
Manouchehr Safari
Research Center of Nervous System Stem Cells, Department of Anatomy, Semnan University of Medical Sciences, Semnan, Iran
Hamid Reza Sameni
Research Center of Nervous System Stem Cells, Department of Anatomy, Semnan University of Medical Sciences, Semnan, Iran
Leila Badban
Research Center of Nervous System Stem Cells, Department of Anatomy, Semnan University of Medical Sciences, Semnan, Iran
Ahmad Raze Bandegi
Department of Biochemistry, Semnan University of Medical Sciences, Semnan, Iran
Abbas Ali Vafaei
Research Center of Physiology, Semnan University of Medical Sciences, Semnan, Iran
Ali Rashidy Pour
Research Center of Physiology, Semnan University of Medical Sciences, Semnan, Iran
Laya Ghahari
Research Center of Physiology, Semnan University of Medical Sciences, Semnan, Iran
International Journal of Pharmacology
Year: 2015 | Volume: 11 | Issue: 4 | Page No.: 300-308







ABSTRACT
Propolis is a resinous natural live product derived from plant exudates collected by honey bees. It has been extensively used in folk medicine. In the present study, the neuroprotective effects of propolis was investigated in the rat model of Parkinson’s disease. The PD rats which was induced by 6-OHDA were compared with control (C) and treatment groups with different dose of Propolis. After 4 weeks of follow-up, behavioral activity, tyrosine hydroxylase positive neurons in substantia nigra pars compacta, brain derived neurotrophic factor in serum and brain and stress oxidative factors such as superoxide dismutase, glutathione peroxidase activity, malondialdehyde and ferric reducing ability of plasma in the substantia nigra were analyzed. It was confirmed that 6-OHDA led to behavioral deficits, reduction in superoxide dismutase (6.02±0.1), glutathione peroxidase activity (0.12±0.01), ferric reducing ability of plasma (0.72±0.09) and the elevation of malondialdehyde levels (6.70±0.2) in the substantia nigra. When Propolis administered prior to 6-OHDA, improved motor coordination, increased superoxide dismutase (6.66±0.1), glutathione peroxidase (0.20±0.01) and ferric reducing ability of plasma (0.76±0.01) activity and reduced the content of malondialdehyde (6.2±0.1) compared with rat treated with 6-OHDA alone. Immunohistochemical studies showed that medication of propolis could prevent the loss of tyrosine hydroxylase positive neurons induced by 6-OHDA (107±5.77 vs 63±6.58). Decrease brain derived neurotrophic factor and some stress oxidative factors has an important effect in damage of tyrosine hydroxylase neurons in the brain. Propolis improves these factors and protects tyrosine hydroxylase neurons in the brain.
PDF Abstract XML References Citation
Received: January 05, 2015;
Accepted: March 07, 2015;
Published: April 18, 2015
How to cite this article
Manouchehr Safari, Hamid Reza Sameni, Leila Badban, Ahmad Raze Bandegi, Abbas Ali Vafaei, Ali Rashidy Pour and Laya Ghahari, 2015. Protective Effects of Water Extract of Propolis on Dopaminergic Neurons, Brain
Derived Neurotrophic Factor and Stress Oxidative Factors in the Rat Model of
Parkinson’s Disease. International Journal of Pharmacology, 11: 300-308.
DOI: 10.3923/ijp.2015.300.308
URL: https://scialert.net/abstract/?doi=ijp.2015.300.308
DOI: 10.3923/ijp.2015.300.308
URL: https://scialert.net/abstract/?doi=ijp.2015.300.308
INTRODUCTION
Parkinson’s Disease (PD) is the second common neurodegenerative disease in the world (Hirtz et al., 2007). characterized by loss of dopaminergic neurons that project to the striatum in Substantia Nigra pars compacta (SNpc) leading to a decrease dopamine level in the target structure (Cooper et al., 2006; Dorsey et al., 2007; Van den Eeden et al., 2003). Loss of dopamine containing neurons in brain stem and mid brain results rigidity, motor dysfunction, tremor, postural instability, disturbance in sleep, mood and cognition (Dauer and Przedborski, 2003; Hanrott et al., 2006). One of the pathological features of PD is the presence of intracellular inclusions called Lewy bodies that consist of aggregates of the pre synaptic protein called a-synuclein (Healy et al., 2008). There are various factors that influence the pathological depositions and in general, the cause of neuronal death in neurological disorders appears to be multi factorial (Dorsey et al., 2007; Desai et al., 2010). The oxidative stress linked with PD is supported by postmortem studies and by researches that show the increased level of oxidative stress in the substantia nigra pars compacta, demonstrating thus the capacity of oxidative stress to induce nigral cell degeneration (Healy et al., 2008). 6-Hydroxydopamine 6-OHDA widely used to generate PD like models in animals (Uversky et al., 2001). Injection of this toxin into SN results in selective lesion to dopaminergic Neuron (DA) (Blum et al., 2001; Bove et al., 2005). Research has been shown that 6-OHDA is also able to increase apoptosis and cell death via caspase-dependent (with a mitochondrial release of cytochrome c) and Caspase-independent pathway (Hastings, 2009; Zhang et al., 2007).
