
Tomoe Ishikawa
Graduate School of Pharmaceutical Sciences, University of Tokyo, Tokyo, 113-0033, Japan
Kazuki Okamoto
Graduate School of Pharmaceutical Sciences, University of Tokyo, Tokyo, 113-0033, Japan
Yuji Ikegaya
Graduate School of Pharmaceutical Sciences, University of Tokyo, Tokyo, 113-0033, Japan
International Journal of Pharmacology
Year: 2015 | Volume: 11 | Issue: 4 | Page No.: 318-326







ABSTRACT
Glutamate mediates the majority of excitatory neurotransmission in the brain. Thus, blockade of non-NMDA-type receptors, the main type of ionotropic glutamate receptors under baseline conditions is expected to eliminate neuronal network activity. In the present work, we challenge this simple notion by showing the stability of spontaneous neuronal activity in cultured hippocampal networks in situ. We monitored spiking activity of hippocampal CA3 neuron populations by using a functional Multineuron Calcium Imaging (fMCI) technique. Bath application of competitive non-NMDA receptor antagonists decreased excitatory neurotransmission by approximately 80%. Surprisingly, however, it did not change the level of spontaneous network activity. The antagonists also reduced inhibitory synaptic inputs in CA3 pyramidal cells and thereby maintained the ratio between excitation and inhibition as a whole. Moreover, the antagonists induced an increase in the input resistance of CA3 pyramidal cells. These compensatory adaptations in excitability balance and neuronal intrinsic properties may provide ongoing network activity with homeostatic robustness against an external perturbation of non-NMDA receptors. Interestingly, the non-NMDA receptor antagonists reduced epilepsy-like synchronous hyperactivity to the normal activity level.
PDF Abstract XML References Citation
Received: January 30, 2015;
Accepted: March 21, 2015;
Published: April 18, 2015
How to cite this article
Tomoe Ishikawa, Kazuki Okamoto and Yuji Ikegaya, 2015. Homeostatic Hippocampal Activity against Reduced Glutamatergic Neurotransmission. International Journal of Pharmacology, 11: 318-326.
DOI: 10.3923/ijp.2015.318.326
URL: https://scialert.net/abstract/?doi=ijp.2015.318.326
DOI: 10.3923/ijp.2015.318.326
URL: https://scialert.net/abstract/?doi=ijp.2015.318.326
INTRODUCTION
Neuronal networks are spontaneously active even in the absence of sensory inputs. Spontaneous activity accounts for an overwhelming fraction of energy consumption in the brain (Arieli et al., 1996) and is fundamental to neural information processing (Arieli et al., 1996; Pare et al., 1998; Petersen et al., 2003; Shu et al., 2003a; Ikegaya et al., 2013). Its activity patterns are associated with the network states (Destexhe and Contreras, 2006) and sometimes report the pathological states in neurological and psychiatric diseases (Greicius et al., 2004; Rombouts et al., 2005; Garrity et al., 2007). Thus, elucidating spontaneous activity is essential for our understanding of the brain function and malfunction, however, the network mechanisms by which spontaneous activity is stably maintained have not been fully clarified.
In the telencephalon, including the cerebral cortex and the hippocampus, excitatory and inhibitory synaptic neurotransmissions are mediated mainly by glutamate and GABA, respectively (Kandel et al., 2012). Ionotropic receptors of glutamate are roughly classified into two types, N-methyl-d-aspartate (NMDA) receptors and non-NMDA receptors; non-NMDA receptors contribute mainly to baseline excitatory transmission at the resting membrane potential level. The proper balance between glutamatergic and GABAergic transmissions is believed to be essential for preserving normal network activity. Indeed, proportional fluctuations in excitatory and inhibitory synaptic conductances emerge in spontaneous and sensory-evoked cortical activity (Anderson et al., 2000; Monier et al., 2003; Shu et al., 2003b; Swadlow, 2003; Wehr and Zador, 2003; Tan et al., 2004; Wilent and Contreras, 2005; Okun and Lampl, 2008; Wu et al., 2008; Poo and Isaacson, 2009), suggesting that a tightly regulated interaction between glutamatergic and GABAergic synaptic transmission underlies stable network activity. On the other hand, excessive excitation relative to inhibition results in pathological hyperactivity with synchronous burst discharges (Ikegaya, 1999; Matsumoto et al., 2013). Therefore, a disruption of the excitation-to-inhibition (E/I) relationship is expected to perturb spontaneous network activity.
In contrast to this idea, we found that a reduction of excitatory synaptic activity by non-NMDA receptor antagonists does not affect the level of spontaneous network activity. We thus investigated the synaptic mechanisms underlying these homeostatic properties of spontaneous activity.
MATERIAL AND METHODS
Animals: Experiments were performed with the approval of the animal experiment ethics committee at the University of Tokyo (approval No. P24-6) and according to the University of Tokyo guidelines for the care and use of laboratory animals. Wistar/ST rats (either male or female) were housed in cages under standard laboratory conditions (12 h light/dark cycle, ad libitum access to food and water). All efforts were made to minimize the animals’ suffering and the number of animals used.
