change, when they were compared with the negative control group (p>0.05) (Table 1).
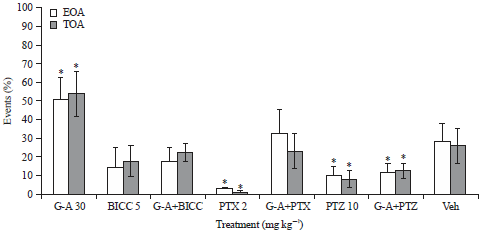 | |
| Fig. 3: | Effect produced by administration of Bicuculline (BICC), Picrotoxin (PTX) and Pentylenetetrazole (PTZ) and each of this drugs in combination with G-A on the percentage of Entries on open arms (EOA%) and Time spent on open arms (TOA%) of ICR mice on the Elevated Plus Maze (EPM) test. One-way Analysis of variance (ANOVA) post-hoc Dunnett test (n = 8, Mean±Standard Deviation (SD)), *p<0.05 in comparison with Vehicle (Veh) |
| Table 1: | Effect produced by oral administration of different doses of Galphimine-A (G-A) and different GABAergic drugs on Total number of Crossings (TC) and on number of vertical Rearings (R) of ICR mice exposed to the open field paradigm |
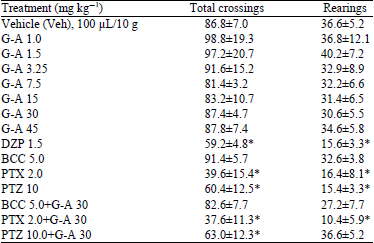 | |
Values represent the Mean±Standard Deviation of the mean. *p<0.05, compared with the Vehicle (Veh): One-way Analysis of variance (ANOVA) post hoc Dunnett test (n = 8), G-A: Galphimine A, DZP: Diazepam, BCC: Bicuculline, PTX: Picrotoxin, PTZ: Pentylenetetrazole | |
Groups of animals that were administered with a GABAA antagonist (BCC, PTX or PTZ) plus G-A (30 mg kg–1) evidenced that G-A (30 mg kg–1) was unable to modify the effect produced by this drugs upon the TC and R parameters, which was similar to that obtained by the administration of these drugs alone (p>0.05) (Table 1).
It has been reported that G-A is one of the characteristic triterpenes of the secondary metabolism of the plant species Galphimia glauca and it has been demonstrated (in pharmacological tests in vivo) that it exerts an anxiolytic effect when administered at a 15 mg kg–1 dose i.p. (Herrera-Ruiz et al., 2006a). In the present work, the anxiolytic effect of this compound was corroborated however, in this case, G-A was administered by the oral route. This allowed affirmation that this substance, highly liposoluble, acts directly on the CNS. It is important to highlight that there are reports of other natural compounds isolated from medicinal plants, such as some flavonoids (quercetin and kaempferol), which exert an anxiolytic effect only when administered orally to mice, although when the i.p. route was used, the effect is not observed. Thus, it is proposed that they act as pro-drugs (Vissiennon et al., 2012). In this case, the response of the mice to oral administration of G-A was very similar to that produced when it was administered i.p., which was reported by Herrera-Ruiz et al. (2006b). In this report, it was indicated that i.p. administration of G-A at a dose of 15 mg kg–1 caused a significant increase in EOA (48%) and TOA (53%) parameters while, in the present work, the oral administration (at the same dose) achieved an increase in EOA of 61% and in TOA of 52%. It can be appreciated that Time spent on open arms (TOA) has remained practically the same; this parameter is the main one employed in EPM validation, in terms of considering a substance as an anxiolytic. During the assay, it was also observed that the effect on the EOA and TOA parameters produced by G-A was dose-dependent. The latter, demonstrates that the margin of activity of G-A on the anxiety disorder (analyzed in mice with the EPM test) is broad and that this range of action did not induce a sedative effect as can be appreciated in the case of the benzodiazepines. The BDZ, in restricted dose ranges, give rise to an anxiolytic effect and even to a sedative and hypnotic effect. These substances are employed experimentally to validate procedures that permit the evaluation of new drugs with potential anxiolytic effect, such as the EPM. This test has sensitivity in the exploration of anxiolytic and anxiogenic drugs (Pellow et al., 1985), whose mechanism of action is utilizing benzodiazepinic receptors. However, the EPM test is also sensitive to substances that possess a non-benzodiazepinic mechanism of action (Pellow and File, 1986). The DZP exerts different biological effects, which are dose-dependent and which range from sedation to anxiolytic and hypnosis (Bousono et al., 2009). In the present study, the 1.0 mg kg–1 dose of DZP administered i.p. produced an increase in the number of EOA and TOA in the EPM model, results that have been widely documented in the literature for this drug.
