
Ming Yang
Beijing Key Laboratory of TCVM, Department of Veterinary Medicine, Beijing University of Agriculture, 102206, China
Yan Lu
Beijing Key Laboratory of TCVM, Department of Veterinary Medicine, Beijing University of Agriculture, 102206, China
Yuanyuan Ma
Laboratory Animal Center, Peking University, 100034, China
Guoying Wu
Maternity and Child Care Center, Changchun City, Jilin, 130600, China
Ross C. Beier
USDA, Agricultural Research Service, Southern Plains Agricultural Research Center, Food and Feed Safety Research Unit, College Station, Texas, USA
Xiaolin Hou
Beijing Key Laboratory of TCVM, Department of Veterinary Medicine, Beijing University of Agriculture, 102206, China
Guojuan Wu
Beijing Key Laboratory of TCVM, Department of Veterinary Medicine, Beijing University of Agriculture, 102206, China
International Journal of Pharmacology
Year: 2015 | Volume: 11 | Issue: 4 | Page No.: 394-399







ABSTRACT
Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) represents a significant challenge to the swine industry worldwide. Current control strategies against PRRSV are still inadequate and there is a need for new antiviral therapy method. Forsythoside is a compound derived from fruit of Forsythia suspensa (Thunb). Vahl suspensa, a traditional antiviral Chinese medicine. The effect of forsythoside on the infection of Marc-145 cells by PRRSV was investigated through the in vitro antivirus action and cytopathic change assay, It was found that forsythoside had a significant protective effect on Marc-145 cells pathological changes, the protective activity from high to low was treatment prior infection, treatment of infected cells, direct virucidal effect, respectively. Furthermore, its effect on the PRRSV RNA capacity and secretion of IFN-α was also evaluated via measurement of the cytopathic changes, real time PCR and ELISA assay. It indicated that forsythpside A inhibited PRRSV RNA synthesis and promoted secretion of IFN-α and in infected Marc-145 cells at a certain concentrations. The research has prepared the ground for later development of forsythoside as an anti-virus agent.
PDF Abstract XML References Citation
Received: December 10, 2014;
Accepted: February 21, 2015;
Published: April 18, 2015
How to cite this article
Ming Yang, Yan Lu, Yuanyuan Ma, Guoying Wu, Ross C. Beier, Xiaolin Hou and Guojuan Wu, 2015. Inhibition of Porcine Reproductive and Respiratory Syndrome Virus in vitro By Forsythoside A. International Journal of Pharmacology, 11: 394-399.
DOI: 10.3923/ijp.2015.394.399
URL: https://scialert.net/abstract/?doi=ijp.2015.394.399
DOI: 10.3923/ijp.2015.394.399
URL: https://scialert.net/abstract/?doi=ijp.2015.394.399
INTRODUCTION
Porcine Reproductive and Respiratory Syndrome (PRRS) is one of the most important contagious diseases of swine in the worldwide (Chen et al., 2011; Neumann et al., 2005). PRRS virus (PRRSV), the causative agent of PRRS, causes reproductive failures in breeding age swine which are characterized by mummification, stillbirth, late-term abortion and delayed return to estrus. The virus also causes respiratory disorders in growing pigs which can also be subclinical or fatal depending on the virulence of the virus. The PRRSV-infected pigs usually decreased growth performance and are highly susceptible to other pathogenic microorganism co-or secondary infections.
Great efforts have been made to control and eradicate PRRSV infection since it was identified. However, unsatisfactory progress has been made and PRRSV still remains one of the biggest challenges to swine industry. The present management approach to control PRRS mainly depends on vaccines. But, unfortunately, the currently commercialized vaccines only confer limited immunity and protection from heterologous infections and cannot completely eradicate the virus (Murtaugh and Genzow, 2011). Thus, it is an urgent demand for novel strategies to control PRRSV infection.
Antiviral therapeutics is an alternative tool for combating virus infections, especially for virus that does not have vaccines to match well with the circulating virus. Several compounds and compositions have been investigated positive activities against PRRSV (Cheng et al., 2013; Gao et al., 2013; Karuppannan et al., 2012; Wang et al., 2013a; Yang et al., 2013). However, there is a long way for effective commercial drugs available to control PRRSV infection.
The fruit of Forsythia suspensa Vahl (Oleaceae) is widely used as traditional Chinese medicine to treat inflammation, pyrexia, sepsis, bacterial and viral infection. Forsythoside A is one of the active ingredients of Forsythia suspense. And it inhibited pathogenic Staphylococcus aureus, avian infectious bronchitis virus and coxsackie virus in vitro (Li et al., 2011).