Slice preparations: Entorhinal-hippocampal organotypic slices were prepared from postnatal day 7 Wistar/ST rats of either sex (Koyama et al., 2007). Rat pups were deeply anesthetized by hypothermia and decapitated. The brains were removed and placed in aerated ice-cold Gey’s balanced salt solution supplemented with 25 mM glucose. Horizontal entorhinal-hippocampal slices were cut at a thickness of 300 μm using a vibratome. Slices were placed on Omnipore membrane filters and incubated in 5% CO2 at 37°C. The culture medium was composed of 50% minimal essential medium, 25% Hanks’ balanced salt solution supplemented with 133 mM glucose, 25% horse serum and antibiotics and was changed every 3.5 days. Experiments were performed at 6-12 days in vitro.
Functional multineuron calcium imaging: Oregon Green 488 BAPTA-1AM was dissolved in DMSO containing 10% Pluronic F-127 to yield a concentration of 200 μM (Takahashi et al., 2007, 2011). Immediately before use, this solution was diluted tenfold with oxygenated Artificial Cerebrospinal Fluid (aCSF) consisting of (mM): 127 NaCl, 26 NaHCO3, 1.5 KCl, 1.24 KH2PO4, 1.4 MgSO4, 2.4 CaCl2, 10 glucose and bubbled with 95% O2 and 5% CO2. The preparations were washed three times with oxygenated aCSF. They were transferred into a dish (35-mm diameter) containing 2 mL of the dye solution and were incubated for 1 h in a humidified incubator at 35°C under 5% CO2. After being washed, the cultured slices were incubated at 35°C for 40 min and were mounted in a recording chamber perfused at 6-8 mL min-1 with oxygenated aCSF at 35-37°C. Fluorophores were excited at 488 nm with a laser diode and visualized using 507 nm long pass emission filters. Videos were recorded at 10 frames/s using a 16x objective (0.8 numerical aperture, Nikon), a spinning-disk confocal microscope (CSU-X1, Yokogawa Electric, Tokyo, Japan), a cooled CCD camera (iXonEM+DV897, Andor Technology, Belfast, UK) and an upright microscope (Eclipse FN1, Nikon, Tokyo, Japan). The fluorescence change was measured as:
(Ft-F0)/F0
where, Ft is the fluorescence intensity at time t and F0 is the fluorescence intensity averaged from -10 to 10 sec relative to t. Using the principal component analysis and a support vector machine optimized to calcium imaging, spike-elicited calcium transients were semiautomatically detected with a custom-written program in Visual Basic (Sasaki et al., 2008).
Electrical physiological recordings: Recordings were performed in a submerged chamber perfused at 6-8 mL min-1 with oxygenated aCSF at 35°C unless otherwise specified. Whole-cell patch-clamp recordings were obtained from hippocampal pyramidal cells visually identified under infrared differential interference contrast microscopy. Patch pipettes (3-6 MΩ) were filled with a potassium-based solution consisting of (in mM) 120 potassium gluconate, 10 KCl, 10 HEPES, 10 creatine phosphate, 4 Mg-ATP, 0.3 Na2-GTP and 0.2 EGTA for current-clamp recordings or a cesium-based solution consisting of (in mM) 130 CsMeSO4, 10 CsCl, 10 HEPES, 10 creatine phosphate, 4 Mg-ATP and 0.3 Na2-GTP for voltage-clamp recordings. For cell-attached recordings, pipettes were filled with aCSF. Excitatory and inhibitory postsynaptic currents (EPSCs and IPSCs) were recorded at clamped voltages of -70 and 0 mV, respectively. For electrical field stimulation, glass pipettes were filled with aCSF and were placed in the CA3 stratum radiatum and 60 single rectangular-pulse stimuli (20 μsec duration at 30-80 μA) were applied at intervals of 10 sec. The membrane properties were measured in the whole-cell current-clamp configuration. Series resistance was not compensated. The signals were amplified and digitized at a sampling rate of 20 kHz using a MultiClamp 700B amplifier and a Digidata 1440A digitizer that were controlled by pCLAMP 10.4 software (Molecular Devices). Data were analyzed offline using custom MATLAB R2013b (The MathWorks) routines. Synaptic events were detected from EPSC and IPSC traces. Events with amplitudes of 3-7 SDs of the baseline noise after local filtering and smoothing were defined as synaptic events.
Pharmacology: Tetrodotoxin (2 μM), picrotoxin (50 μM), l,d-2-amino-5-phosphonopentanoic acid (AP5, 50 μM), 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX, 50 μM) and 6,7- dinitroquinoxaline-2,3-dione (DNQX, 50 μM) were used in the experiment. The drug solution was prepared immediately before use and perfused into the recording chamber.
Statistics: We reported the data as Mean±SDs unless otherwise specified. Paired t-test, Welch’s t-test, Wilcoxon signed-rank test and one-way repeated-measure ANOVA were performed to assess the significance of the difference. p<0.05 was considered statistically significant.