In order to assess whether the anxiolytic triterpene, G-A, is capable of produce its activity by an interaction of GABAergic system, we decided to measure the behavioral results of mice that received a 30 mg kg–1 dose of G-A co-administered with different substances that represents a useful tool because it induced effects on the CNS by modulating GABA-A receptor, for example BCC (5 mg kg–1), a competitive antagonist; PTX (2.0 mg kg–1), a non-competitive antagonist of GABA-A receptor chloride channels that blocks the anxiolytic-like effects of diazepam; and PTZ (10 mg kg–1), a product with a mechanism of action that is not yet well understood however that represents a useful tool because it induces effects on CNS by modulating GABA-A receptor. All of these compounds have a pro-convulsivant effect and, at low doses, are anxiogenic. In this study was observed the anxiogenic activity of each drug, because all caused a diminution in EOA and TOA parameters in mice submitted to the EPM test. There exist in the literature data that indicate that PTX (0.03-0.5 mg kg–1) and BCC (1-8 mg kg–1) do not possess an anxiogenic effect in the EPM test when administered i.p. (Clenet et al., 2005), it is probable, at least for the case of PTX, that the lack of effect that these authors report is due to the low doses in comparison with that used in the present work (2.0 mg kg–1).
Co-administration of an oral dose of 30 mg kg–1 of G-A with BCC and PTZ did not cause any change of the behavior of mice in comparison with those animals that received these drugs alone. When G-A was in combination with PTX, a diminution of the anxiogenic effect of this drug was observed. At this moment, apparently, G-A seems-not act on binding site of GABA, although it could be capable of interact with the chloride ion channels of GABA-A receptor.
It has been demonstrated that GABAergic drugs, such as the benzodiazepines, exert their depressor effect on the CNS in a dose-dependent manner and that activation of this system gives rise to effects such as anxiolytic, sedation and even hypnosis and death (Bousono et al., 2009). The DZP can exert an anxiolytic plus a sedative effect, considering this as one of its undesirable side effects and one that justifies its use mainly at night. Experimentally, the anxiolytic effect of DZP at different doses i.p. as well as its effect on motor activity have been described and both activities are related according to the dosage increase in the mice (for example, 2.5 and 10 mg kg–1). They also exert an anxiolytic effect, measured as an increase in number of EOA and TOA in the EPM test and a sedative effect, observed as a diminution in locomotion in the OFT (Klodzinska et al., 2004; Clenet et al., 2005). In another assay, administration of 0.05, 0.2, 0.8 and 1.5 mg kg–1 i.p. of DZP to Wistar rats caused diminution of motor activity in OFT (Siemiatkowski et al., 2000). In agreement with these data, in the present work, we observed that the 1.0 mg kg–1 dose of DZP caused an anxiolytic effect accompanied by diminution in spontaneous motor activity in the OFT, which is manifested as a parameter of sedation (Aitkenhead et al., 2007). In this test, the animals were placed in a novel environment and, locomotion, number of rearings (R) and number of groomings (G) among other parameters, were measured, which serve to determine the side or non-specific effects of drugs demonstrating anxiolytic activity in the EPM (Choleris et al., 2001). In contrast with DZP, administration of different doses of G-A did not induce changes in locomotor activity neither in the number of R, indicating, at least in the dose evaluated that this triterpene does not provoke the sedative effects observed with the benzodiazepines. Among the GABAergic drugs, PTX and PTZ caused diminution of motor activity in the OFT, an affect that has been observed in other investigations, for example, administration of subcutaneous doses of 0.6 and 1.0 mg kg–1 to Swiss albino mice induced a diminution of locomotion parameters in this test (Stankevicius et al., 2008). When 30 mg kg–1 of G-A was administered together with the GABAergic antagonists on the OFT, it could be observed that motor activity (TC and R) was not modified respect to animals that received the antagonist drugs alone, which is indicative of no- interaction of G-A with these substances. This test allows us to exclude possible non-specific locomotor effects, of all treatments.