Propolis or bee glue is used by humans since a long time. It is a resinous product derived from different leaf buds or plants exudates collected by honey bees (Mogi et al., 1999; Naik et al., 2013). It has been used for various purpose such as sealant and sterility in honeybee nests (Bankova et al., 1999) medicine (Duran et al., 2007) now it has been reported to have a wide range of biological and pharmacological activities, including anticancer (Kim et al., 2008), antibacterial (Kujumgiev et al., 1999; Liberio et al., 2009), anti-inflammatory (Hu et al., 2005), anti oxidative (Okutan et al., 2005), anti microbial (Sforcin et al., 2000), anti fungal (Banskota et al., 2001), tumoricidal (Orsolic et al., 2004). Propolis contains a wide variety of components. including flavonoids, benzoic acids, essential oils10%, resins 50-70% (Popova et al., 2009). The main components of propolis contains flavonoids, phenolics and aromatic compounds (Banskota et al., 2001). Water extract of propolis (WEP) is the natural form of propolis and because of the main constituents (caffeoyliquinic acid) have a greater antioxidative effects and better inhibitory effects against certain enzymes (Nakajima et al., 2007). Propolis is rich in polyphenol and flavonoid content (Ghasem et al., 2007). So, WEP has a highest anti oxidative activity. More researchers have found that propolis has a neuroprotective effects in several models of neurological disorders including cerebral ischemia, neural apoptosis and encephalomyelitis (Han et al., 2003; Navarro and Boveris, 2009).
The purpose of this study was to examine the neuroprotective effects of propolis on the dopaminergic neurons in midbrain and some stress oxidative factors in rat models of PD.
MATERIALS AND METHODS
Animals: Adult male albino-wistar rat weighing 200-250 g were obtained from the animal center of Iran University in Tehran, Iran. All rats were housed with food and water available ad libitum. The storage room was maintained at constant temperature and humidity on a 12 h dark and light. All procedure were carried out in accordance with the National Institutes of Health Guide for Care and Use of Laboratory Animals that authorized by Ethics Committee of Semnan Medical University Semnan, Iran. Rat were randomly divided into seven groups (n = 10). Each group was housed in a separate cages the first group received only saline as a control. The second group received saline plus ascorbic acid (vehicle of 6-OHDA as a sham). The third group received 4 μg, 6-OHDA in left substantia nigra SNpc as PD model. The forth and fifth groups, received propolis with doses of 50 (PD/Ext50) and 100 mg kg-1 (PD/Ext100) after PD for 4 weeks. The sixth and seventh groups (pretreatment) received 50 (PreT50/PD) and 100 mg kg-1 (Pre T100/PD) propolis every 24 h for 28 consecutive days before injection of 6-OHDA.
6-Hydroxydopamine lesion: To obtain unilateral lesion of nigral system, all rats received 6-OHDA into the left SN. All animals were anesthetized using ketamine hydrochloride/ xylazine hydrochloride (100-20 mg kg-1) (sigma-Aldrich) and placed into stereotaxic instrument (stoelting, USA) with nose and ear bar adapted for rat. The skull was exposed by a 2 cm incision in the scalp and a single hole was drilled over each side of the Substantia Nigra pars compacta (SNpc) at the following coordinates according to the Rat Brain in Stereotaxic Coordinates (Paxinos et al., 1997): AP = - 4.8 mm posterior to bregma , ML = -1.6 mm lateral to the midline, DV = 8.2 mm vertical from the dura and 4 μL of 6-OHDA (2 μg μL-1) dissolved in vehicle 2 mg mL-1 anti ascorbic acid (Sigma Chemical Co., St. Louis, USA) in saline was injected to the Snpc. The 6-OHDA was injected to the left side at the rate of 1 μL min-1.
Preparation of WEP: A WEP was prepared by the method described by Nakajima et al. (2007). Fresh propolis was purchased from department of agriculture beekeeping in Semnan, Iran. Briefly, 50 g propolis was cut into small pieces and then 5 volume of distilled water were added and stirred for 24 h at 45°C. The sample was also boiled for 2 h. Extracts were then filtered twice and stored in sealed bottles at 4°C until use.
Determination of total poly phenol content: Propolis extract was redissolved in 95% distilled water at a concentration of 50 mg mL-1. Water extract (0.1 mL) were diluted with 95% distilled water (0.9 mL) and mixed with 5 mL of 10 fold diluted solution of 2N Folin-Ciocalteu reagent (Sigma, UK). Four milliliter of saturated sodium carbonate solution was added to the mixture and then and then was stirred. The absorbance of the reaction mixture was measured at 765 nm after 3 h (Spanos and Wrolstad, 1990). Caffeic acid (Sigma, Germany) was used as a standard compound in the range of 100-500 μg mL-1 concentration to construct a standard curve.
Immunohistochemical and histological study: Transcardial perfusion was performed with 200 mL normal saline followed by 400 mL 4% par formaldehyde in 0.1 mmol L-1 PBS. Animals were deeply sedated (pentobarbital, 50 mg kg-1, i.p.) and perfuse Transcardial with 4% par formaldehyde (PFA). Brain stems were fixed with 4% par formaldehyde, the tissue was then cut (5-7 μm thick). The De paraffinized, next step was antigen retrieval with tris/EDTA pH 9 buffer and 3% hydrogen peroxide, blocked/permeabilized with 10% normal goat serum in PBS containing 0.3% Triton-X100, stained with a mouse anti-tyrosine hydroxylase TH polyclonal antibody (1-100 dilution, contain 10% normal goat serum abcam), followed by a biotin-conjugated goat anti-mouse IgG (1-200 dilution, abcam). After washing, sections were incubated with an avidin and biotinylated horseradish peroxidase complex followed by diaminoazobenzidine (abcam). Dopaminergic cells were visualized and quantified using Olympus microscope. Some sections were stained with hematoxylin and eosin or with cresyl fast violet.