RESULTS
CNQX reduces the amplitude of evoked EPSCs: To examine spontaneous network activity in vitro, we employed organotypic network cultures of hippocampal slices, because these networks are known to emit spontaneous activity that resembles to in vivo networks in the brain (Takahashi et al., 2010; Okamoto et al., 2014). First, we confirmed the efficacy of CNQX, a competitive non-NMDA receptor antagonist, on excitatory synaptic transmission. While compound EPSCs evoked by field stimulation of CA3 afferents were recorded from CA3 pyramidal cells at a clamped voltage at -70 mV, CNQX was bath-applied to the recording chamber at 50 μM. At the steady state of the drug effect (10 min after CNQX perfusion), the mean EPSC amplitude was decreased to 16.3±15.2% of the control level (Fig. 1, control: 580.5±192.9 pA, CNQX: 100.9±94.6 pA; p = 0.012, t3 = 5.4, paired t-test, Mean±SD of 4 cells).
CNQX does not abolish spontaneous activity: To examine the effect of CNQX on spontaneous activity of hippocampal networks, we recorded spiking activity in individual CA3 neurons using fMCI, in which 96.9±7.2 neurons were simultaneously monitored per video (Mean±SD of 11 videos from 11 slices). Cell-attached recording revealed that transient calcium activity of individual neurons reflected action potentials of the neurons (Fig. 2a). Under baseline conditions prior to bath-application of CNQX, all slices tested spontaneously emitted calcium activity (Fig. 2b). The mean activity rate of individual neurons was 1.62±0.55 events/cell/min (Mean±SD of = 11 slices).
We then perfused these spontaneously active slices with 50 μM CNQX. The spontaneous activity did not disappear even 10 min after the onset of CNQX perfusion (Fig. 2c, n = 4 slices). The same phenomenon was replicable using DNQX, another competitive non-NMDA receptor antagonist (n = 3 slices). Thus, data of CNQX and DNQX were pooled in Fig. 2d. After application of CNQX and DNQX, the mean activity rate was 1.86±1.19 events/cell/min (Mean±SD of 7 slices) and was not significantly different from the pre-application control level (p = 0.64, t8 = 0.49, Welch’s t-test). Because calcium transients were still a faithful reporter of action potentials in the presence of CNQX (Fig. 2a right) and also because 2 mM tetrodotoxin readily eliminated calcium activity (Fig. 2d, 0.11±0.08 events/cell/min, Mean±SD of 4 slices, p = 4.1×10&-5, t4 = 19.5, Welch’s t-test), we concluded that neither CQNX nor DNQX changed the frequencies of spontaneous activity.
After addition of 50 μM AP5, a competitive NMDA receptor antagonist, to 50 μM CNQX and DNQX, the mean activity rate was 1.48±0.80 events/cell/min (Fig. 2d, Mean±SD of 4 slices) which was not significantly different from the control level (p = 0.67, t6 = 0.47). When 50 μM picrotoxin, a GABAA receptor antagonist was further added to the CNQX/DNQX and AP5 solution, the mean activity rate was reduced to 0.26±0.17 events/cell/min (Fig. 2d, Mean±SD of 4 slices, p = 1.1×10‾4, t6 = 8.9 versus control). These data indicated that the impairment of both ionotropic glutamatergic and GABAergic synaptic transmission attenuates spontaneous activity, although the reduction in glutamatergic transmission alone does not.
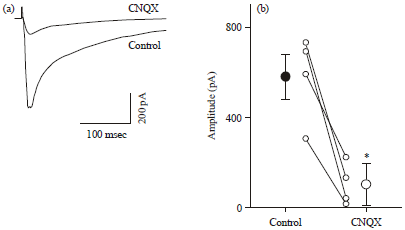 | |
| Fig. 1(a-b): | CNQX reduces hippocampal synaptic transmission (a) Representative whole-cell traces of evoked EPSCs in a CA3 pyramidal cell 5 min before (control) and 10 min after bath-perfusion of 50 μM CNQX. EPSCs were evoked by stimulation of the CA3 stratum radiatum and (b) Mean EPSC amplitudes before and after bath-perfusion of 50 μM CNQX. Error bars are SDs of 4 cells. *p = 0.012, t3 = 5.4, paired t-test |
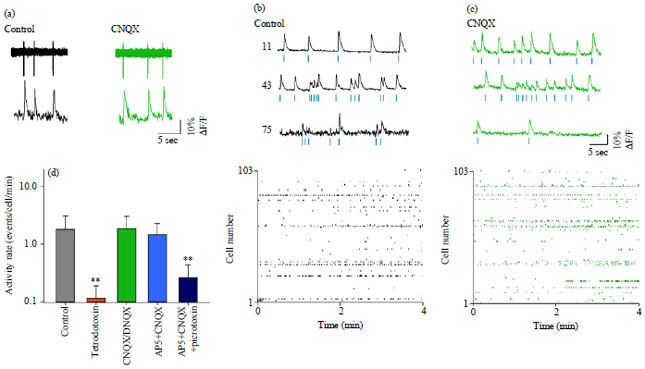 | |