Electrophysiological recordings
Extracellular recording in basolateral amygdala (BLA): A slight dose-dependent increment on the relative index of the firing rate from BLA neurons was induced by the administration (i.v.) of different doses (0.125, 0.25, 0.75, 1.5, 3 and 6 mg kg–1) of G-A (Fig. 4-6) with an Emax = 1.1432 and an ED50 = 0.0150 mg kg–1.
This effect demonstrates that BLA is sensitive to the effects produced by G-A. Which is a brain area involved in the adjustment of mood conditions with a high concentration of glutamatergic neurons and with an important presence of local GABAergic inhibitory neurons (Pape and Pare, 2010). Thus, the increase of firing frequency induced by G-A in BLA could be due to a direct or indirect modulation of the neurotransmission of these endogen messengers. Some experiments for evaluating electrophysiological activity have shown that the chemical and electrical stimulation of neurons in this brain structure is able to alter situations of fear or anxiety. It has been widely observed that the presence of anxiety states is correlated with an increase of neuronal activity at BLA and consequently, higher excitability of BLA has a tendency to be anxiogenic, while lower excitability of this structure will be anxiolytic (Davis et al., 1994). The anxiolytic drug, DZP, produced an inhibition of relative index of firing rate in BLA (Fig. 4a, b) in a dose-dependent manner, which agrees with the anxiolytic activity (Hodges et al., 1987). The effect produced by G-A on this structure was different to that of DZP when the higher doses were used (Hodges et al., 1987).
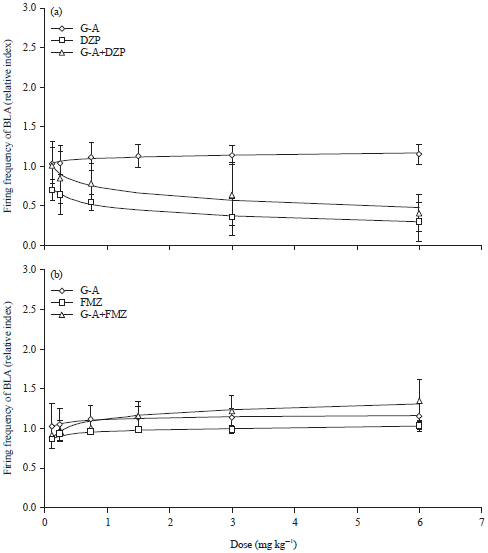 | |
| Fig. 4(a-b): | Effect produced by the co-administration of Galphimine-A (G-A) plus Diazepam (DZP) or Flumazenil (FMZ) on the relativized index of BLA bursting (a) DZP cumulative doses (0.125, 0.25, 0.5, 1.0, 10 mg kg–1), co-administration of G-A (0.125 mg kg–1) plus DZP (0.125, 0.25, 0.5, 1.0 and 10 mg kg–1) and (b) FMZ cumulative doses (0.125, 0.25, 0.75, 1.5, 3, 6 mg kg–1), co-administration of G-A (0.125 mg kg–1) plus FMZ (0.125, 0.25, 0.75, 1.5, 3 and 6 mg kg–1), Student t-test, *p<0.05 |
In this case, G-A produced a slight dose dependent increase of firing rate of BLA. When G-A was co-administered with DZP, the relative index of firing rate was statistically similar to that of the group treated only with DZP (p>0.05). The results indicated that although both substances are able to exert an anxiolytic effect in behavioral tests such as EPM, there could be a difference on the action mechanism.
