
Ruth Chu
Department of Biology, Gilead Sciences, Inc., 7601 Dumbarton Circle, Fremont, California, 94555, USA
Ying Ge
Department of Biology, Gilead Sciences, Inc., 7601 Dumbarton Circle, Fremont, California, 94555, USA
Ming Yang
Department of Biology, Gilead Sciences, Inc., 7601 Dumbarton Circle, Fremont, California, 94555, USA
International Journal of Pharmacology
Year: 2015 | Volume: 11 | Issue: 4 | Page No.: 291-299







ABSTRACT
KB-R7943, a Na+/Ca+ exchanger 1 (NCX1) inhibitor, was found to enhance Glucose-Stimulated Insulin Secretion (GSIS) from human and mouse pancreatic islets through inhibiting the forward mode of NCX1. This report studied the effects of KB-R7943 and a different NCX1 inhibitor SN-6, on GSIS from INS-1E cells, a rat pancreatic β-cell line and the potential mechanisms of action. It was found that KB-R7943 significantly enhanced GSIS from INS-1E cells in a concentration-dependent manner. In contrast, no significant effect was observed for SN-6 on GSIS. Similarly, KB-R7943 but not SN-6, enhanced glipizide, a sulfonylurea, stimulated insulin secretion from INS-1E cells. In addition, KB-R7943 but not SN-6, significantly enhanced glipizide-stimulated increase of intracellular Ca2+ ([Ca2+]i). Transient transfection with an NCX1 siRNA resulted in 70 and 62% knockdown of NCX1 gene and protein expression, respectively, in INS-1E cells. Surprisingly, knockdown of NCX1 had little impact on KB-R7943-induced enhancement of glucose- or glipizide-stimulated insulin secretion. Similarly knockdown of NCX1 did not affect KB-R7943 mediated enhancement of glipizide-stimulated increase of [Ca2+]i. Data suggest that the enhancement effect of KB-R7943 on GSIS in INS-1E cells most likely is mediated through an NCX1-independent pathway.
PDF Abstract XML References Citation
Received: December 13, 2014;
Accepted: February 27, 2015;
Published: April 18, 2015
How to cite this article
Ruth Chu, Ying Ge and Ming Yang, 2015. KB-R7943 Increases Glucose-Stimulated Insulin Secretion from INS-1E Cells through an NCX1-Independent Pathway. International Journal of Pharmacology, 11: 291-299.
DOI: 10.3923/ijp.2015.291.299
URL: https://scialert.net/abstract/?doi=ijp.2015.291.299
DOI: 10.3923/ijp.2015.291.299
URL: https://scialert.net/abstract/?doi=ijp.2015.291.299
INTRODUCTION
Calcium (Ca2+) is a signaling molecule which plays an important role as a second messenger. Changes of intracellular free Ca2+ ([Ca2+]i) levels regulate many cellular functions, including secretion, contraction-relaxation, cell-cycle progression and apoptosis, etc. In pancreatic β-cells, glucose and sulfonylureas increase [Ca2+]i levels through inhibition of KATP channels, followed by activation of L-type Ca2+ channels. The increase of [Ca2+]i leads to insulin granule exocytosis (MacDonald et al., 2005; Straub and Sharp, 2002). In addition to KATP and Ca2+ channels, many other membrane proteins also participate in the regulation of [Ca2+]i in pancreatic β-cells, including the Na+/Ca+ exchanger 1.
The Na+/Ca+ exchanger (NCX) is a bidirectional ion exchanger located at the plasma membrane that controls the exchange of Na+ and Ca2+ in either Ca2+ extrusion (forward) or Ca2+ influx (reverse) mode. Four isoforms of NCX have been cloned, namely NCX1, NCX2, NCX3 and NCX4 (Herchuelz et al., 2013). The NCX1 isoform is ubiquitously expressed in different cell types from different organs (Ottolia and Philipson, 2013). In cardiomyocytes, NCX1 plays a critical role in Ca2+ extrusion through forward mode to restore basal [Ca2+]i levels between heartbeats (Bers et al., 1996). During cardiac ischemia, NCX1 also operates Ca2+ influx through reverse mode that induces undesired increase of [Ca2+]i, leading to [Ca2+]i overload. It was reported that NCX1 also participates in regulating [Ca2+]i and insulin release in pancreatic β-cells (Herchuelz et al., 2007; Hamming et al., 2010; Van Eylen et al., 1998; Herchuelz et al., 2002). However, the role of NCX1 in Glucose-Stimulated Insulin Secretion (GSIS) has been controversial. Van Eylen et al. (1998) discovered that knockdown of NCX1 in β-cells could reduce sulfonylurea or potassium induced Ca2+ influx through the reverse mode, leading to decrease of [Ca2+]i. They also found that knockdown of NCX1 could reduce the rate of [Ca2+]i decrease after removal of sulfonylurea or potassium, therefore maintain the [Ca2+]i levels. On other hand, heterozygous inactivation of NCX1 was reported to lead to increases of both GSIS and β-cell mass (Herchuelz et al., 2013; Nguidjoe et al., 2011). Similarly, it was also reported that blockade of NCX1 forward mode by KB-R7943 could retain [Ca2+]i through reducing Ca2+ extrusion, leading to increase of GSIS in islets (Hamming et al., 2010).