The virus-infected cells have been shown to produce various biologically active cytokines and chemokines, including the chemotactic cytokines, pro-inflammatory cytokines (IL-1β and TNF-α) and antiviral cytokines (IFN-α, β) (Ferrari et al., 2013). However, porcine reproductive and respiratory syndrome virus inhibits type I interferon signaling (Patel et al., 2010). IFN-α not only inhibits virus replication but also stimulates immune cells. Thus, induction of IFN-α production has been implicated to produce a beneficial effect on controlling PRRS exacerbation.
Based on the antiviral activities of forsythoside A, we hypothesized that the forsythoside A may be an effective anti-PRRSV ingredient. In this study, the protective activity of forsythoside A against PRRSV infection and regulation effect on PRRSV RNA synthesis and IFN-α production in vitro were examined.
MATERIALS AND METHODS
Reagents, virus and cells: Eagles’s Minimum Essential Medium (MEM) (Sigma, USA) supplemented with 10% heat-inactivated fetal bovine serum (Hyclone, USA), 100 IU mL-1 Penicillin G and 100 μg mL-1. Streptomycin was used for cell growth or maintenance medium. Forsythoside A was purchased from China Pharmaceutical and Biological Identification Institute, was dissolved in pH 7.2 PBS (1 mg mL-1). The solution was sterilized by a 0.22 μm millipore membrane filter and aliquots made for future use and diluted with MEM. Pig IFN-a ELISA kit was produced by Rapidbio with detection limit at 5 pg mL-1 (USA). PRRSV (VR2332, the prototype of Type 2 PRRSV strain was bought from China Institute of Veterinary Drug Control.
Cells culture and virus preparation: Marc-145 cells were diluted to 2×105 cells mL-1 with MEM supplemented with 10% fetal bovine serum and cultured in 24-well plates at 37°C in 5% CO2 atmosphere. Subculture was carried out every 2-3 days after it had formed a confluent monolayer. The PRRSV (VR2332, the prototype of Type 2 PRRSV strain, bought from China Institute of Veterinary Drug Control) was propagated in Marc-145 cells. PRRSV was serially diluted at 10-fold in MEM to infect Marc-145 cells in 24-well plates. The viruses diluted in 10-1, 10-2,…10-8 folds were inoculated to each well (100 μL per well), respectively. Each dilution was repeated eight times. Virus titer was determined by the Reed-Muench assay method and expressed as tissue culture infective dose TCID50 (Quinting et al., 2007).
Cytotoxicity assay of forsythoside A: Marc-145 cells suspension was cultured in 24-well plates at a density of 1×104 cells (0.1 mL) per well at 37°C for 36 h. Marc-145 cells in a 24-well plate were cultured in 100 μL MEM containing a series concentrations of forsythoside A at 37°C. Each concentration set six parallel and repeated three times. Five days later. The cytotoxic effect was evaluated by MTT assay (Wang et al., 2013a). The MTT signal was measured at an absorbance of 550 nm. The maximum non-cytotoxic concentration was calculated as the concentration which conserves the 90% cellular viability.
Real-time reverse-transcription PCR (RT-PCR): The total RNA of Marc-145 cells was extracted using trizol reagent (Invitrogen), RNA was reverse transcripted into cDNA using avian AMV system (Promega). The quantity of extracellular PRRSV RNA was detected by real-time PCR (ABI PRISM 7300 Sequence Detector, PE Biosystems) with primers using specific primers for PRRSV ORF7 (Forward: CAAATAACAACGGCAAGCAG, Reverse: AAACTCCACAGCGTAACTTATC). Real-time Quantification PCR was completed as following: 10 μL of SYBR Green I Master mix, 0.1 μL of 10 pmol μL-1 forward and reverse primers, respectively, 0.2 μL of referencing dye (ROX), 1.5 μL of recombined PRRSV ORF7 pEASY-T3 plasmid DNA templates or cDNA sample, 9.1 μL of water. The reaction mixture was incubated at 94°C for 30 sec, 55ΊC for 30 sec and 72ΊC for 30 sec, for 40 circles. The assay was completed on ABI 7500 Real Time PCR System.