Behavioral testing: Motor asymmetry following unilateral lesion of the nigro striatal pathway was assessed by apomorphine-induced rotational behavior. All behavioral testing was performed by an observer blinded to the group. All groups were tested for rotational behavior 6 days after the first surgery and after treatment (4 week). The rats received subcutaneously 0.5 mg kg-1 apomorphine hydrochloride (sigma) dissolved in 0.9% NaCl. Twenty minutes after injection total turns contra lateral to the lesion were counted over a period of 40 min. The value was expressed as contra lateral net turns/min.
Morphometric studies: Ten coronal sections from rostral to caudal of the SNc in each animal were analyzed. The picture of each section was taken by an Olympus AX70 microscope and DP11 digital camera under magnification of 40×. An area of 10,000 μm2 was measured randomly in the region of SNc in ten separate microscopic fields. To count the number of neuronal cells in Nissl staining (including neuronal cells and interneuron) and IHC (the TH positive cells), the pictures were transferred to the computer using OLYSIA autobiorepot software (Olympus optical co. Ltd, Japan). A grid was superimposed on the picture and the cells with obvious and normal nucleus were counted.
Measurement of brain oxidative stress markers, serum and brain BDNF concentration: At the end of experiments, after deep anesthesia serum brain-derived neurotrophic factor levels were measured with the reagents provided in the BDNF Immunoassay System and according to the manufacturer’s recommended protocols (R and D Systems, Minneapolis, MN, USA). The heart blood was initially collected in anticoagulant-free tubes and incubated at room temperature for 30 min. Next, samples were centrifuged for 15 min at 1000×g. The supernatant was collected and stored at -70°C. Then rats decapitated, brains were removed. The extraction of BDNF from the brain tissue was performed on ice and following the description of Szapacs et al. (2004). Briefly, the tissue was suspended in 5 volume of lysis buffer containing 137 mM NaCl, 20 mM Tris-HCl, 1% NP40, 15% glycerol, 0.5 mM PMSF, 0.5 mM sodium and protein inhibitor cocktail (Calbiochem, USA). The suspension was homogenized on ice for 20 min using a sonicator at power level 2. The brain homogen were then centrifuged at 16000×g for 20 min at 4°C. The resulting supernatant was stored at -70°C for further analyses. Next 5 brain stem was selected for Immunohistochemistry and Nissl staining and 5 others for analysis of stress oxidative markers such as MDA, SOD, GSH-px and FRAP.
For assessing of stress oxidative markers such as MDA, SOD, GSH-px and FRAP, after decapitation the substantia nigra was isolated. Samples were weighed accurately and prepared with 0.90% normal saline to give 10% tissue homogenate by super-audible cell disintegrator (Sonicator) which was then centrifuged at 5000 rpm for 20 min at 4°C. The supernatant was collected and kept at -70°C until use. The protein concentration of the substantia nigra was determined by the method of Bradford (Bradford, 1976). The activities of SOD, GSH-PX and the content of MDA in the substantia nigra were determined following the kit specifications (Benzie and Strain, 1999; Uchiyama and Mihara, 1978). Results are presented as units of activity per mg of protein (wet weight) or content.
Statistical analysis: All data was reported as Mean±SEM enzyme activities (expressed as OD value). Statistical analysis was carried out by computer using SPSS ver.11.5 software. For each tissue was analyzed by one-way ANOVA and Tukey post hoc multiple comparison tests. The p-values less than 0.05 were considered significant.
RESULTS
Total polyphenol content (g/100 g extract): Total polyphenol content that determined by Folin-Ciocalteu colorimetric methods was 5.8±0.03.
Drug-induced rotational behavior assessment: Contra lateral turns induced by apomorphine hydrochloride were increased in the 6-OHDA group in the first test (2 weeks after surgery) compared to treatment and control groups (p<0.01), as shown in Fig. 1. The frequency and duration of the rotation in 6-OHDA group were 7.11±0.1 circles/min. In the second test, 4 weeks after surgery the number of contra lateral turns observed in the 6-OHDA group was greater than all the other groups (7.57±0.08) (p<0.01). When each group was analyzed considering, in the pre treatment groups (50 and 100 mg kg-1) showed a significant reduction in total contra lateral turns (4.13±0.29, 3.06±0.1). On the other hand, the number of contra lateral turns decreased in propolis treatment groups but in pretreatment it was very noticeable. There were no significant differences between the first and second tests of the 6-OHDA group with the treatment groups 50 and100 mg kg-1.
TH-Immunohistochemistry: Tyrosine hydroxylase staining was performed for assessment of the survival of dopaminergic neurons. Injection of 6-OHDA into the SNpc significantly reduced the absorbance of TH immunostaining (63±6.58, p<0.001) neurons as illustrated in Fig. 2 and 3. These cells reduction was observed in all experimental groups but in pretreatment groups cell number was higher than other groups. Figure 1, show the boundaries of the SNpc which were used to measure cell number in the experimental groups in Nissl and IHC staining. Statistical and comparative light microscopic analyses demonstrated that population of Nissl and TH-stained cell number in the 6-OHDA group had a prominent decrease compared to the control and treatment groups (63±6.58, p<0.001) (Fig. 3).