Several benzyloxyphenyl derivatives, SEA0400, SN-6 and KB-R7943, have been developed as selective NCX inhibitors (Iwamoto et al., 2007; Amran et al., 2003; Iwamoto, 2004). These NCX inhibitors are more potent in inhibiting the reverse mode vs. forward mode for NCX1 and subsequently prevent [Ca2+]i overload in cardiomyocytes. In pancreatic β-cells, it was reported that KB-R7943 is also very potent in blocking forward mode of NCX1, leading to sustained high level of [Ca2+]i in the presence of high glucose or sulfonylureas (Hamming et al., 2010). However, KB-R7943 is much less selective among three benzyloxyphenyl derivatives (Annunziato et al., 2004), it also blocks several other types of channels and receptors, etc. (Kraft, 2007; Ouardouz et al., 2005; Cheng et al., 2012; Terracciano and Hancox, 2013; Brustovetsky et al., 2011; Pintado et al., 2000; Barrientos et al., 2009; Santo-Domingo et al., 2007) and activates large conductance Ca2+-activated K+ (BK) channels (Liang et al., 2008a).
This study investigated the role of NCX1 in the regulation of insulin secretion in INS-1E cells by using two NCX1 inhibitors KB-R7943 and SN-6 and silencing NCX1.
MATERIALS AND METHODS
Reagents and materials: KB-R7943 (2-[4-[(4-nitrophenyl) methoxy]phenyl]ethyl ester, methanesulfonate (1:1), Carbamimidothioic acid), SN-6 (2-[[4-[(4-Nitrophenyl) methoxy]phenyl]methyl]-4-thiazolidinecarboxylic acid ethyl ester), Glipizide and exendin-4 were purchased from Sigma Aldrich Corp. (St. Louis, MO). Fetal bovine serum was purchased from Mediatech (Manassas, VA). RNA miniprep kits were purchased from Stratagene (LA Jolla, CA), iScript Reverse Transcription kits were purchased from Bio-Rad Laboratories (Hercules, CA) and SYBR Green PCR kits were obtained from Life Technologies, Inc (Grand Island, NY). Antibody against NCX1 was purchased from R and D Systems, Inc. (Minneapolis, MN). Antibody against β-actin was obtained from Cell Signalling Technology (Boston, MA). All reagents for cell culture were purchased from Life Technologies, Inc. FLIPR Calcium 4 assay kit was purchased from Molecular Devices (Sunnyvale, CA). Primers for rat NCX1, SUR1 and GLP-1R were designed using Beacon Designer 7.0 (PREMIER Biosoft International, Palo Alto, CA). Silencer Select siRNA for rat NCX1 was purchased from Life Technologies, Inc (sense sequence: GUGCGUAUCUGGAAUGAGATT, anti-sense sequence: UCUCAUUCCAGAUACGCACTG). Primers were synthesized by Eurofins MWG Operon (Huntsville, AL). The primers sequences are as follows: NCX1, forward: 5’-GGAAGATGACGATGATGATGAATG-3’, reverse: 5’-ATGAGGATGGAGACAATGAAGC-3’. Rat β-actin, forward: 5-TTCAACACCCCAGCCATGT-3’, reverse: 5’-AGTGGTACGACCAGAGGCATACA-3’. Sulfonylurea receptor type 1 (SUR1), forward: 5'-GTGCCCCTCAAGGTCGTAAAC-3, reverse: 5'-GAAGTTGTCAGCGTCTCCATCC-3. Glucagon-like peptide-1 receptor (GLP-1R), forward: 5'-ACTGGTTGCTGGTGGAAG-3, reverse: 5'-TCGTAGAGATACTTGACAATGC-3
Cell culture: INS-1E cells (passage 70-95) were cultured in RPMI 1640 medium supplemented with 11 mM glucose, 10% Fetal Bovine Serum (FBS), 1% penicillin/streptomycin /glutamine, 10 mM HEPES, 1 mM sodium pyruvate, 50 μM β-mercaptoethanol (optional). Cells were subcultured every 3-4 days.
Insulin secretion assay for INS-1E cells: INS-1E cells were seeded in 96-well plates at a density of 5×104 cells/well and cultured at 37°C in room air/5% CO2 for 2-3 days. The cell medium was replaced with Krebs-Ringer buffer (129 mM NaCl, 4.8 mM KCl, 2.5 mM CaCl2, 1.2 mM MgSO4, 1.2 mM KH2PO4, 5 mM NaHCO3 and 10 mM HEPES, pH 7.4) containing 0.1% BSA (fatty acid free) and 3 mM glucose for 30 min. Cells were then treated with agents in Krebs-Ringer buffer with different concentrations of glucose (3 mM or 11 mM glucose) and agents (indicated otherwise) for 1 h. Supernatants (100 μL) were harvested and stored at -80oC until analysis. Insulin levels in supernatant were measured using an ELISA kit from Millipore (Billerica, MA) or a Homogenous Time-Resolved Fluorescence (HTRF) assay from Cisbio Assays (Bedford, MA).