In an attempt to investigate a possible interaction with benzodiazepinic receptor, it was tested the combination of G-A with Flumazenil (FMZ) (a benzodiazepine receptor antagonist). This drug was administered at increasing doses, exhibiting a slight increase in the neuronal excitability of BLA. Although this effect was not significantly different with G-A group, the results are in agreement with reports in which substances that block GABAergic transmission induce an anxiogenic effect and decrease neuronal firing rate in this structure (Fig. 4b). When G-A was co-administered with FMZ, the firing rate of the dose response curve was increased in comparison with the group treated with FMZ alone (Fig. 4b) and with G-A alone. But, again the effect was no different between these groups (p>0.05). This finding indicates that there is no pharmacological interaction between G-A and drugs that modify neurotransmission with the GABA-A receptor, specifically on the benzodiazepine receptor.
In panel A of Fig. 5, the neuronal firing rate of BLA by administration of PTX (non-competitive GABA-A receptor antagonist) is shown. An increase of firing rate is observed, which is not significantly different from that of animals that received G-A alone. However, co-administration of G-A+PTX produced a significant increase in firing rate, but only at the highest doses (p<0.05). This result produced by G-A, of modifying the effect produced by PTX was also observed in the EPM test, in which the combination of G-A and PTX was able to increase significantly the percentage of TOA and EOA.
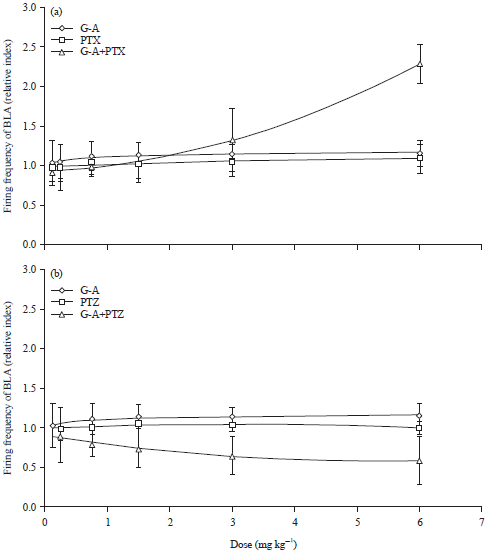 | |
| Fig. 5(a-b): | Effect produced by co-administration of Galphimine-A (G-A) plus picrotoxin (PTX) or pentylenetetrazole (PTZ) on the relativized index of BLA bursting (a) PTX cumulative doses (0.125, 0.25, 0.75, 1.5, 3, 6 mg kg–1), co-administration of G-A (0.125 mg kg–1) plus PTX (0.125, 0.25, 0.75, 1.5, 3, 6 mg kg–1) and (b) PTZ cumulative doses (0.75, 1.25, 2.5, 5.0, 10 and 15 mg kg–1), co-administration of G-A (0.125 mg kg–1) plus PTZ (0.75, 1.25, 2.5, 5.0, 10, 15 mg kg–1), Student t-test, *p<0.05 |
Panel B of the same figure illustrates that administration of increasing doses of PTZ did not modify the frequency of neuronal firing and although the co-administration of G-A with PTX provoked a decrement of firing frequency, there was not a significantly difference (p>0.05).
Panel A of Fig. 6 shows that BCC (a competitive antagonist of GABA receptors) did not modify the frequency of neuronal firing, while the co-administration of G-A+BCC significantly increased this parameter in a dose-dependent manner. Moreover, these curve values were also higher than those of the effect of G-A when it was administered alone (p<0.005).
Phaclofen (PHC, an antagonist of the GABA-B receptor) induced a significant increase of firing rate in the BLA in a dose-dependent manner. This effect could be considered similar to that produced by G-A, however when both substances were co-administered (G-A+PHC), this produced a consistent and dose dependent diminution of the firing rate (Panel B Fig. 6).