The cell number in the control and sham groups were similar (132±5.59 and 28±4.96, respectively) but in the pretreatment (100 mg kg-1) group, the number of Nissl-stained cells was 107±5.77. So, it can be guessed that propolis could protect dopaminergic neurons from 6-OHDA neuro toxicity in rat.
BDNF assay: Neurotrophic factors, such as the BDNF, have been considered to play an important role in neuroprotection. In this study, In addition, It has been demonstrated that endogenous BDNF are altered in PD and then propolis can change the BDNF level in serum and brain. It was found that the amount of brain BDNF was higher and significant in pretreatment and treatment (100 mg kg-1) groups in compared with the 6-OHDA group (7840±415 and 6589±215 vs. 5696±301.79 pg/mg protein, respectively) p = 0.001 (Fig. 4), whereas in this two groups, serum BDNF content were not statistically different with the control groups (5565±472 and 5340±273.86 vs. 5296±125 pg/mg protein, respectively) (p = 0.147) (Fig. 4).
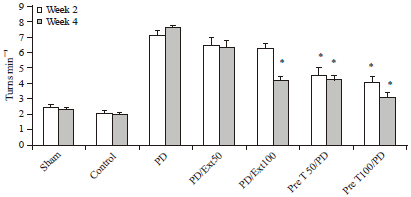 | |
| Fig. 1: | Apomorphine-induced rotations of rats over time reveal a significant reduction in the number of turns in pretreatment groups 4 weeks after surgery compared with PD rats. All values are Mean±SEM. *Compared with the PD group (p≤0.01) |
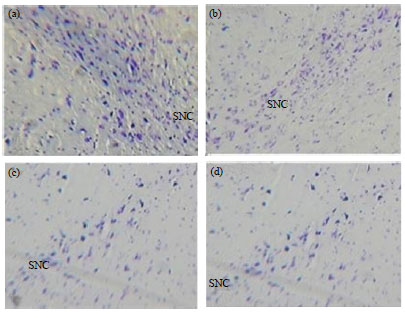 | |
| Fig. 2(a-d): | Nissl staining of substantia nigra of rat photomicrographs were taken at a magnification of 200 (a) Sham, (b) Pre treatment (100 mg kg-1), (c) 6-OHDA and (d) Propolis (100 mg kg-1) |
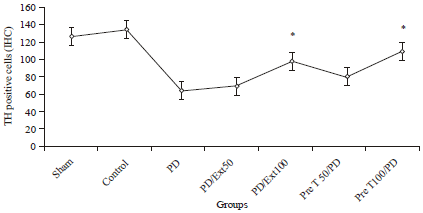 | |
| Fig. 3: | Immunohistochemical staining of Tyrosine Hydroxylase (TH) cells in substantia nigra of rats. All values are Mean±SEM for 3 rats per group and 5 sections per rat, *Compared with the PD group (p≤0.01) |
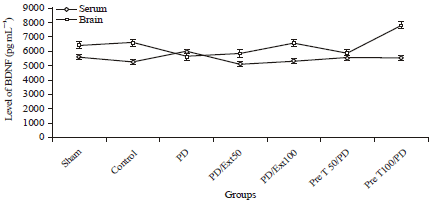 | |
| Fig. 4: | Levels of BDNF in the serum and brain of rats. In serum Ratio was significantly elevated by 6-OHDA treatment in comparison with the control group. At the same time it was induced in the propolis treatment. All values are Mean±SEM |
Effects of propolis on the activity of SOD, GSH-Px and MDA levels: GSH-Px is the most important antioxidant defense molecule and SOD is an antioxidant enzyme responsible for dismutation of superoxide radicals. The effects of propolis on the activity of SOD, GSH-Px and the content of MDA in the substantia nigra of rats are shown in Table 1. 6-OHDA administration in the PD model of rats causes a significant reduction in SOD and GSH-Px activity and increased level of MDA relative to the control group (p≤0.01). However, these conditions are significant in the pre-treatment groups. The activities of SOD in either propolis groups were increased but in the pre-treatment group (100 mg kg-1) was significant (6.66±0.1 p≤0.01 for all comparisons). The GSH-Px activity as well as SOD was increased in the propolis groups but in pre-treatment (100 mg kg-1) and treatment (100 mg kg-1) vs. 6-OHDA group was significant (0.20±0.01 and 0.16±0.01vs. 0.12±0.01, p≤0.01). The level of MDA was decreased in propolis groups and was significant in pre-treatment (100 mg kg-1) ant (treatment 100 mg kg-1) groups in contrast to 6-OHDA and even other groups. (6.20±0.1 and 6.24±0.2 vs. 6.70±0.2 p≤0.01).
Effects of propolis on the FRAP: As shown in Table 1, 6-OHDA treatment induced a marked decrease in FRAP activity (0.72±0.09, p≤0.01). 6-OHDA, the well known antioxidant inhibitor, exerted a strong effect on FRAP inhibition.
| Table 1: | Activity of Superoxide dismutase (SOD), Glutathione peroxidase (GSH-Px), Malondialdehyde (MDA) and the content of Ferric Reducing Ability of Plasma (FRAP) |
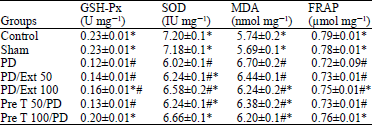 | |
| Values are presented as Mean±SEM (n = 5), *Compared with the PD group (p≤ 0.01), #Compared with the control group (p≤0.01) | |
However, pre-treatment and treatment with propolis at the dose of 100 mg kg-1 significantly affect the activity of FRAP (0.76±0.01 and 0.75±0.01 p≤0.05), therefore, suggesting that the neuroprotective effects of propolis may be due to the FRAP activity.