For the quantification of the real samples, PRRSV standard was reversely transcript, PRRSV ORF7 was amplicated using the primers above and linked into pEASY-T3 plasmid. The recombined pEASY-T3 plasmid was transformed and amplicated in Escherichia coli. After extracting, the plasmid was sequenced and conformed to AY262352 in genebank. For each assay, a standard curve was generated using serially diluted pEASY-T3 plasmid. The sample RNA copies in all samples were calculated by comparing them with it.
Treatment of Marc-145 cells prior to virus infection: Marc-145 cells in 24-well plates were treated with forsythoside A (10, 20, 40, 60 μg mL) for 24 h. Four replicates were carried for each concentration. Then the Marc-145 cells were incubated with PRRSV (100 TCID50) for 1 h. The infected cells were washed by PBS three times and then fresh MEM was added. 12, 24, 48 and 72 h later, the cultured supernatants was collected from uninfected and infected Marc-145 cells and assayed for IFN-α using commercially available ELISA kits according to the procedures provided by the manufacturer. The PRRSV RNA loading capacity was determined by Real time-PCR.
The virus-induced CPE of the cells was observed and scored under light microscopy in comparison with the negative control. When the untreated virus-infected cells reached ++++ CPE, the MTT assay was carried out. The inhibition rate (I %) was estimated as the formula below (Gescher et al., 2011).
where, ODT( PRRSV) represents the Optical Density (OD) of cells infected with PRRSV and treated with the compounds, ODC(PRRSV) corresponds to the OD of the untreated PRRSV-infected cells and ODC(mock) is the OD of untreated mock-infected cells. The 50% inhibitive concentration (IC50) of the compound was determined as 50% cytoprotection against PRRSV infection.
Direct virucidal effect on PRRSV: PRRSV was incubated with a series concentration of forsythoside A (10, 20, 40 and 60 μg mL-1) at 37°C for 1 h. Then the cell monolayers in 24-well was infected with the drug-pretreated virus (100 TCID50) at 37°C for 1 h. After washing with PBS for three times, then fresh MEM was added. To determine residual infectivity in a plaque formation assay the CPE observation and MTT assay were carried out post-infection as the same to the above. The virucidal concentration 50% (VC50) defined as the concentration required to reduce virus titer by 50%, was determined.
Treatment of infected Marc-145 cells: Marc-145 cells (3×105 cells) were grown in 24-well plastic tissue culture plates. Confluent monolayer of the cell cultures was inoculated with PRRSV (100 TCID50) for 1 h. Infected cells were washed with PBS three times to remove the unbound virus and then the infected cells were incubated with a series of forsythoside A dissolved in MEM for 72 h. Each concentration of forsythoside A was repeated four times. The CPE observation and MTT assay were carried post-infection as the same to the above.
Parameters calculation and statistical assay: Statistical significance was assessed by one-way analysis of variance (ANOVA) and Student’s t-test (GraphPad Software, San Diego, CA) to determine the statistical significance of differences between the test samples and control. A value of p<0.05 was considered statistically significant. Data were expressed as mean SD.
RESULTS
Titration of PRRSV: The cells CPE was observed induced by PRRS virus with eight replicate wells under each dilution. The cells infected with PRRSV showed early CPE on monolayer of cells in a well at about 72 h postinfection. The normal cells appeared round ridge with an intact structure and well-defined border. In regard to the cells CPE appeared at 0 and 100%. The TCID50 of PRRSV was 10-5.7/0.1 mL according to the Reed-Muench Methods.
Toxicity detection for forsythoside A: To rule out the possibility that nonspecific toxicity caused by forsythoside A, its cytotoxicity was observed and assayed by MTT test. Forsythoside A showed non-toxic on Marc-145 cells with concentrations up to 60 μg mL-1. Its cytotoxic concentration 50% (CC50) was 89 μg mL-1.
Forsythoside A showed an inhibitive efficiency on PRRSV infection: The antiviral infection activity of the forsythoside A against PRRS virus was investigated. The cytopathic effect of three kinds of infection modes were shown in Fig. 1.