Ca2+ Mobilization assay: INS-1E cells were seeded in 96-well plates at a density of 7.0×104/well and cultured overnight. Cell medium was then replaced with Calcium 4 reagent in HBSS with 10 mM HEPES, 1% FBS and 2 mM probenecid in 100 μL and incubated for 40 min in CO2 incubator. Cells were then challenged with agents and fluorescence activity was measured using a FlexStation II. [Ca2+]i concentration is reported as relative fluorescence units (maximum increase of relative fluorescence units above basal) Data is presented as Mean±SEM.
Knockdown of NCX1 by transient transfection with siRNAs: Silencer Select siRNAs were pre-designed by Life Technolologies. INS-1E cells were transiently transfected with scrambled, NCX1 specific siRNAs at 20 nM concentration, using Lipofectamine 2000 regent according to the instruction from manufacturer ADDIN{ ADDIN EN.CITE.DATA (Elbashir et al., 2001). Forty eight hour later, cells were harvested either for quantitative real-time RT-PCR or Western blot analysis.
Quantitative Real-time RT-PCR: Total RNA was extracted using a RNA miniprep kit. cDNA was then synthesized using an iScript Reverse Transcription kit. Quantitative Real-time RT-PCR (qPCR) was performed using SYBR Green PCR reagents on Stratagene M×3000P (Stratagene, La Jolla, CA). Relative mRNA levels were calculated based on the delta Ct values (threshold cycle time) normalized to the levels of rat β-actin.
Western blot analysis: Proteins from whole cell extracts were separated on 8% acrylamide SDS-PAGE gels by electrophoresis at 50 mA. Proteins were then electrotransfered to nitrocellulose membranes with an i-Blot (Life Technologies). The membrane was pretreated with 5% non-fat milk in TTBS (20 mM Tris-HCl, pH 7.5, 120 mM NaCl, 0.05% Tween-20) for 1-2 h at room temperature. Incubation with primary antibodies (anti-NCX1 and anti-β-actin) was done at 4°C in TTBS with 5% BSA for 16 h. The membrane was then washed for 10 min, 3 times with TTBS and incubated with horseradish peroxidase-conjugated secondary antibodies for 1 h at room temperature. After 3 washes with TTBS, the bound antibody was detected by enhanced chemiluminescence (Pierce Biotechnology, Inc., Rockford, IL) with a VersaDoc Image System (Bio-Rad). Protein intensity was measured by using Quantity One software (Bio-Rad).
RESULTS
KB-R7943 but not SN-6, enhanced both glucose-and glipizide-stimulated insulin secretion from INS-1E cells: Results of qPCR showed that INS-1E cells express NCX1 at a reasonable level compared to the highly expressed sulfonylurea receptor 1 (SUR1) and glucagon-liked peptide -1 receptor (GLP-1R) (Fig. 1). Therefore, INS-1E can be used as a cell line model to study the role of NCX1 in insulin secretion. Two commercially available NCX1 inhibitors, KB- R7943 and SN-6, were tested for their effects on GSIS. In the presence of 11 mM glucose, KB-R7943 significantly increased GSIS in a concentration-dependent manner (Fig. 2a). Similar effect of KB-R7943 on GSIS was observed in the presence of 5 mM but not 3 mM glucose. In contrast, no significant effect of SN-6 on GSIS was observed in the presence of 11 mM glucose (Fig. 2b). Similarly, KB-R7943 but not SN-6 significantly increased 10 μM glipizide-stimulated insulin secretion from INS-1E cells (Fig. 2c-d).
KB-R7943 but not SN-6, concentration-dependently increased [Ca2+]i in the presence of 10 μM glipizide in INS-1E cells: [Ca2+]i plays a critical role in insulin granule exocytosis. Therefore we examined the effects of KB-R7943 and SN-6 on [Ca2+]i in INS-1E cells in the presence of a sulfonylurea. As shown in Fig. 3, in the presence of 10 μM glipizide, KB-R7943 but not SN-6, significantly increased [Ca2+]i in a concentration-dependent manner.