DISCUSSION
This study demonstrates that Water Extract of Propolis (WEP) and its main constituent, polyphone content, can protect dopaminergic neurons from 6-OHDA induced cell death and moreover these neuroprotective effects were paralleled by the same factors antioxidant effects. Oxidative damage is strongly involved in the pathogenesis of neurodegenerative diseases such as Huntington’s (Beal, 1998), Alzheimer’s (Hensley et al., 1995) and Parkinson’s (Adams et al., 2001). A number of studies have shown that in PD the dopaminergic neurons exists in a state of constant oxidative stress, in large part because of generation of H2O2 (Sherer et al., 2002; Beal, 1998).
The neurotoxin 6-OHDA is known to cause Parkinson in rat. The behavioral protests, neurochemical features and primary pathological condition induced by 6-OHDA in rat are similar to PD patients. Therefore, 6-OHDA treated rat make well conventional models and this product is widely used for studies on PD (Cutillas et al., 1999; Ferger et al., 2001). In the present study, 6-OHDA-lesioned rats showed a behavioral deficit, during the first week, the average body weight was slightly decreased. Propolis pre-treated rat (50-100 mg kg-1) exhibited increased motor coordination compared with 6-OHDA treated rat alone.
The composition of propolis depends upon the vegetation of the area from where it was collected. Colorimetric methods are convenient and appropriate for routine analysis of phenolics (Spanos and Wrolstad, 1990). Folin-Ciocalteu colorimetric methods have been applied to determine total polyphenol contents of crude water extracts of propolis samples. This may be helpful in evaluation of propolis quality and its properties. Caffeic acid was employed as a standard compound for estimation of total polyphone contents because it is one of the most abundant phenolic acids found in propolis. Antioxidative activity was demonstrated in propolis it is proposed that strong antioxidative activity occurs in propolis with high amounts of phenolic compounds and weak activity with low amounts. Total polyphone content of water extracts of this sample was (5.8 g/100 g±0.03). According to the other studies total polyphenol content of this extract is higher than. Propolis is believed to protection against 6-OHDA neurotoxicity via., antioxidant activity and free radical scavenging mechanism in the brain. Its neuroprotective action was due to the inhibition of the metabolism of dopamine in the brain by dopamine reuptake inhibition, stimulation of antioxidant enzyme, such as SOD and GSH-Px and increased turnover of dopamine (Kahle et al., 2009). Therefore, because of higher polyphenolic compounds in semnan propolis this study used as a protective factor for the dopaminergic neurons in the brain stem.
Dopamine is the main primary neurotransmitter that involved in motor functions; its loss directly implicated physical movements and contributes to the clinical symptoms, depletion of dopamine cause PD in humans or in animal models of the disease. 6-OHDA causes a partial lesion of the substantia nigra and a significant reduction in striatal dopamine levels (Zhang et al., 2007). .Factors that are able to improve 6-OHDA induced neuronal damage are considered to be neuroprotector. The results of the present study show that the pre-intake of propolis markedly improved 6-OHDA-induced dopamine and dopaminergic neurons in the striatum. The enhancement of dopamine content by propolis might have restored the changes in locomotors and rotary activity. The neuroprotective effect of propolis may be due to an increase in dopamine metabolism in the striata.
Dopamine is synthesized in two steps. Firstly, tyrosine is converted to L-dihydroxyphenylalanine (L-DOPA) by TH and then L-DOPA is then converted to dopamine by L-DOPA decarboxylase. Tyrosine hydroxylase is, therefore, a key enzyme for dopamine biosynthesis and is used as a marker for dopaminergic neurons (Ishikawa et al., 2010). The present results show that administration of propolis reduced the 6-OHDA induced loss of tyrosine hydroxylase-positive neurons in the rat substantia nigra which suggest that propolis may enhance the survival of dopamine neurons in lesion site.
To further investigate the mechanism of action of propolis, the anti-oxidative capacity and the activity of FRAP was also measured. Oxidative stress refers to the cytological consequences of a mismatch between the production of free radicals and the ability of the cell to defend against them. A defect naturally occurring antioxidant defenses could lead to neurodegeneration in PD. This imbalance results in apoptosis of neurons and auto-oxidation of dopamine (Khullar et al., 2010). The SOD catalyzes the dismutation of superoxide (O‾2) to hydrogen peroxide H2O2 which protect against oxidative injury. GPx located in mitochondrion, convert H2O2 to H2O. Thus it showed the role of protecting cellular structures (Hastings, 2009). Current studies show a decrease in the activities of SOD and GSH-PX in the 6-OHDA groups. However, the decrease of antioxidant enzyme activities caused by 6-OHDA was markedly restored by treatment and pre-treatment with propolis. Meanwhile, the level of MDA which serves as an index for determining the extent of lipid per oxidation, was reduced by propolis which was altered by 6-OHDA. Restoration of the activities of SOD, GSH-Px, FRAP and reduction of the content of MDA due to pre-treatment with propolis demonstrate the protective and anti oxidant role of propolis.