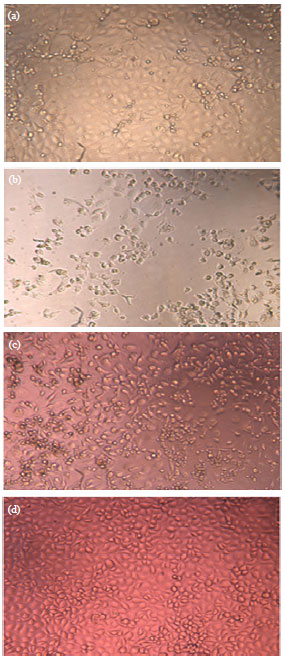 | |
| Fig. 1(a-d): | Protective efficiency of 40 μg mL-1 forsythiaside against PRRSV infection by cytopathic effect observation (a) Treatment prior infection, (b) Treatment of infected cells, (c) Direct antivirus effect and (d) Mocked control |
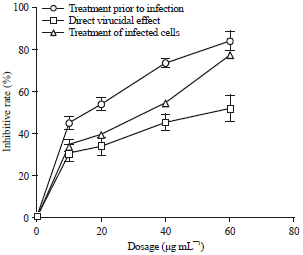 | |
| Fig. 2: | PRRSV inhibitive rate by forsythiaside A |
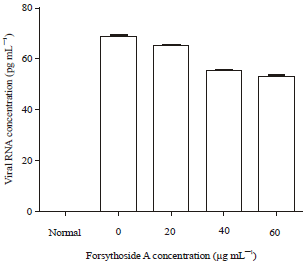 | |
| Fig. 3: | Inhibitive efficiency for PRRSV RNA synethsis by forsythoside A |
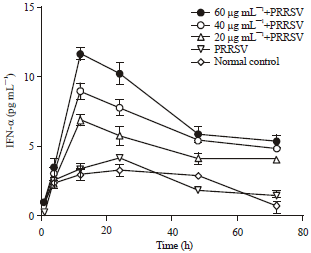 | |
| Fig. 4: | Inductive effect for IFN-⟨ secretion in PRRSV-infected Marc-145 cells. The peak concentration was at 12 h after treatment |
Three kinds of anti-infection efficiency were all tested positive. The 50% virus inhibitive rates (IC50) were 10.6, 18.4 and 41.9 μg mL-1 of the drug under three modes, respectively, for treatment prior to infection, treatment of the infected cells and direct virucidal effect on PRRSV using forsythside A. The viral inhibitive curves were shown in Fig. 2. The virus inhibitive rate raised with the increase of forsythoside A concentrations in each treatment mode. Thus Forsythoside A is an effective antivirus agent which showed inhibitive PRRSV from infection in vitro and the inhibiting infection showed to be dose-dependent and time-dependent.
Compared among the three kinds of treatments, the best efficiency was treatment prior to PRRSV infection, the second was the treatment of the infected cells, the worst was the direct virucidal effect. The inhibitive rate was near 80% when treated with 60 μg mL-1 of forsythoside A before and after infectionwhile virus inhibitive rate was only 55.6% under the direct virucidal mode.
Forsythoside A inhibited PRRSV RNA synthesis: The effect of forsythoside A mediated inhibition of PRRSV RNA synthesis was also investigated. Because the best efficiency was treatment prior to PRRSV infection, the inhibitive efficiency for RNA sythesis was experimented under this mode. The viral RNA copies in cells were quantified by real-time QPCR and the PRRSV ORF7 plasmid standard curves. Compared with the non-drug-treated cells, the addition of forsythoside A resulted in a significant reduction in viral RNA copies (p<0.05) (Fig. 3). The intracellular viral RNA level decreased with the increase of forsythoside A concentration. Thus, forsythoside could dose-dependently inhibit the replicate of viral RNA in Marc 145 cells efficiently.
Forsythoside A induced IFN-α secretion on PRRSV-infected cells: For the assay of IFN-α production under the treatment of forsythoside A, Marc-145 cells were grown infected by PRRSV in the absence or presence of forsythoside A for 3 days, respectively. Cultured supernatants were assayed for IFN-α at a series time point. Compared with cells infected with PRRSV alone, IFN-α production significantly increased in the presence of 20, 40 and 60 μg mL-1 of forsythoside A. while PRRSV infection poorly induced the production of IFN-α (Fig. 4). It demonstrated that forsythoside A had obviously promotive effect on IFN-α secretion in a concentration-dependent manner.
DISCUSSION
PRRSV infection has led to serious economic loss. Drug control is an alternative therapy. But because the synthesize antivirus drugs is time consuming, the development of antivirus drugs is very slow compared with that of antibacterial drugs. Intensive efforts have been made to look for anti-PRRSV agents from natural medicinal herbs in vitro (Cheng et al., 2013; Karuppannan et al., 2012; Wang et al., 2013a; Yang et al., 2013). The fruit of Forsythia suspensa Vahl (Oleaceae) is a traditional Chinese medicinal herb. Forsythoside A is one of the main active ingredients. There are many reports about its anti-inflammatory (Jiang et al., 2010a, b, 2012) and antiviral activity (Li et al., 2011). The purpose of the present study was to identify its effect for PRRSV infection in vitro.