Data presented here suggest that the effect produced by G-A in BLA neurons is different of that of DZP and does not interact directly with GABAergic system. The possible interaction between G-A and PTX, that it was observed in the present work, probably do not involve the GABAergic system. It is well known that PTX is also able to inhibit the Cl-influx in Glycine and Glu receptors; as well as to block receptors that are Cation selective, as it is the case of the 5-HT3A (Das et al., 2003). The PTX is also able to antagonize the excitatory effect of DOI (an agonist of the 5-HT2A/2C receptor) in BLA (Sun et al., 2013) in which, the activation produces anxiolytic effect.
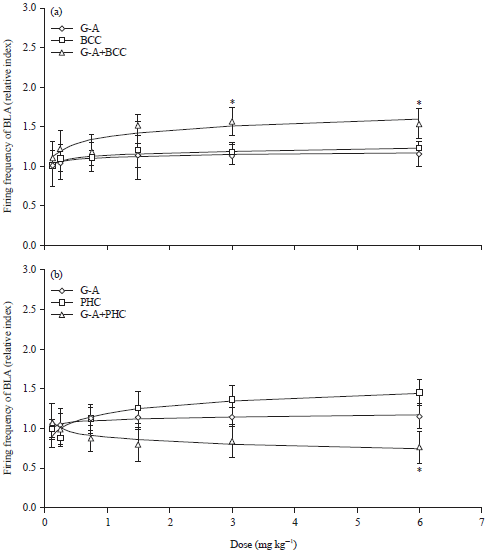 | |
| Fig. 6(a-b): | Effect produced by the co-administration of Galphimine-A (G-A) plus bicuculline (BCC) or phaclofen (PHC) under the relativized index of BLA bursting (a) Cumulative increasing doses of BCC (0.125, 0.25, 0.75, 1.5, 3 and 6 mg kg–1, n), co-administration of G-A (0.125 mg kg–1) plus increasing doses of BCC (p) and (b) Cumulative increasing doses of PHC (0.125, 0.25, 0.75, 1.5, 3 and 6 mg kg–1), co-administration of G-A (0.125 mg kg–1) plus increasing doses of PHC (p), Student t-test, *p<0.05 |
With these facts and due to the structural similarity between G-A and G-B (the other anxiolytic triterpene isolated from G. glauca, that has shown an interaction with the serotoninergic system in hippocampus), it is possible to suggest that the in vivo anxiolytic activity of G-A could involve the interaction with other neurotransmission system as could be the serotoninergic.
CONCLUSION
It was demonstrated that the oral administration of the triterpene G-A, isolated from the medicinal plant species Galphimia glauca, possesses a dose dependent anxiolytic effect, which does not produce an associated sedative effect. Also, it was observed that G-A did not interact with PTZ and BICC in the EPM, although it is interacting with PTX. Results obtained from electrophysiological recordings showed that the effect produced by G-A is different to that of DZP and suggest that G-A do not interact directly with the GABAergic system, while it was able to modify the effect produced by PTX, BCC and PHC, evidencing a different action mechanism.
ACKNOWLEDGMENT
This study was supported by financing from the Coordination of Research in Health of the Mexican Institute of Social Security (FIS/IMSS/PROT/G11/928) and from CONACyT-México (CB-2012-01-181180). We thank the Consejo Nacional de Ciencia y Tecnología (CONACyT) for the grant (206839) for the Doctoral studies of Dante Avilés-Montes at the Universidad Autónoma Metropolitana (UAM)-Iztapalapa. Thanks are also given to the Mexican Institute of Social Security (IMSS) for the grant awarded (99183985) for the Doctoral studies of Dante Avilés-Montes.
REFERENCES
- Archer, J., 1973. Tests for emotionality in rats and mice: A review. Anim. Behav., 21: 205-235.
CrossRefPubMedDirect Link - Cannistraro, P.R. and S.L. Rauch, 2003. Neural circuitry of anxiety: Evidence from structural and functional neuroimaging studies. Psychopharmacol. Bull., 37: 8-25.