Present study showed that the activity of SOD and GSH-Px substantia nigra were induced compared with 6-OHDA group. Under normal physiological conditions, free radicals produced by metabolism can be inactivated by free radicals scavenging system such as catalase, GSH-PX and SOD.
There are two possible explanations for the induction of GSH-Px activity: (1) The increase in GSH-Px activity is elicited by the antioxidative activity of propolis which protects the enzyme from inactivation by hydroxyl radicals and (2) Enzyme induction is mediated by changes in GSH-Px gene expression. According with these theories, our studies showed higher activities of GSH-Px in the pretreatment ant treatment group of propolis and protect neurons from damages.
Striatal neurons depend on BDNF for function and survival which is chiefly provided by anterograde transport from corticostriatal afferents (Baquet et al., 2004). The present study reports that elevation of brain and serum BDNF following 6-OHDA treatment support dopaminergic neurons of SNpc. One of the mechanisms that propolis might protect striatal neurons is through the production of neurotrophins such as BDNF. High concentrations of BDNF have not only been found in the central nervous system but also in non neuronal cells, particularly in platelets. A positive correlation between serum and cortical BDNF concentrations has been observed in rats (Karege et al., 2002) and humans (Weickert et al., 2003). In this study, level of BDNF was increased in both serum and brain in pretreatment group (100 mg kg-1).
We think that there was a constitutive up-regulation of serum BDNF concentrations might compensate for defective intracellular protein signaling in the SNpc of brain stem.
Post mortem study of brain tissue from patients with Parkinson’s disease suggests that striatal neurodegeneration correlates with reductions in BDNF expression. Moreover, in brain tissue from patients with Parkinson’s (Jenner et al., 1992; Yasui et al., 2006) or with Lewy body dementia, striatal BDNF immunoreactivity is reduced as compared to age- and sex-matched controls.
Now, there is no efficient treatment for PD patients (Kitajima, 2008). A great number of clinical trials, testing pharmacological approaches, have so far produced disappointing results. Neuroprotective strategies using several growth factors have also been proposed for PD (Weickert et al., 2003). Among them, BDNF, so far the best candidate, has been shown to regulate the onset and Severity of motor and cognitive functions in PD mouse models (Karege et al., 2002). And participate in the disruption of the cortico-striatal glutamatergic transmission.
In animal models, a striatal stab wound in 6-8 week old mice increases BDNF level around the injury site and appear to be localized to activated microglia and macrophages (Braak et al., 2004). Further, production of BDNF occurs in concert with sprouting dopaminergic fibers and dopamine-transporter positive neuritis (Collier et al., 2005) thus these studies suggest that increased BDNF expression following administration of propolis support dopaminergic neurons.
CONCLUSION
Our data provide evidence that propolis has neuroprotective effects in 6-OHDA-induced PD rat which may have contributed to its anti-oxidative capacity to scavenge free radicals and resulted in the survival of more dopamine neurons. We propose that propolis has shown anti-parkinsonian properties, pending future studies to elucidate further detailed mechanisms of action.
ACKNOWLEDGMENT
This study was supported by a Grant from the Semnan University of Medical Sciences (No. 405).
REFERENCES
- Adams, Jr. J.D., M.L. Chang and L. Klaidman, 2001. Parkinsons disease-redox mechanisms. Curr. Med. Chem., 8: 809-814.
CrossRefDirect Link - Bankova, V., G. Boudourova-Krasteva, J.M. Sforcin, X. Frete, A. Kujumgiev, R. Maimoni-Rodella and S. Popov, 1999. Phytochemical evidence for the plant origin of Brazilian propolis from Sao Paulo state. Zeitschrift Naturforschung C, 54: 401-405.
PubMed - Banskota, A.H., Y. Tezuka and S.H. Kadota, 2001. Recent progress in pharmacological research of propolis. Phytoter. Res., 15: 561-571.
CrossRefPubMedDirect Link - Baquet, Z.C., J.A. Gorski and K.R. Jones, 2004. Early striatal dendrite deficits followed by neuron loss with advanced age in the absence of anterograde cortical brain-derived neurotrophic factor. J. Neurosci., 24: 4250-4258.
CrossRefDirect Link - Beal, M.F., 1998. Mitochondrial dysfunction in neurodegenerative diseases. Biochimica Biophysica Acta (BBA)-Bioenerget., 1366: 211-223.
CrossRefDirect Link - Benzie, I.F.F. and J.J. Strain, 1999. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Method Enzymol., 299: 15-27.
CrossRefPubMedDirect Link - Blum, D., S. Torch, N. Lambeng, M.F. Nissou, A.L. Benabid, R. Sadoul and J.M. Verna, 2001. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: Contribution to the apoptotic theory in Parkinson's disease. Prog. Neurobiol., 65: 135-172.
CrossRefDirect Link - Bove, J., D. Prou, C. Perier and S. Przedborski, 2005. Toxin-induced models of Parkinson's disease. NeuroRx, 2: 484-494.
CrossRefDirect Link - Braak, H., E. Ghebremedhin, U. Rub, H. Bratzke and K.D. Tredici, 2004. Stages in the development of Parkinson's disease-related pathology. Cell Tissue Res., 318: 121-134.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Collier, T.J., Z.D. Ling, P.M. Carvey, A. Fletcher-Turner, D.M. Yurek, J.R. Sladek Jr. and J.H. Kordower, 2005. Striatal trophic factor activity in aging monkeys with unilateral MPTP-induced parkinsonism. Exp. Neurol., 191: 60-67.