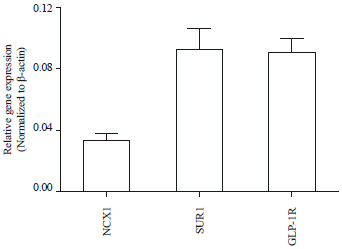 | |
| Fig. 1: | INS-1E cells express NCX1. Gene expression of NCX1 was measured using SYBR green qPCR according to the Method. The data shown is Mean±SEM from 5 independent experiments |
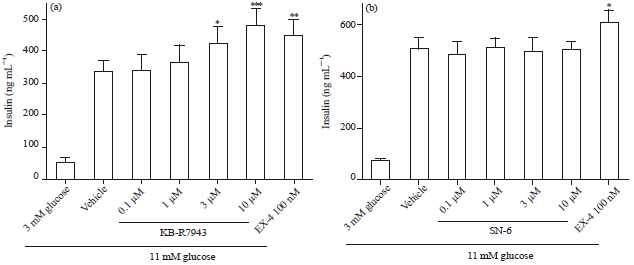 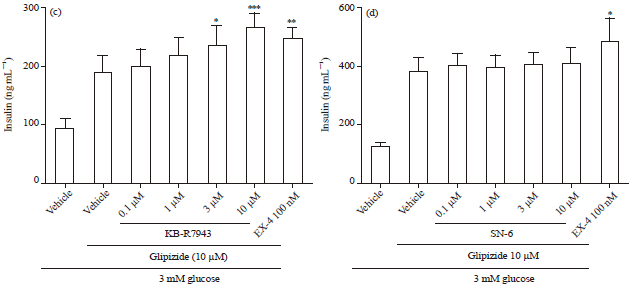 | |
| Fig. 2(a-d): | Different effects of KB-R7943 and SN-6 on glucose and glipizide-stimulated insulin secretion in INS-1E cells. INS-1E cells in 96-well plates were treated with agents for 1 h, supernatants were collected for insulin measurement according to the Method. Concentration response of (a) KB-R7943 (b) SN-6 on 11 mM glucose-stimulated insulin secretion in INS-1E cells. Concentration response of (c) KB-R7943 (d) SN-6 on 10 μM glipizide-stimulated insulin secretion in INS-1E cells. Data is Mean±SEM of 4-7 independent experiments where each experimental condition was ran in triplicates. * p<0.05, **p<0.01, ***p<0.001, compared to vehicle at 11 mM glucose or 10 μM glipizide by one-way ANOVA |
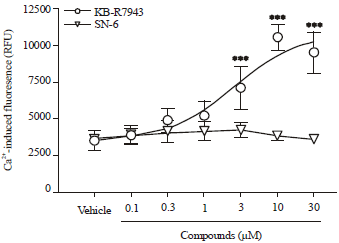 | |
| Fig. 3: | KB-R7943 but not SN-6 increased [Ca2+]i in the presence of glipizide in INS-1E cells. INS-1E cells were labeled with Calcium-4 dye and then challenged with KB-R7943 or SN-6 in the presence of glipizide (10 μM). Free intracellular calcium-induced fluoresence was measured according to the Method. Data is Mean±SEM of 6 independent experiments where each experimental condition was ran in triplicates. ***p<0.001 compared to vehicle control by one-way ANOVA |
No significant effect on [Ca2+]i was observed when INS-1E cells were treated with KB-R7943 alone.
Knockdown of NCX1 does not attenuate KB-R7943-mediated enhancement of glucose or glipizide-stimulated insulin secretion and glipizide-stimulated increase of [Ca2+]i: To examine the role of NCX1 in KB-R7943-mediated enhancement of glucose- or glipizide-stimulated insulin secretion and glipizide-stimulated increase of [Ca2+]i, we used the approach of knockdown of NCX1 by a specific siRNA. First, several pairs of pre-designed NCX1 siRNAs were examined and the siRNA with the highest knockdown on NCX1 gene expression was selected. Transfection with the NCX1 siRNA (20 nM) for 48 h reduced NCX1 gene expression by 70% measured by qPCR (Fig. 4a) and NCX1 protein expression by 62% measured by Western blots (Fig. 4b) in INS-1E cells.
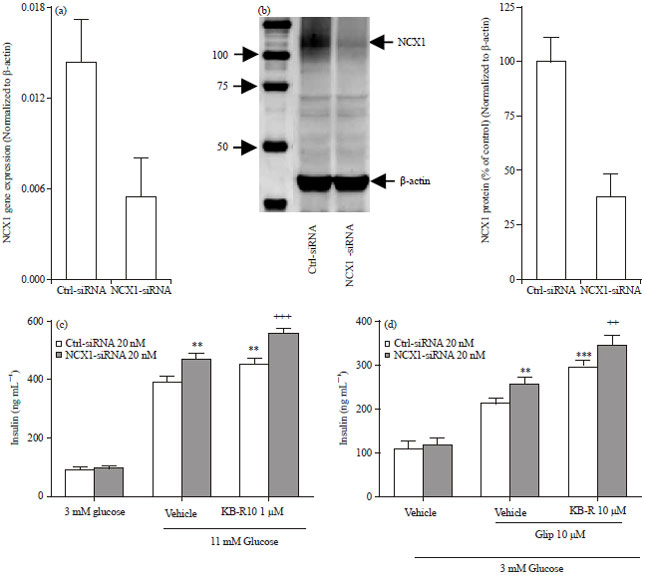 | |
| Fig. 4(a-d): | Effects of the NCX1 siRNA on NCX1 expression and KB-R7943 enhanced insulin secretion in INS-1E cells. (a) INS-1E cells transfected with scrambled control (Ctrl-siRNA) or NCX1 siRNA at 20 nM each for 48 h. NCX1 gene expression levels were measured according to Method and normalized against the gene expression levels of β-actin. Data shown are Mean±SEM from 3 independent experiments, (b) Left panel, INS-1E cells were transfected with scrambled control or NCX1 siRNAs for 48 h, NCX1 and β-actin protein levels were measured using western blot according to the Method. Bands shown on the left side are molecular weights (kDa). A representative Western blot is shown. Right panel, NCX1 protein intensities in Western blots were measured using quantity one and normalized against protein intensities of corresponding β-actin. Data is normalized by scrambled control-siRNA group which is set as 100%. Data is Mean±SEM from 5 independent experiments, (c) INS-1E cells were transfected with siRNAs, 48 h later, cells were treated with 3 or 11 mM glucose with or without KB-R7943 (KB-R10 μM) for 1 h. Insulin levels in supernatants were measured. Data is Mean±SEM from 3 independent experiments. **p<0.01 compared to Ctrl-siRNA group treated with vehicle. ++p<0.01 compared to NCX1-siRNA group treated with vehicle, (d) Transfected INS-1E cells were treated with or without KB-R7943 (KB-R10 μM) in the presence or absence of glipizide (10 μM). Data shown are Mean±SEM from 4 independent experiments. **p<0.01 and ***p<0.001, compared to Ctrl-siRNA group treated with vehicle. ++p<0.01 compared to NCX1-siRNA group treated with vehicle |
However, knockdown of NCX1 by the siRNA had no significant effect on KB-R7943-mediated enhancement of both glucose (11 mM) and glipizide-stimulated insulin secretion (Fig. 4c, d), although knockdown of NCX1 itself enhanced glucose- or glipizide-stimulated insulin secretion. Similarly, knockdown of NCX1 by the siRNA did not significantly change KB-R7943-mediated enhancement of [Ca2+]i in the presence of glipizide (Fig. 5).