| Fig. 2(a-d): | Non-NMDA receptor antagonists do not affect spontaneous network activity level, (a) Simultaneous cell-attached recording and calcium imaging before and 10 min after perfusion with 50 μM CNQX. Individual calcium transients reflected action potentials of the neuron, (b-c) Calcium traces recorded from 3 representative CA3 neurons (top) and raster plots of spontaneous activity of a total 103 neurons (bottom) before (b) and 10 min after CNQX application (c). Blue dots below the traces indicate the timings of detected spikes and (d) Mean±SD firing rates of CA3 neurons before (control, n = 11 slices) and after bath application of tetrodotoxin (n = 4 slices), CNQX/DNQX (n = 7 slices), CNQX/DNQX+AP5: (n = 4 slices), CNQX/DNQX+AP5+picrotoxin (n = 4 slices). **p<0.01, Welch’s t-test after one-way repeated-measure ANOVA (p = 1.3×10‾3, F4,25 = 6.1) |
CNQX attenuates both excitatory and inhibitory synaptic transmission: To elucidate the mechanisms of the network stability against reduced non-NMDA receptor activity, we recorded spontaneous EPSCs (sEPSCs) and IPSCs (sIPSCs) from CA3 pyramidal cells in the voltage-clamp configuration. Like the case of evoked EPSCs (Fig. 1), CNQX decreased the mean amplitudes of sEPSCs (Fig. 3a, p = 0.016, W = 21, Wilcoxon signed-rank test, n = 6 cells). However, it did not change the mean event frequencies of sEPSCs (Fig. 3a, p = 0.46, W = 10). It should be noted that no change in the sEPSC frequencies is consistent with our finding that CNQX did not alter the mean spike frequency of CA3 neurons, the main presynaptic cell populations (Fig. 2d).
Interestingly, CNQX also reduced the mean amplitudes, but not mean frequencies of sIPSCs (Fig. 3b, amplitude: p = 0.016, W = 21, frequency: p = 0.093, W = 19, Wilcoxon signed-rank test, n = 6 cells). Because the extent of CNQX-induced reduction in sIPSCs was similar to that of sEPSCs, we considered that the E/I balance was preserved after CNQX application. To confirm this possibility, we calculated the charge of synaptic inputs, i.e., the areas under the curves of individual sEPSC and sIPSC traces and computed the E/I ratio by dividing the mean charge of sEPSCs by that of sIPSCs. Indeed, the E/I ratios did not change before and after CNQX application (Fig. 3c, p = 0.12, t7 = 1.7, Welch’s t-test, n = 6 cells). This balanced reduction in the excitation and inhibition levels is likely a reason why spontaneous activity was maintained in the presence of CNQX.
CNQX increases the membrane resistance of neurons: The interplay between excitation and inhibition is known to regulate neuronal output by modulating the membrane potential and the input resistance of the neuron (Higley and Contreras, 2006). We next examined the effect of CNQX on the fundamental membrane properties of CA3 pyramidal cells using whole-cell recordings. Neither the resting membrane potentials (Fig. 4a, p = 0.33, t7 = 1.0, paired t-test, n = 8 cells) nor membrane fluctuations (Fig. 4b, p = 0.71, t8 = 0.39, n = 9 cells) differed before and after bath-application of 50 mM CNQX. However, CNQX increased the input resistances (Fig. 4c, p = 0.041, t6 = 2.6, n = 7 cells) without affecting the membrane capacitance (Fig. 4d, p = 0.31, t6 = 1.1, n = 7 cells).
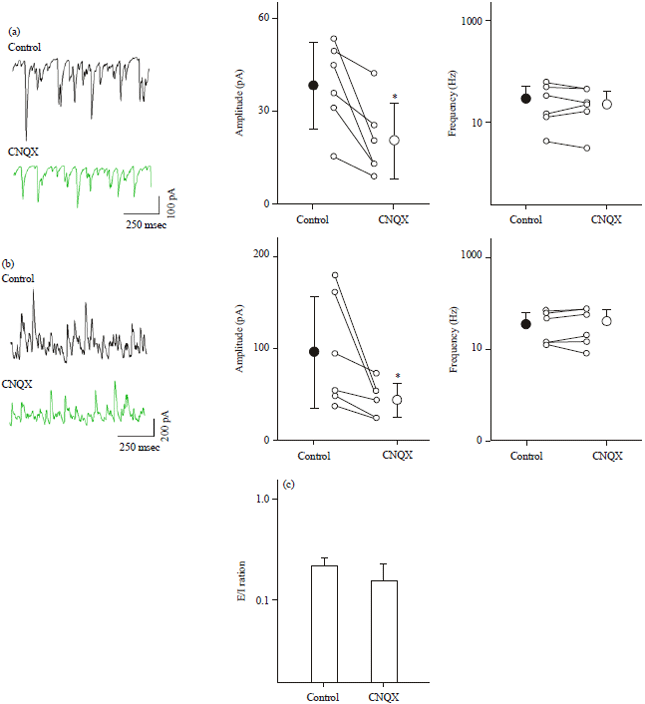 | |
| Fig. 3(a-c): | CNQX reduces both sEPSC and sIPSC amplitudes to a similar level, (a) Representative sEPSC traces recorded from a CA3 pyramidal cell 5 min before and 10 min after CNQX treatment (left). The neuron was clamped at -70 mV. The amplitude (middle) and frequency (right) of sEPSCs were summarized as the Mean±SD of 6 neurons, *p = 0.016, W = 21, Wilcoxon signed-rank test, (b) Same as a, but for IPSCs recorded at 0 mV. *p = 0.016. W = 21 and (c) Mean±SD E/I ratios of 6 neurons. p = 0.12, t7 = 1.7, Welch’s t-test |
The input resistance is a parameter that is linked to neuronal responsiveness, that is, its increase makes the neuron’s membrane potential more sensitive to synaptic inputs. Thus, the increased input resistance may counteract reduced sEPSC amplitudes.