CrossRefDirect Link - Cooper, A.A., A.D. Gitler, A. Cashikar, C.M. Haynes and K.J. Hill et al., 2006. α-synuclein blocks ER-golgi traffic and rab1 rescues neuron loss in parkinson's models. Science, 313: 324-328.
CrossRefDirect Link - Cutillas, B., M. Espejo, J. Gil, I. Ferrer and S. Ambrosio, 1999. Caspase inhibition protects nigral neurons against 6‐OHDA‐induced retrograde degeneration. Neuroreport, 10: 2605-2608.
Direct Link - Dauer, W. and S. Przedborski, 2003. Parkinson's disease: Mechanisms and models. Neuron, 39: 889-909.
CrossRefPubMedDirect Link - Desai, K.M., T. Chang, H. Wang, A. Banigesh and A. Dhar et al., 2010. Oxidative stress and aging: Is methylglyoxal the hidden enemy? Can. J. Physiol. Pharmacol., 88: 273-284.
CrossRefDirect Link - Dorsey, E.R., R. Constantinescu, J.P. Thompson, K.M. Biglan and R.G. Holloway et al., 2007. Projected number of people with Parkinson disease in the most populous nations, 2005 through 2030. Neurology, 68: 384-386.
PubMed - Duran, N., P.D. Marcato, C.M. Buffo, M.M. de Azevedo and E. Esposito, 2007. Poly (epsilon-caprolactone)/propolis extract: Microencapsulation and antibacterial activity evaluation. Pharmazie, 62: 287-290.
PubMed - Ferger, B., S. Rose, A. Jenner, B. Halliwell and P. Jenner, 2001. 6-hydroxydopamine increases hydroxyl free radical production and DNA damage in rat striatum. Neuroreport, 12: 1155-1159.
Direct Link - Ghasem, Y.B., O. Abdolghaffar and M. Hasanloei, 2007. Antibacterial and antifungal activity of Iranian propolis against Staphylococcus aureus and Candida albicans. Pak. J. Biol. Sci., 10: 1343-1345.
CrossRefPubMedDirect Link - Han, B.S., H.S. Hong, W.S. Choi, G.J. Markelonis, T.H. Oh and Y.J. Oh, 2003. Caspase-dependent and -independent cell death pathways in primary cultures of mesencephalic dopaminergic neurons after neurotoxin treatment. J. Neurosci., 23: 5069-5076.
Direct Link - Hanrott, K., L. Gudmunsen, M.J. O'Neill and S. Wonnacott, 2006. 6-Hydroxydopamine-induced apoptosis is mediated via extracellular auto-oxidation and caspase 3-dependent activation of protein kinase Cδ. J. Biol. Chem., 281: 5373-5382.
CrossRef - Hastings, T.G., 2009. The role of dopamine oxidation in mitochondrial dysfunction: Implications for Parkinson's disease. J. Bioenerg. Biomembr., 41: 469-472.
CrossRefDirect Link - Healy, D.G., M.O. Falchi, S.S. O'Sullivan, V. Bonifati and A. Durr et al., 2008. Phenotype, genotype and worldwide genetic penetrance of LRRK2-associated Parkinson's disease: A case-control study. Lancet Neurol., 7: 583-590.
CrossRefDirect Link - Hensley, K., N. Hall, R. Subramaniam, P. Cole and M. Harris et al., 1995. Brain regional correspondence between Alzheimer's disease histopathology and biomarkers of protein oxidation. J. Neurochem., 65: 2146-2156.
CrossRefDirect Link - Ishikawa, S., T. Taira, K. Takahashi-Niki, T. Niki, H. Ariga and S.M.M. Iguchi-Ariga, 2010. Human DJ-1-specific transcriptional activation of tyrosine hydroxylase gene. J. Biol. Chem., 285: 39718-39731.
CrossRefDirect Link - Jenner, P., A.H.V. Schapira and C.D. Marsden, 1992. New insights into the cause of Parkinson's disease. Neurology, 42: 2241-2250.
Direct Link - Kahle, P.J., J. Waak and T. Gasser, 2009. DJ-1 and prevention of oxidative stress in Parkinson's disease and other age-related disorders. Free Radical Biol. Med., 47: 1354-1361.
CrossRefDirect Link - Karege, F., M. Schwald and M. Cisse, 2002. Postnatal developmental profile of brain-derived neurotrophic factor in rat brain and platelets. Neurosci. Lett., 328: 261-264.
CrossRefDirect Link - Khullar, M., A.A.S. Al-Shudiefat, A. Ludke, G. Binepal and P.K. Singal, 2010. Oxidative stress: A key contributor to diabetic cardiomyopathy. Can. J. Physiol. Pharmacol., 88: 233-240.
CrossRefDirect Link - Kim, D.M., G.D. Lee, S.H. Aum and H.J. Kim, 2008. Preparation of propolis nanofood and application to human cancer. Biol. Pharmaceut. Bull., 31: 1704-1710.
CrossRefDirect Link - Kitajima, S., 2008. Hydrogen peroxide-mediated inactivation of two chloroplastic peroxidases, ascorbate peroxidase and 2-cys peroxiredoxin. Photochem. Photobiol., 84: 1404-1409.
CrossRefDirect Link - Kujumgiev, A., I. Tsvetkova, Y. Serkedjieva, V. Bankova, R. Christov and S. Popov, 1999. Antibacterial, antifungal and antiviral activity of propolis of different geographic origin. J. Ethnopharmacol., 64: 235-240.