It was found that forsythoside A had potent antiviral effects against PRRSV infection at 3 treatment concentration of 20, 40 and 60 μg mL-1. It exerted the best therapy when treatment before the PRRSV infection, the second was treatment the infected cells while the direct virucidal effect was worst. The forsythoside A showed anti-PRRS virus effect for that the CPE induced by PRRSV decreased and cells viable rate promoted, with increasing concentrations of forsythoside A. According to the results of toxicity detection of forsythoside A, it was indicated that forsythoside A had no toxicity to Marc-145 cells at the test concentration range. Furthermore, forsythoside A reduced intracellular PRRSV RNA synthesis in virus-infected cells and consequently inhibited production of progeny virus in infected cells.
The causal PRRSV could escape from innate immunity and causes persistent infections. Type I IFNs ( IFN-α/β) play an important role in innate immunity against viral infections by inducing antiviral responses. The IFN-a constitutes an early host response to PRRSV (Chung et al., 2004) but PRRSV circumvents the host innate response, inadequately induce or even suppress type I IFNs production (Charerntantanakul et al., 2013; Patel et al., 2010; Sipos et al., 2003; Wang et al., 2013b); consequently, production of IFN-γ, cellular immunity, neutralizing antibodies and viral clearance are delayed (Murtaugh et al., 2002; Xiao et al., 2004). Thus IFN-α is critical for the innate immunity and play an important role in the modulation of adaptive immunity. Regulating IFN-α secretion by drug may interrupt the viral infection. In this study, it was found that forsythoside A continously promoted the IFN-α excrection which provided a possibility that forsythoside A inhibit the PRRSV replication by regulating some antiviral cytokines.
CONCLUSION
This study indicated that forsythoside A may possesses a significant anti-PRRSV effect and can enhance PRRSV-suppressed IFN-α secretion by Marc-145 cells which may be beneficial for the treatment of native immunity conditions followed by viral infection. It could prove useful for further antivirus research on PRRSV infections. However, its definite antiviral mechanism is still needed to further research.
ACKNOWLEDGEMENT
This study was supported by grants from China National Natural Science Foundation (31372485, 31172362) and the National Twelve-Five Technological Supported Plan of China (2011BAD34B).
REFERENCES
- Charerntantanakul, W., S. Yamkanchoo and W. Kasinrerk, 2013. Plasmids expressing porcine interferon gamma up-regulate pro-inflammatory cytokine and co-stimulatory molecule expression which are suppressed by porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol., 153: 107-117.
CrossRef - Chen, N., Z. Cao, X. Yu, X. Deng and T. Zhao et al., 2011. Emergence of novel European genotype porcine reproductive and respiratory syndrome virus in mainland China. J. Gen. Virol., 92: 880-892.
CrossRefDirect Link - Cheng, J., N. Sun, X. Zhao, L. Niu and M. Song et al., 2013. In vitro screening for compounds derived from traditional Chinese medicines with antiviral activities against porcine reproductive and respiratory syndrome virus. J. Microbiol. Biotechnol., 23: 1076-1083.
PubMedDirect Link - Chung, H.K., J.H. Lee, S.H. Kim and C. Chae, 2004. Expression of interferon-α and Mx1 protein in pigs acutely infected with Porcine Reproductive and Respiratory Syndrome Virus (PRRSV). J. Comp. Pathol., 130: 299-305.
CrossRefDirect Link - Ferrari, L., P. Martelli, R. Saleri, E. de Angelis and V. Cavalli et al., 2013. Lymphocyte activation as cytokine gene expression and secretion is related to the Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) isolate after in vitro homologous and heterologous recall of Peripheral Blood Mononuclear Cells (PBMC) from pigs vaccinated and exposed to natural infection. Vet. Immunol. Immunopathol., 151: 193-206.
CrossRefDirect Link - Gao, L., W. Zhang, Y. Sun, Q. Yang and J. Ren et al., 2013. Cryptoporus volvatus extract inhibits Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) in vitro and in vivo. PLoS ONE, Vol. 8.
CrossRef - Gescher, K., J. Kuhn, W. Hafezi, A. Louis and A. Derksen et al., 2011. Inhibition of viral adsorption and penetration by an aqueous extract from Rhododendron ferrugineum L. as antiviral principle against herpes simplex virus type-1. Fitoterapia, 82: 408-413.