CNQX reduces epileptic burst activity: The unbalance of excitation and inhibition may induce highly synchronized network activity (Avanzini and Franceschetti, 2003b; Dudek and Sutula, 2007) which is often observed in epileptic conditions. Recently developed antiepileptic drugs include non-NMDA receptor antagonists (Loscher and Honack, 1994; Bleakman and Lodge, 1998; De Sarro et al., 2005; Hanada et al., 2011; Rogawski, 2011). We hypothesized that while CNQX does not affect baseline neuronal activity, it could reduce pathological hyperactivity to the normal range of network activity. To evaluate the effect of CNQX on the epileptic burst activity, we returned to fMCI experiments and monitored spiking activity of CA3 neurons. Synchronized discharges were induced by increasing the temperature of the recording chamber from 35-40.5°C. This heat-induced hyperactivity is used as an in vitro model of febrile seizures (Mizunuma et al., 2009; Koyama et al., 2012). When these "Epileptic" slices were perfused with CNQX, synchronized burst activity disappeared completely (Fig. 5a).
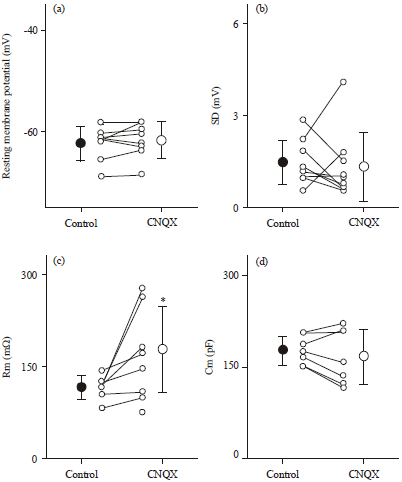 | |
| Fig. 4(a-d): | CNQX increases the membrane resistance of CA3 pyramidal cells, (a-d) Mean±SD of resting membrane potentials (a, n = 8 neurons), membrane fluctuation (b, n = 9), membrane resistance (c, n = 7) and membrane capacitance (d, n = 7). *p = 0.041, t6 = 2.6, paired t-test |
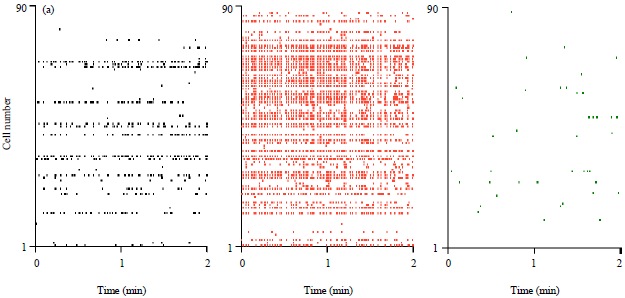 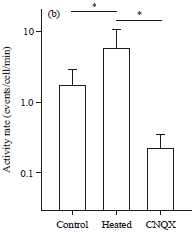 | |
| Fig. 5(a-b): | CNQX decreases epileptiform network activity, (a) Representative raster plots of the activity of 90 CA3 cells before heating (left, control), during 40.5°C heating (middle) and 10 min after CNQX application at 40.5°C (right), (b) Mean firing rates of 482 neurons from 5 slices are compared among these three conditions. Error bars are SDs of 5 slices. *p<0.05, paired t-test after one-way repeated-measure ANOVA (p = 0.020, F2, 12 = 5.6) |
The firing rates that had increased in the heated recording chamber were reduced by CNQX (Fig. 5b, control: 1.79±1.15 events/cell/min, heated: 6.56±4.48 events/cell/min, CNQX: 0.67±0.21 events/cell/min, SD of 5 slices, control versus heated, p = 0.048, t4 = 2.8, heated versus CNQX; p = 0.034, t4 = 3.2, paired t-test). The activity level after CNQX was not different from the pre-heating, control level (control versus heated; p = 0.066, t4 = 2.5).