PubMedDirect Link - Clement, Y. and G. Chapouthier, 1998. Biological bases of anxiety. Neurosci. Biobehav. Rev., 22: 623-633.
CrossRefDirect Link - Clenet, F., M. Hascoet, G. Fillion, H. Galons and M. Bourin, 2005. Role of GABA-ergic and serotonergic systems in the anxiolytic-like mechanism of action of a 5-HT-moduline antagonist in the mouse elevated plus maze. Behav. Brain Res., 158: 339-348.
CrossRefDirect Link - Choleris, E., A.W. Thomas, M. Kavaliers and F.S. Prato, 2001. A detailed ethological analysis of the mouse open field test: Effects of diazepam, chlordiazepoxide and an extremely low frequency pulsed magnetic field. Neurosci. Biobehav. Rev., 25: 235-260.
CrossRefDirect Link - Das, P., C.L. Bell-Horner, T.K. Machu and G.H. Dillon, 2003. The GABAA receptor antagonist picrotoxin inhibits 5-hydroxytryptamine type 3A receptors. Neuropharmacology, 44: 431-438.
CrossRefDirect Link - Davis, M., 1998. Are different parts of the extended amygdala involved in fear versus anxiety? Biol. Psychiatry, 44: 1239-1247.
CrossRefDirect Link - Davis, M., D. Rainnie and M. Cassell, 1994. Neurotransmission in the rat amygdala related to fear and anxiety. Trends Neurosci., 17: 208-214.
CrossRefDirect Link - Gaudreau, H. and D. Pare, 1996. Projection neurons of the lateral amygdaloid nucleus are virtually silent throughout the sleep-waking cycle. J. Neurophysiol., 75: 1301-1315.
PubMedDirect Link - Gonzalez-Cortazar, M., J. Tortoriello and L. Alvarez, 2005. Norsecofriedelanes as spasmolytics, advances of structure-activity relationships. Planta Med., 71: 711-716.
CrossRefPubMedDirect Link - Herrera-Arellano, A., E. Jimenez-Ferrer, A. Zamilpa, M. Morales-Valdez, C.E. Garcia-Valencia and J. Tortoriello, 2007. Efficacy and tolerability of a standardized herbal product from Galphimia glauca on generalized anxiety disorder. A randomized, double-blind clinical trial controlled with lorazepam. Planta Med., 73: 713-717.
CrossRefPubMedDirect Link - Herrera-Arellano, A., J.E. Jimenez-Ferrer, A. Zamilpa, G. Garcia-Alonso, S. Herrera-Alvarez and J. Tortoriello, 2012. Therapeutic effectiveness of Galphimia glauca vs. lorazepam in generalized anxiety disorder. A controlled 15-week clinical trial. Planta Med., 78: 1529-1535.
CrossRefPubMedDirect Link - Herrera-Ruiz, M., M. Gonzalez-Cortazar, E. Jimenez-Ferrer, A. Zamilpa, L. Alvarez, G. Ramirez and J. Tortoriello, 2006. Anxiolytic effect of natural galphimines from Galphimia glauca and their chemical derivatives. J. Nat. Prod., 69: 59-61.
CrossRefDirect Link - Herrera-Ruiz, M., J.E. Jimenez-Ferrer, T.C.M. de Lima, D. Aviles-Montes, D. Perez-Garcia, M. Gonzalez-Cortazar and J. Tortoriello, 2006. Anxiolytic and antidepressant-like activity of a standardized extract from Galphimia glauca. Phytomedicine, 13: 23-28.
CrossRefDirect Link - Hodges, H., S. Green and B. Glenn, 1987. Evidence that the amygdala is involved in benzodiazepine and serotonergic effects on punished responding but not on discrimination. Psychopharmacology, 92: 491-504.
CrossRefDirect Link - Jimenez-Ferrer, E., M. Herrera-Ruiz, R. Ramirez-Garcia, A. Herrera-Arellano and J. Tortoriello, 2011. Interaction of the natural anxiolytic galphimine-b with serotonergic drugs on dorsal hippocampus in rats. J. Ethnopharmacol., 137: 724-729.