CrossRef - Jiang, W.L., F.H. Fu, B.M. Xu, J.W. Tian and H.B. Zhu, 2010. Cardioprotection with forsythoside B in rat myocardial ischemia-reperfusion injury: Relation to inflammation response. Phytomedicine, 17: 635-639.
CrossRefDirect Link - Jiang, W.L., J.W. Tian, F.H. Fu, H.B. Zhu and J. Hou, 2010. Neuroprotective efficacy and therapeutic window of Forsythoside B: In a rat model of cerebral ischemia and reperfusion injury. Eur. J. Pharmacol., 640: 75-81.
CrossRefDirect Link - Jiang, W.L., Yong-Xu, S.P. Zhang, H.B. Zhu and Jian-Hou, 2012. Forsythoside B protects against experimental sepsis by modulating inflammatory factors. Phytother. Res., 26: 981-987.
CrossRefDirect Link - Karuppannan, A.K., K.X. Wu, J. Qiang, J.J.H. Chu and J. Kwang, 2012. Natural compounds inhibiting the replication of Porcine reproductive and respiratory syndrome virus. Antiviral Res., 94: 188-194.
CrossRef - Li, H., J. Wu, Z. Zhang, Y. Ma, F. Liao, Y. Zhang and G. Wu, 2011. Forsythoside a inhibits the avian infectious bronchitis virus in cell culture. Phytother. Res., 25: 338-342.
CrossRefDirect Link - Murtaugh, M.P. and M. Genzow, 2011. Immunological solutions for treatment and prevention of Porcine Reproductive and Respiratory Syndrome (PRRS). Vaccine, 29: 8192-8204.
CrossRefDirect Link - Murtaugh, M.P., Z. Xiao and F. Zuckermann, 2002. Immunological responses of swine to porcine reproductive and respiratory syndrome virus infection. Viral Immunol., 15: 533-547.
CrossRefDirect Link - Neumann, E.J., J.B. Kliebenstein, C.D. Johnson, J.W. Mabry and E.J. Bush et al., 2005. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J. Am. Vet. Med. Assoc., 227: 385-392.
CrossRefPubMedDirect Link - Patel, D., Y. Nan, M. Shen, K. Ritthipichai, X. Zhu and Y.J. Zhang,, 2010. Porcine reproductive and respiratory syndrome virus inhibits type I interferon signaling by blocking STAT1/STAT2 nuclear translocation. J. Virol., 84: 11045-11055.
CrossRefPubMedDirect Link - Quinting, B., B. Robert, C. Letellier, M. Boxus, P. Kerkhofs, F. Schynts and A. Collard, 2007. Development of a 1-step enzyme-linked immunosorbent assay for the rapid diagnosis of bovine respiratory syncytial virus in postmortem specimens. J. Vet. Diagn. Invest., 19: 238-243.
CrossRefDirect Link - Sipos, W., C. Duvigneau, P. Pietschmann, K. Holler and R. Hartl et al., 2003. Parameters of humoral and cellular immunity following vaccination of pigs with a European modified-live strain of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV). Viral Immunol., 16: 335-346.
CrossRefDirect Link - Wang, R., Y. Xiao, T. Opriessnig, Y. Ding and Y. Yu et al., 2013. Enhancing neutralizing antibody production by an interferon-inducing porcine reproductive and respiratory syndrome virus strain. Vaccine, 31: 5537-5543.
CrossRefPubMedDirect Link - Wang, Z.W., N. Sun, C.H. Wu, J.B. Jiang, Y.S. Bai and H.Q. Li, 2013. In vitro antiviral activity and underlying molecular mechanisms of dipotassium glycyrrhetate against porcine reproductive and respiratory syndrome virus. Antiviral Ther., 18: 997-1004.
CrossRefDirect Link - Xiao, Z., L. Batista, S. Dee, P. Halbur and M.P. Murtaugh, 2004. The level of virus-specific T-Cell and macrophage recruitment in porcine reproductive and respiratory syndrome virus infection in pigs is independent of virus load. J. Virol., 78: 5923-5933.
CrossRefDirect Link - Yang, Q., L. Gao, J. Si, Y. Sun, J. Liu, L. Cao and W.H. Feng, 2013. Inhibition of porcine reproductive and respiratory syndrome virus replication by flavaspidic acid AB. Antiviral Res., 97: 66-73.
CrossRefDirect Link