DISCUSSION
In this study, we discovered that spontaneous activity was preserved under artificial conditions of largely reduced excitatory synaptic activity. This robust homeostasis seemed to be supported by a proportional reduction in inhibitory synaptic activity and an increase in the input resistance of individual neurons. A previous work has demonstrated that, during chronic application of DNQX for days, spontaneous network activity is first reduced and thereafter reappears in organotypic slice cultures of the visual cortex (Uesaka et al., 2005). However, our data indicated a more rapid homeostatic action of spontaneous activity, under which some different mechanisms must work to acutely compensate the reduced excitation.
CNQX is a non-NMDA receptor antagonist and thus, CNQX-induced decrease in the sEPSC amplitude is an expected result. However, it also reduced the sIPSCs amplitude in CA3 pyramidal cells. Both glutamatergic principle neurons and GABAergic interneurons receive excitatory synaptic inputs and thereby fire their action potentials. The CNQX would also reduce non-NMDA receptor-mediated excitatory inputs into GABAergic interneurons. GABAergic interneurons, especially basket cells that innervate the somata of pyramidal cells, often emit bursts of action potentials (Klausberger et al., 2003). Thus, CNQX may reduce the number of action potentials involved in individual bursts and thereby attenuate the amplitudes of sIPSCs in postsynaptic pyramidal cells.
The CNQX-induced increases in the input resistances of pyramidal cells seem to be a natural consequence of CNQX-induced decreases in sEPSCs and sIPSCs, because a reduction in the total number of open ion channels at postsynaptic sites causes a reduction in the total leak currents of the neurons. However, the increased input resistance can be explained by other mechanisms, such as rapid changes in receptor expression and spine morphology. Synaptic non-NMDA receptors are known to be laterally dispersed or internalize within minutes (Lissin et al., 1998, 1999; Carroll et al., 1999; Liao et al., 2001). The surface expression level of non-NMDA receptors is reported to determine the input resistance (Turrigiano et al., 1998). Thus, it is feasible that CNQX induced a change in the number of active non-NMDA receptors on synaptic surfaces. Morphological modifications of dendritic spine are known to control a gain between neuronal input and output (Araya et al., 2014). Blockade of non-NMDA receptor activity is reported to facilitate the spontaneous motility of spines (Fischer et al., 2000). This mechanism may also contribute to CNQX-induced increase in the input resistance.
Excessive glutamatergic neurotransmission is believed to cause epileptiform activity of neuronal networks and eventually induce systemic seizures (Avanzini and Franceschetti, 2003a; Meldrum and Rogawski, 2007; Scharfman, 2007). Based on this idea, non-NMDA receptor antagonists have recently been designed as a new class of antiepileptic drugs (Weiser, 2005), note that classical types of anticonvulsants are mainly aimed to enhance GABAergic function. Importantly, non-NMDA receptor antagonists seem to have no marked side effects (Loscher and Honack, 1994; Bleakman and Lodge, 1998; De Sarro et al., 2005; Hanada et al., 2011; Rogawski, 2011) and their action mechanisms remain to be elucidated. We found that CNQX abolishes epileptic burst activity without affecting normal spontaneous activity. We do not still know how CNQX acts selectively on anomalous spontaneous activity but this phenomenon will provide the first step to understand the mechanisms how non-NMDA receptor antagonists efficiently reduce seizure episodes without an apparent impairment in normal cognitive performance.
ACKNOWLEDGMENT
This study was supported by supported by Grants-in-Aid for Science Research on Innovative Areas (22115003, 25119004).
REFERENCES
- Anderson, J.S., M. Carandini and D. Ferster, 2000. Orientation tuning of input conductance, excitation and inhibition in cat primary visual cortex. J. Neurophysiol., 84: 909-926.
Direct Link - Araya, R., T.P. Vogels and R. Yuste, 2014. Activity-dependent dendritic spine neck changes are correlated with synaptic strength. Proc. Nat. Acad. Sci., 111: E2895-E2904.
CrossRefPubMedDirect Link - Arieli, A., A. Sterkin, A. Grinvald and A. Aertsen, 1996. Dynamics of ongoing activity: Explanation of the large variability in evoked cortical responses. Science, 273: 1868-1871.
Direct Link - Avanzini, P. and S. Franceschetti, 2003. Cellular biology of epileptogenesis. Lancet Neurol., 2: 33-42.
CrossRefDirect Link - Avanzini, G. and S. Franceschetti, 2003. Prospects for novel antiepileptic drugs. Curr. Opin. Invest. Drugs, 4: 805-814.
Direct Link - Bleakman, D. and D. Lodge, 1998. Neuropharmacology of AMPA and kainate receptors. Neuropharmacology, 37: 1187-1204.
PubMed - Carroll, R.C., D.V. Lissin, M. von Zastrow, R.A. Nicoll and R.C. Malenka, 1999. Rapid redistribution of glutamate receptors contributes to long-term depression in hippocampal cultures. Nat. Neurosci., 2: 454-460.
CrossRefDirect Link - De Sarro, G., R. Gitto, E. Russo, G.F. Ibbadu, M.L. Barreca, L. De Luca and A. Chimirri, 2005. AMPA receptor antagonists as potential anticonvulsant drugs. Curr. Top. Med. Chem., 5: 31-42.