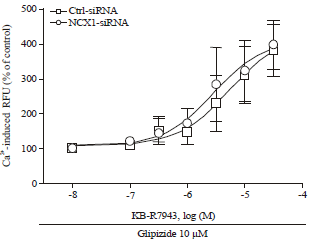 | |
| Fig. 5: | Effect of knockdown of NCX1 on KB-R7943 induced enhancement in [Ca2+]i. INS-1E cells transfected with scrambled control (Ctrl-siRNA) or the NCX1 siRNA. 48 h later, KB-R7943-induced enhancement of [Ca2+]i in the presence of 10 μM glipizide and 3 mM glucose was measured according to the Method. Data shown are Mean±SEM of percent of controls from 3 independent experiments where each condition was run in triplicates. Control groups were set as 100% |
DISCUSSION
Pancreatic β-cells express NCX1 at a relatively high level that may play a role in the regulation of insulin secretion. In this study, it was found that KB-R7943, a known NCX1 inhibitor, enhanced glucose- and sulfonylurea-stimulated insulin secretion and sulfonylurea-stimulated increase of [Ca2+]i in INS-1E cells. These results are consistent with the observations by Hamming et al. (2010) that KB-R7943 enhanced GSIS and sulfonylurea-stimulated increase of [Ca2+]i in mouse and human pancreatic islets. However, in Hamming et al. (2010) study the effects of other NCX1 inhibitors including SN-6 on GSIS or [Ca2+]i in pancreatic islets were not tested, therefore whether the results of KB-R7943 can be reproduced by SN-6 in pancreatic islets is unknown. In addition, the effects of knockdown of NCX1 on KB-R7943-induced enhancement on GSIS or sulfonylurea-stimulated increase of [Ca2+]i in pancreatic islets or islet cells were not examined in that study, whether the enhancement effects by KB-R7943 on GSIS or [Ca2+]i could be reduced after knockdown of NCX1 in pancreatic islet cells is also unknown. In current study, the effect of KB-R7943 on INS-1E cells could not be reproduced using another NCX1 inhibitor SN-6. Although, both KB-R7943 and SN-6 are benzyloxyphenyl derivatives and share similar structure, SN-6 is more potent for NCX1 vs. NCX2 and NCX3. KB-R7943 is less potent for NCX1 but more potent for NCX3 and NCX2 (Iwamoto, 2004; Amran et al., 2003). Nevertheless, the potencies of KB-R7943 and SN-6 for NCX1 are very similar (IC50: 4.9 μM vs. 3.2 μM, respectively). The ineffectiveness of SN-6 on glucose-and glipizide-stimulated insulin secretion and glipizide-stimulated increase of [Ca2+]i suggests that NCX1 is not the molecular target for the effect of KB-R7943 on GSIS in INS-1E cells. This argument is supported by the observation that knockdown of NCX1 did not attenuate KB-R7943-mediated enhancement on glucose-and glipizide-stimulated insulin secretion and glipizide-stimulated increase of [Ca2+]i. These results strongly suggested that the enhancement effect of KB-R7943 on insulin secretion and [Ca2+]i in INS-1E cells is mediated through an NCX1 independent mechanism.
Nguidjoe et al. (2011) and Herchuelz et al. (2013) previously reported that heterozygous inactivation of NCX1 (NCX1+/-) can lead to an increase of GSIS in islets of NCX1+/- mice compared to that in islets from wild type (NCX1+/+) mice. In agreement with that observation, current study also found that knockdown of NCX1 significantly enhanced glucose-and glipizide-stimulated insulin secretion compared to that in the control siRNA group in INS-1E cells (Fig. 4c, d). In NCX1+/- mouse model, Nguidjoe et al. (2011) found that insulin content in islet β-cells significantly increased and β-cell mass in islets significantly enlarged when compared to those from the wild type mice. Significant increase of insulin content in β-cells or enlarged β-cell mass in islets could have significant impacts on GSIS from islets. Therefore, the results from Nguidjoe et al. (2011) study could be driven by several mechanisms in addition to the inactivation of NCX1, most likely involving the higher insulin content and larger β-cell mass. Similarly, knockdown of NCX1 in INS-1E cells many have some uncovered effects on cellular function, although we did not investigate the potential mechanism of silencing NCX1-induced increase of GSIS. Interestingly, Nguidjoe et al. (2011) did not test the effect of NCX1 inhibitors including KB-R7943 on GSIS in islets from either NCX1+/- or NCX1+/+ mice, it is unclear whether KB-R7943 can enhance GSIS in islets from NCX1+/- mice. Therefore, it is entirely possible that the enhancement effects on GSIS in NCX1+/- islets or in INS-1E cells with NCX1 knockdown may be mediated through various mechanisms.