CrossRefDirect Link - Klodzinska, A., E. Tatarczynska, E. Chojnacka-Wojcik, G. Nowak, N.D.P. Cosford and A. Pilc, 2004. Anxiolytic-like effects of mtep, a potent and selective mglu5 receptor agonist does not involve GABAA signaling. Neuropharmacology, 47: 342-350.
CrossRefDirect Link - LeDoux, J., 2003. The emotional brain, fear and the amygdala. Cell. Mol. Neurobiol., 23: 727-738.
CrossRefDirect Link - Lister, R.G., 1987. The use of a plus-maze to measure anxiety in the mouse. Psychopharmacology, 92: 180-185.
CrossRefDirect Link - McDonald, A.J., 1998. Cortical pathways to the mammalian amygdala. Progr. Neurobiol., 55: 257-332.
CrossRefDirect Link - Pape, H.C. and D. Pare, 2010. Plastic synaptic networks of the amygdala for the acquisition, expression and extinction of conditioned fear. Physiol. Rev., 90: 419-463.
CrossRefDirect Link - Pare, D. and D.R. Collins, 2000. Neuronal correlates of fear in the lateral amygdala: Multiple extracellular recordings in conscious cats. J. Neurosci., 20: 2701-2710.
PubMedDirect Link - Pellow, S., P. Chopin, S.E. File and M. Briley, 1985. Validation of open: Closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Methods, 14: 149-167.
CrossRefDirect Link - Pellow, S. and S.E. File, 1986. Anxiolytic and anxiogenic drug effects on exploratory activity in an elevated plus-maze: A novel test of anxiety in the rat. Pharmacol. Biochem. Behav., 24: 525-529.
CrossRefDirect Link - Prieto-Gomez, B., J. Tortoriello, A. Vazquez-Alvarez and C. Reyes-Vazquez, 2003. Galphimine B modulates synaptic transmission on dopaminergic ventral tegmental area neurons. Planta Med., 69: 38-43.
CrossRefPubMedDirect Link - Siemiatkowski, M., H. Sienkiewicz-Jarosz, A.I. CzLonkowska, A. Bidzinski and A. Plaznik, 2000. Effects of buspirone, diazepam and zolpidem on open field behavior and brain [3H]muscimol binding after buspirone pretreatment. Pharmacol. Biochem. Behav., 66: 645-651.
CrossRefDirect Link - Stankevicius, D., E.C. Rodrigues-Costa, J.C. Florio and J. Palermo-Neto, 2008. Neuroendocrine, behavioral and macrophage activity changes induced by picrotoxin effects in mice. Neuropharmacology, 54: 300-308.
CrossRefDirect Link - Sun, Y.N., L.B. Li, Q.J. Zhang, Y.P. Hui and Y. Wang et al., 2013. The response of juxtacellular labeled GABA interneurons in the basolateral amygdaloid nucleus anterior part to 5-HT2A/2C receptor activation is decreased in rats with 6-hydroxydopamine lesions. Neuropharmacology, 73: 404-414.
CrossRefDirect Link - Tortoriello, J. and A. Ortega, 1993. Sedative effect of galphimine B, a nor-seco-triterpenoid from Galphimia glauca. Planta Med., 59: 398-400.
CrossRefPubMedDirect Link - Tortoriello, J., A. Ortega, M. Herrera-Ruiz, J. Trujillo and C. Reyes-Vazquez, 1998. Galphimine-b modifies electrical activity of ventral tegmental area neurons in rats. Planta Med., 64: 309-313.
CrossRefPubMedDirect Link - Vissiennon, C., K. Nieber, O. Kelber and V. Butterweck, 2012. Route of administration determines the anxiolytic activity of the flavonols kaempferol, quercetin and myricetin-are they prodrugs? J. Nutr. Biochem., 23: 733-740.
CrossRefDirect Link - Zhang, Z.J., 2004. Therapeutic effects of herbal extracts and constituents in animal models of psychiatric disorders. Life Sci., 75: 1659-1699.
CrossRefDirect Link