PubMedDirect Link - Destexhe, A. and D. Contreras, 2006. Neuronal computations with stochastic network states. Science, 314: 85-90.
PubMed - Dudek, F.E. and T.P. Sutula, 2007. Epileptogenesis in the dentate gyrus: A critical perspective. Prog. Brain Res., 163: 755-773.
CrossRefDirect Link - Fischer, M., S. Kaech, U. Wagner, H. Brinkhaus and A. Matus, 2000. Glutamate receptors regulate actin-based plasticity in dendritic spines. Nat. Neurosci., 3: 887-894.
CrossRefDirect Link - Garrity, A.G., G.D. Pearlson, K. McKiernan, D. Lloyd, K.A. Kiehl and V.D. Calhoun, 2007. Aberrant default mode functional connectivity in schizophrenia. Am. J. Psychiatry, 164: 450-457.
Direct Link - Greicius, M.D., G. Srivastava, A.L. Reiss and V. Menon, 2004. Default-mode network activity distinguishes Alzheimer's disease from healthy aging: Evidence from functional MRI. Proc. Nat. Acad. Sci. USA., 101: 4637-4642.
PubMed - Hanada, T., Y. Hashizume, N. Tokuhara, O. Takenaka and N. Kohmura et al., 2011. Perampanel: A novel, orally active, noncompetitive AMPA-receptor antagonist that reduces seizure activity in rodent models of epilepsy. Epilepsia, 52: 1331-1340.
CrossRefDirect Link - Higley, M.J. and D. Contreras, 2006. Balanced excitation and inhibition determine spike timing during frequency adaptation. J. Neurosci., 26: 448-457.
CrossRefDirect Link - Ikegaya, Y., 1999. Abnormal targeting of developing hippocampal mossy fibers after epileptiform activities via L-type Ca2+ channel activation in vitro. J. Neurosci., 19: 802-812.
Direct Link - Ikegaya, Y., T. Sasaki, D. Ishikawa, N. Honma and K. Tao et al., 2013. Interpyramid spike transmission stabilizes the sparseness of recurrent network activity. Cerebral Cortex, 23: 293-304.
Direct Link - Klausberger, T., P.J. Magill, L.F. Marton, J.D.B. Roberts, P.M. Cobden, G. Buzsaki and P. Somogyi, 2003. Brain-state- and cell-type-specific firing of hippocampal interneurons in vivo. Nature, 421: 844-848.
CrossRefDirect Link - Koyama, R., R. Muramatsu, T. Sasaki, R. Kimura and C. Ueyama et al., 2007. A low-cost method for brain slice cultures. J. Pharmacol. Sci., 104: 191-194.
CrossRefDirect Link - Koyama, R., K. Tao, T. Sasaki, J. Ichikawa and D. Miyamoto et al., 2012. GABAergic excitation after febrile seizures induces ectopic granule cells and adult epilepsy. Nat. Med., 18: 1271-1278.
CrossRefDirect Link - Liao, D., R.H. Scannevin and R. Huganir, 2001. Activation of silent synapses by rapid activity-dependent synaptic recruitment of AMPA receptors. J. Neurosci., 21: 6008-6017.
Direct Link - Lissin, D.V., R.C. Carroll, R.A. Nicoll, R.C. Malenka and M. von Zastrow, 1999. Rapid, activation-induced redistribution of ionotropic glutamate receptors in cultured hippocampal neurons. J. Neurosci., 19: 1263-1272.
Direct Link - Lissin, D.V., S.N. Gomperts, R.C. Carroll, C.W. Christine and D. Kalman et al., 1998. Activity differentially regulates the surface expression of synaptic AMPA and NMDA glutamate receptors. Proc. Nat. Acad. Sci., 95: 7097-7102.
Direct Link - Loscher, W. and D. Honack, 1994. Effects of the non-NMDA antagonists NBQX and the 2,3-benzodiazepine GYKI 52466 on different seizure types in mice: Comparison with diazepam and interactions with flumazenil. Br. J. Pharmacol., 113: 1349-1357.
CrossRefDirect Link - Meldrum, B.S. and M.A. Rogawski, 2007. Molecular targets for antiepileptic drug development. Neurotherapeutics, 4: 18-61.
CrossRefDirect Link - Mizunuma, M., N. Takahashi, A. Usami, N. Matsuki and Y. Ikegaya, 2009. High-temperature, but not high-pressure, conditions alter neuronal activity. J. Pharmacol. Sci., 110: 117-121.
CrossRefDirect Link - Monier, C., F. Chavane, P. Baudot, L.J. Graham and Y. Fregnac, 2003. Orientation and direction selectivity of synaptic inputs in visual cortical neurons: A diversity of combinations produces spike tuning. Neuron, 37: 663-680.
CrossRefDirect Link - Okamoto, K., T. Ishikawa, R. Abe, D. Ishikawa and C. Kobayashi et al., 2014. Ex vivo cultured neuronal networks emit in vivo-like spontaneous activity. J. Physiol. Sci., 64: 421-431.