Because the effect of KB-R7943 on GSIS in INS-1E cells is unlikely mediated through NCX1, other membrane molecules probably are targets for the effect. Compared to SN-6, KB-R7943 is much less selective, also interacts with many other membrane proteins. It was reported that KB-R7943 can block canonical transient receptor potential channels (TRPC (Pezier et al., 2009). TRPC belongs to a subgroup of Transient Receptor Potential (TRP) channel superfamily. It is known that KB-R7943 but not SN-6, blocks TRPC3, 5 and 6 channels with IC50 around 0.4-2 μM range (Kraft, 2007). However, it is unknown whether these TRPC are expressed on pancreatic β-cells and what kind of role they may play in regulating insulin secretion in β-cells. Pancreatic β-cells such as RINm5F cells and rat pancreatic islets express Transient Receptor Potential Ankyrin 1 (TRPA1) channels, a member of subgroup of TRP channel superfamily (Numazawa et al., 2012). TRPC and TRPA1 channels are closely related, it is possible that KB-R7943 may also block TRPA1. However, activation of TRPA1 was reported to enhance GSIS (Numazawa et al., 2012), therefore it is unlikely that the blockade effect of KB-R7943 on TRP channels will contribute to its enhancement effects on insulin secretion.
KB-R7943 was also reported to be a potent inhibitor for hERG potassium channels with IC50 around 89-120 nM (Cheng et al., 2012). There was one report suggested that inhibition of hERG potassium channels could increase GSIS (Rosati et al., 2000), however, we believe that the inhibitory effect on hERG does not contribute to its enhancement on GSIS from INS-1E, because KB-R7943 is much less potent to enhance GSIS (EC50 is around 5.6 μM) in INS-1E cells compared to the potency to inhibit hERG (IC50 = 89-120 nM).
It was also reported that KB-R7943 directly inhibits type 1 ryanodine receptor with IC50 of 5.1 μM and type 2 ryanodine receptor with IC50 of 13.4 μM (Barrientos et al., 2009). In contrast, very little effect was detected for SN-6 on ryanodine receptors (Barrientos et al., 2009). This off-target effect of KB-R7943 is also unlikely to contribute to its effect on GSIS in INS-1E cells, because blockade of ryanodine receptors reduces GSIS in β-cells (Mitchell et al., 2003). Similarly, the effect on L-type calcium channels by KB-R7943 is also unlikely to play a role in its enhancement effect on GSIS in INS-1E cells, because inhibition of L-type calcium channels reduces GSIS (Liu et al., 2003; Mears, 2004). KB-R7943 but not SN-6 also inhibits mitochondrial Ca2+ uniporter (Santo Domingo et al., 2007). This effect of KB-R7943 should not contribute to its role in GSIS, because inhibition of mitochondrial Ca2+ uniporter reduces cytoplasmic ATP/ADP ratio, leading to decrease of insulin secretion (Tarasov et al., 2012). It was also reported that KB-R7943 inhibits mitochondrial complex I (Brustovetsky et al., 2011), however inhibition of mitochondrial complex I will reduce mitochondrial metabolism, leading to decrease of glucose-stimulated insulin secretion (Westerlund and Bergsten, 2001). Moreover, KB-R7943 also interacts with NMDA receptor and nicotinic receptor, two G-protein-coupled receptors mainly expressed in central nerve systems. There is very little information available up-to-date about their role in pancreatic β-cells, whether the effect of KB-R7943 on GSIS from INS-1E cells involves the antagonism property for NMDA and nicotinic receptors is a question which needs further investigation.
In addition to the inhibitory effects on many membrane proteins, KB-R7943 was also found to activate large conductance Ca2+-activated K+ (BK) channels in Human Umbilical Vein Endothelial Cells (HUVECs) and mouse aortic smooth muscle cells (Liang et al., 2008a, b). It has been shown that BK channels express on rodent and human pancreatic β-cells and are involved in regulating electrical activity for insulin secretion (Braun et al., 2008). However the role of BK channels in β-cells for insulin secretion is still controversial and different results obtained using different model systems. Braun et al. (2008) initially found that inhibition of BK channels in human β-cells enhanced GSIS. However, it was reported recently that GSIS from islets of BK knockout mice was significantly lower than that from islets of wild type mice (Dufer et al., 2011). Similarly, they also found that BK channel specific inhibitor iberiotoxin significantly reduced GSIS from pancreatic islets obtained from wild type mice. Therefore it is possible that activation of BK channels may contribute to the enhancement effect of KB-R7943 on GSIS in β-cells.
In addition to the effects of calcium channels and other membrane proteins on insulin secretion through regulating [Ca2+]i levels, the balance between intracellular free radicals and antioxidants also affects insulin secretion (Pourkhalili et al., 2009). Whether KB-R7943 can change the balance between free radicals and antioxidants is unknown, however potential effect of KB-R7943 on the balance may also contribute to its enhancement effect on GSIS.