CrossRefPubMedDirect Link - Liberio, S.A., A.L.A. Pereira, M.J.A.M. Araujo, R.P. Dutra and F.R.F. Nascimento et al., 2009. The potential use of propolis as a cariostatic agent and its actions on mutans group streptococci. J. Ethnopharmacol., 125: 1-9.
CrossRefDirect Link - Uchiyama, M. and M. Mihara, 1978. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem., 86: 271-278.
CrossRefDirect Link - Mogi, M., A. Togari, T. Kondo, Y. Mizuno and O. Komure et al., 1999. Brain-derived growth factor and nerve growth factor concentrations are decreased in the substantia nigra in Parkinson's disease. Neurosci. Lett., 270: 45-48.
CrossRefDirect Link - Naik, D.G., H.S. Vaidya and T.P. Namjoshi, 2013. Essential oil of Indian propolis: Chemical composition and repellency against the honeybee Apis florea. Chem. Biodivers., 10: 649-657.
CrossRefDirect Link - Nakajima, Y., M. Shimazawa, S. Mishima and H. Hara, 2007. Water extract of propolis and its main constituents, caffeoylquinic acid derivatives, exert neuroprotective effects via antioxidant actions. Life Sci., 80: 370-377.
CrossRefDirect Link - Navarro, A. and A. Boveris, 2009. Brain mitochondrial dysfunction and oxidative damage in Parkinson's disease. J. Bioenerg. Biomembr., 41: 517-521.
CrossRefDirect Link - Okutan, H., N. Ozcelik, H.R. Yilmaz and E. Uz, 2005. Effects of caffeic acid phenethyl ester on lipid peroxidation and antioxidant enzymes in diabetic rat heart. Clin. Biochem., 38: 191-196.
CrossRefDirect Link - Orsolic, N., A.H. Knezevic, L. Sver, S. Terzic and I. Basic, 2004. Immunomodulatory and antimetastatic action of propolis and related polyphenolic compounds. J. Ethnopharmacol., 94: 307-315.
CrossRefPubMedDirect Link - Popova, M.P., I.B. Chinou, I.N. Marekov and V.S. Bankova, 2009. Terpenes with antimicrobial activity from Cretan propolis. Phytochemistry, 70: 1262-1271.
CrossRefDirect Link - Sforcin, J.M., A. Fernandes Jr., C.A.M. Lopes, V. Bankova and S.R.C. Funari, 2000. Seasonal effect on Brazilian propolis antibacterial activity. J. Ethnopharmacol., 73: 243-249.
CrossRefPubMedDirect Link - Sherer, T.B., R. Betarbet, A.K. Stout, S. Lund and M. Baptista et al, 2002. An in vitro model of Parkinson's disease: Linking mitochondrial impairment to altered α-synuclein metabolism and oxidative damage. J. Neurosci., 22: 7006-7015.
PubMed - Spanos, G.A. and R.E. Wrolstad, 1990. Influence of processing and storage on the phenolic composition of Thompson Seedless grape juice. J. Agric. Food Chem., 38: 1565-1571.
CrossRefDirect Link - Szapacs, M.E., T.A. Mathews, L. Tessarollo, W.E. Lyons, L.A. Mamounas and A.M. Andrews, 2004. Exploring the relationship between serotonin and brain-derived neurotrophic factor: Analysis of BDNF protein and extraneuronal 5-HT in mice with reduced serotonin transporter or BDNF expression. J. Neurosci. Methods, 140: 81-92.
CrossRefDirect Link - Uversky, V.N., J. Li and A.L. Fink, 2001. Pesticides directly accelerate the rate of α-synuclein fibril formation: A possible factor in Parkinson's disease. FEBS Lett., 500: 105-108.
CrossRefDirect Link - Van den Eeden, S.K., C.M. Tanner, A.L. Bernstein, R.D. Fross, A. Leimpeter, D.A. Bloch and L.M. Nelson, 2003. Incidence of Parkinson's disease: Variation by age, gender and race/ethnicity. Am. J. Epidemiol., 157: 1015-1022.
Direct Link - Weickert, C.S., T.M. Hyde, B.K. Lipska, M.M. Herman, D.R. Weinberger and J.E. Kleinman, 2003. Reduced brain-derived neurotrophic factor in prefrontal cortex of patients with schizophrenia. Mol. Psychiatry, 8: 592-610.
CrossRefDirect Link - Yasui, K., Y. Inoue, T. Kanbayashi, T. Nomura, M. Kusumi and K. Nakashima, 2006. CSF orexin levels of Parkinson's disease, dementia with Lewy bodies, progressive supranuclear palsy and corticobasal degeneration. J. Neurol. Sci., 250: 120-123.
CrossRefDirect Link - Zhang, D., J.J. Zhang and G.T. Liu, 2007. The novel squamosamide derivative FLZ protects against 6-hydroxydopamine-induced apoptosis through inhibition of related signal transduction in SH-SY5Y cells. Eur. J. Pharmacol., 561: 1-6.
CrossRefDirect Link - Hu, F., H.R. Hepburn, Y. Li, M. Chen, S.E. Radloff and S. Daya, 2005. Effects of ethanol and water extracts of propolis (bee glue) on acute inflammatory animal models. J. Ethnopharmacol., 100: 276-283.
CrossRefDirect Link