CrossRefDirect Link - Okun, M. and I. Lampl, 2008. Instantaneous correlation of excitation and inhibition during ongoing and sensory-evoked activities. Nat. Neurosci., 11: 535-537.
CrossRefDirect Link - Pare, D., E. Shink, H. Gaudreau, A. Destexhe and E.J. Lang, 1998. Impact of spontaneous synaptic activity on the resting properties of cat neocortical pyramidal neurons in vivo. J. Neurophysiol., 79: 1450-1460.
Direct Link - Petersen, C.C.H., T.T.G. Hahn, M. Mehta, A. Grinvald and B. Sakmann, 2003. Interaction of sensory responses with spontaneous depolarization in layer 2/3 barrel cortex. Proc. Nat. Acad. Sci., 100: 13638-13643.
Direct Link - Poo, C. and J.S. Isaacson, 2009. Odor representations in olfactory cortex: Sparse coding, Global inhibition and oscillations. Neuron, 62: 850-861.
CrossRefDirect Link - Rogawski, M.A., 2011. Revisiting AMPA receptors as an antiepileptic drug target. Epilepsy Curr., 11: 56-63.
CrossRefDirect Link - Rombouts, S.A.R.B., F. Barkhof, R. Goekoop, C.J. Stam and P. Scheltens, 2005. Altered resting state networks in mild cognitive impairment and mild Alzheimer's disease: An fMRI study. Hum. Brain Mapping, 26: 231-239.
CrossRefDirect Link - Sasaki, T., N. Takahashi, N. Matsuki and Y. Ikegaya, 2008. Fast and accurate detection of action potentials from somatic calcium fluctuations. J. Neurophysiol., 100: 1668-1676.
CrossRefDirect Link - Scharfman, H.E., 2007. The neurobiology of epilepsy. Curr. Neurol. Neurosci. Rep., 7: 348-354.
CrossRefDirect Link - Shu, Y., A. Hasenstaub, M. Badoual, T. Bal and D.A. McCormick, 2003. Barrages of synaptic activity control the gain and sensitivity of cortical neurons. J. Neurosci., 23: 10388-10401.
Direct Link - Shu, Y., A. Hasenstaub and D.A. McCormick, 2003. Turning on and off recurrent balanced cortical activity. Nature, 423: 288-293.
CrossRefDirect Link - Swadlow, H.A., 2003. Fast-spike interneurons and feedforward inhibition in awake sensory neocortex. Cerebral Cortex, 13: 25-32.
Direct Link - Takahashi, N., S. Oba, N. Yukinawa, S. Ujita and M. Mizunuma et al., 2011. High-speed multineuron calcium imaging using nipkow-type confocal microscopy. Curr. Protoc Neurosci., Vol. 2.
CrossRef - Takahashi, N., T. Sasaki, W. Matsumoto, N. Matsuki and Y. Ikegaya, 2010. Circuit topology for synchronizing neurons in spontaneously active networks. Proc. Nat. Acad. Sci., 107: 10244-10249.
Direct Link - Takahashi, N., T. Sasaki, A. Usami, N. Matsuki and Y. Ikegaya, 2007. Watching neuronal circuit dynamics through functional multineuron calcium imaging (fMCI). Neurosci. Res., 58: 219-225.
CrossRefDirect Link - Tan, A.Y.Y., L.I. Zhang, M.M. Merzenich and C.E. Schreiner, 2004. Tone-evoked excitatory and inhibitory synaptic conductances of primary auditory cortex neurons. J. Neurophysiol., 92: 630-643.
CrossRefDirect Link - Turrigiano, G.G., K.R. Leslie, N.S. Desai, L.C. Rutherford and S.B. Nelson, 1998. Activity-dependent scaling of quantal amplitude in neocortical neurons. Nature, 391: 892-896.
CrossRefDirect Link - Uesaka, N., S. Hirai, T. Maruyama, E.S. Ruthazer and N. Yamamoto, 2005. Activity dependence of cortical axon branch formation: A morphological and electrophysiological study using organotypic slice cultures. J. Neurosci., 25: 1-9.
CrossRefDirect Link - Wehr, M. and A.M. Zador, 2003. Balanced inhibition underlies tuning and sharpens spike timing in auditory cortex. Nature, 426: 442-446.
CrossRefDirect Link - Weiser, T., 2005. AMPA receptor antagonists for the treatment of stroke. Curr. Drug Targets-CNS Neurol. Disorders, 4: 153-159.
CrossRefDirect Link - Wu, G.K., R. Arbuckle, B.H. Liu, H.W. Tao and L.I. Zhang, 2008. Lateral sharpening of cortical frequency tuning by approximately balanced inhibition. Neuron, 58: 132-143.
CrossRefDirect Link - Wilent, W.B. and D. Contreras, 2005. Dynamics of excitation and inhibition underlying stimulus selectivity in rat somatosensory cortex. Nat. Neurosci., 8: 1364-1370.
CrossRefDirect Link