CONCLUSION
In summary, it was discovered that NCX1 inhibitor KB-R7943 but not SN-6, enhances GSIS from INS-1E cells through an NCX1-independent pathway. Since KB-R7943 interacts with many cellular targets, further studies are necessary to address the detail molecular mechanism for KB-R7943 mediated GSIS in INS-1E cells.
ACKNOWLEDGEMENT
We thank Dr. Dongmin Liu for providing the INS-1E cell line and Dr Arvinder Dhalla for suggestions.
REFERENCES
- Amran, M.S., N. Homma and K. Hashimoto, 2003. Pharmacology of KB‐R7943: A Na+-Ca2+ exchange inhibitor. Cardiovas. Drug Rev., 21: 255-276.
CrossRefDirect Link - Annunziato, L., G. Pignataro and G.F. Di Renzo, 2004. Pharmacology of brain Na+/Ca2+ exchanger: From molecular biology to therapeutic perspectives. Pharmacol. Rev., 56: 633-654.
CrossRefDirect Link - Barrientos, G., D.D. Bose, W. Feng, I. Padilla and I.N. Pessah, 2009. The Na+/Ca2+ exchange inhibitor 2-(2-(4-(4-nitrobenzyloxy) phenyl) ethyl) isothiourea methanesulfonate (KB-R7943) also blocks ryanodine receptors type 1 (RyR1) and type 2 (RyR2) channels. Mol. Pharmacol., 76: 560-568.
CrossRefPubMedDirect Link - Bers, D.M., J.W.M. Bassani and R.A. Bassani, 1996. Na-Ca exchange and Ca fluxes during contraction and relaxation in mammalian ventricular muscle. Ann. N. Y. Acad. Sci., 779: 430-442.
CrossRefDirect Link - Braun, M., R. Ramracheya, M. Bengtsson, Q. Zhang and J. Karanauskaite et al., 2008. Voltage-gated ion channels in human pancreatic β-cells: Electrophysiological characterization and role in insulin secretion. Diabetes, 57: 1618-1628.
CrossRefDirect Link - Brustovetsky, T., M.K. Brittain, P.L. Sheets, T.R. Cummins, V. Pinelis and N. Brustovetsky, 2011. KB‐R7943, an inhibitor of the reverse Na+/Ca2+ exchanger, blocks N-methyl-D-aspartate receptor and inhibits mitochondrial complex I. Br. J. Pharmacol., 162: 255-270.
CrossRefDirect Link - Cheng, H., Y. Zhang, C. Du, C.E. Dempsey and J.C. Hancox, 2012. High potency inhibition of hERG potassium channels by the sodium-calcium exchange inhibitor KB‐R7943. Br. J. Pharmacol., 165: 2260-2273.
CrossRefDirect Link - Dufer, M., Y. Neye, K. Horth, P. Krippeit-Drews and A. Hennige et al., 2011. BK channels affect glucose homeostasis and cell viability of murine pancreatic beta cells. Diabetologia, 54: 423-432.
CrossRefDirect Link - Elbashir, S.M., J. Harborth, W. Lendeckel, A. Yalcin, K. Weber and T. Tuschl, 2001. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature, 411: 494-498.
CrossRefDirect Link - Hamming, K.S.C., D. Soliman, N.J. Webster, G.J. Searle and L.C. Matemisz et al., 2010. Inhibition of β-cell sodium-calcium exchange enhances glucose-dependent elevations in cytoplasmic calcium and insulin secretion. Diabetes, 59: 1686-1693.
CrossRefDirect Link - Herchuelz, A., O. Diaz-Horta and F. van Eylen, 2002. Na/Ca exchange in function, growth and demise of β-cells. Ann. N. Y. Acad. Sci., 976: 315-324.
CrossRefDirect Link - Herchuelz, A., A. Kamagate, H. Ximenes and F. van Eylen, 2007. Role of Na/Ca exchange and the plasma membrane Ca2+-ATPase in β cell function and death. Ann. N. Y. Acad. Sci., 1099: 456-467.
CrossRefDirect Link - Herchuelz, A., E. Nguidjoe, L. Jiang and N. Pachera, 2013. Na+/Ca2+ exchange and the plasma membrane Ca2+-ATPase in β-cell function and diabetes. Adv. Exp. Med. Biol., 961: 385-394.
CrossRefDirect Link - Iwamoto, T., 2004. Forefront of Na+/Ca2+ exchanger studies: Molecular pharmacology of Na+/Ca2+ exchange inhibitors. J. Pharmacol. Sci., 96: 27-32.
CrossRefDirect Link - Iwamoto, T., Y. Watanabe, S. Kita and M.P. Blaustein, 2007. Na+/Ca2+exchange inhibitors: A new class of calcium regulators. Cardiovas. Haematol. Disorders-Drug Targets, 7: 188-198.
CrossRefDirect Link - Kraft, R., 2007. The Na+/Ca 2+ exchange inhibitor KB-R7943 potently blocks TRPC channels. Biochem. Biophys. Res. Commun., 361: 230-236.
CrossRefDirect Link - Liang, G.H., J.A. Kim, G.H. Seol, S. Choi and S.H. Suh, 2008. The Na+/Ca2+ exchanger inhibitor KB-R7943 activates large-conductance Ca2+-activated K + channels in endothelial and vascular smooth muscle cells. Eur. J. Pharmacol., 582: 35-41.
CrossRefDirect Link - Liang, G.H., M.Y. Kim, S. Park, J.A. Kim, S. Choi and S.H. Suh, 2008. Intracellular Na+ modulates large conductance Ca2+-activated K+ currents in human umbilical vein endothelial cells. Pflugers Arch. Eur. J. Physiol., 457: 67-75.
CrossRefDirect Link - Liu, G., N. Dilmac, N. Hilliard and G.H. Hockerman, 2003. Ca v 1.3 is preferentially coupled to glucose-stimulated insulin secretion in the pancreatic β-cell line INS-1. J. Pharmacol. Exp. Ther., 305: 271-278.
PubMed - MacDonald, P.E., J.W. Joseph and P. Rorsman, 2005. Glucose-sensing mechanisms in pancreatic β-cells. Philos. Trans. Royal Soc. B: Biol. Sci., 360: 2211-2225.
CrossRefDirect Link - Mears, D., 2004. Regulation of insulin secretion in islets of Langerhans by Ca2+ channels. J. Membr. Biol., 200: 57-66.
CrossRefDirect Link - Mitchell, K.J., F.A. Lai and G.A. Rutter, 2003. Ryanodine receptor type I and nicotinic acid adenine dinucleotide phosphate receptors mediate Ca2+ release from insulin-containing vesicles in living pancreatic beta-cells (MIN6). J. Biol. Chem., 278: 11057-11064.
CrossRefPubMedDirect Link - Nguidjoe, E., S. Sokolow, S. Bigabwa, N. Pachera and E. D'Amico et al., 2011. Heterozygous inactivation of the Na/Ca exchanger increases glucose-induced insulin release, β-cell proliferation and mass. Diabetes, 60: 2076-2085.
CrossRefDirect Link - Numazawa, S., M. Takase, T. Ahiko, M. Ishii, S.I. Shimizu and T. Yoshida, 2012. Possible involvement of transient receptor potential channels in electrophile-induced insulin secretion from RINm5F cells. Biol. Pharm. Bull., 35: 346-354.
CrossRefDirect Link - Ouardouz, M., G.W. Zamponi, W. Barr, L. Kiedrowski and P.K. Stys, 2005. Protection of ischemic rat spinal cord white matter: Dual action of KB-R7943 on Na+/Ca 2+ exchange and L-type Ca 2+ channels. Neuropharmacology, 48: 566-575.
CrossRefDirect Link - Pezier, A., Y.V. Bobkov and B.W. Ache, 2009. The Na+/Ca2+ exchanger inhibitor, KB-R7943, blocks a nonselective cation channel implicated in chemosensory transduction. J. Neurophysiol., 101: 1151-1159.
CrossRefDirect Link - Pintado, A.J., C.J. Herrero, A.G. Garcia and C. Montiel, 2000. The novel Na+/Ca2+ exchange inhibitor KB‐R7943 also blocks native and expressed neuronal nicotinic receptors. Br. J. Pharmacol., 130: 1893-1902.
CrossRefDirect Link - Pourkhalili, N., S. Pournourmohammadi, F. Rahimi, S. Vosough-Ghanbari, M. Baeeri, S.N. Ostad and M. Abdollahi, 2009. Comparative effects of calcium channel blockers, autonomic nervous system blockers and free radical scavengers on diazinon-induced hyposecretion of insulin from isolated islets of Langerhans in rats. Arh. Hig. Rada. Toksikol., 60: 157-164.
PubMed - Rosati, B., P. Marchetti, O. Crociani, M. Lecchi and R. Lupi et al., 2000. Glucose-and arginine-induced insulin secretion by human pancreatic β-cells: The role of HERG K+ channels in firing and release. FASEB J., 14: 2601-2610.
PubMed - Santo‐Domingo, J., L. Vay, E. Hernandez‐SanMiguel, C.D. Lobaton, A. Moreno, M. Montero and J. Alvarez, 2007. The plasma membrane Na+/Ca2+ exchange inhibitor KB‐R7943 is also a potent inhibitor of the mitochondrial Ca2+ uniporter. Br. J. Pharmacol., 151: 647-654.
CrossRefDirect Link - Straub, S.G. and G.W.G. Sharp, 2002. Glucose-stimulated signaling pathways in biphasic insulin secretion. Diabetes/Metab. Res. Rev., 18: 451-463.
CrossRefDirect Link - Tarasov, A.I., F. Semplici, M.A. Ravier, E.A. Bellomo and T.J. Pullen et al., 2012. The mitochondrial Ca2+ uniporter MCU is essential for glucose-induced ATP increases in pancreatic β-cells. PloS One, Vol. 7.
CrossRefDirect Link - Terracciano, C.M. and J.C. Hancox, 2013. ORM‐10103: A significant advance in sodium‐calcium exchanger pharmacology? Br. J. Pharmacol., 170: 765-767.
CrossRefDirect Link - Van Eylen, F., C. Lebeau, J. Albuquerque-Silva and A. Herchuelz, 1998. Contribution of Na/Ca exchange to Ca2+ outflow and entry in the rat pancreatic β-cell: Studies with antisense oligonucleotides. Diabetes, 47: 1873-1880.
CrossRefDirect Link - Westerlund, J. and P. Bergsten, 2001. Glucose metabolism and pulsatile insulin release from isolated islets. Diabetes, 50: 1785-1790.
CrossRefDirect Link