
Kuldeep Dhama
Division of Pathology, Indian Veterinary Research Institute, Izatnagar (IVRI), Bareilly, Uttar Pradesh, 243122, India
Mani Saminathan
Division of Pathology, Indian Veterinary Research Institute, Izatnagar (IVRI), Bareilly, Uttar Pradesh, 243122, India
Siju Susan Jacob
Division of Parasitology, Indian Veterinary Research Institute, Izatnagar (IVRI), Bareilly, Uttar Pradesh, 243122, India
Mithilesh Singh
Immunology Section, Indian Veterinary Research Institute, Izatnagar (IVRI), Bareilly, Uttar Pradesh, 243122, India
K. Karthik
Division of Bacteriology and Mycology, Indian Veterinary Research Institute, Izatnagar (IVRI), Bareilly, Uttar Pradesh, 243122, India
Amarpal .
Divsion of Surgery, Indian Veterinary Research Institute, Izatnagar (IVRI), Bareilly, Uttar Pradesh, 243122, India
Ruchi Tiwari
Department of Veterinary Microbiology and Immunology, College of Veterinary Sciences and Animal Husbandry, Uttar Pradesh Pandit Deen Dayal Upadhayay Pashu Chikitsa Vigyan Vishvidhyalaya Ewam Go-Anusandhan Sansthan (DUVASU), Mathura, Uttar Pradesh, 281001, India
Lakshmi Tulasi Sunkara
Department of Animal Science, Oklahoma State University, Stillwater, OK, 74078, USA
Yashpal Singh Malik
Division of Biological Standardization, Indian Veterinary Research Institute, Izatnagar, Bareilly, Uttar
Pradesh, 243122, India
Raj Kumar Singh
Indian Veterinary Research Institute, Izatnagar, Bareilly Uttar Pardesh, 243122, India
International Journal of Pharmacology
Year: 2015 | Volume: 11 | Issue: 4 | Page No.: 253-290







ABSTRACT
In veterinary and medical sciences, immunomodulation is an area wherein extensive studies have been conducted to devise methods to improve disease resistance as well as to prevent or control immune disorders of host by optimum regulation of the immune system. Today, most infectious diseases of man and animals are treated and controlled mainly by using broad-spectrum antibiotics and vaccines. However, the antibacterial agents are becoming increasingly ineffective due to rapid emergence of resistant microbial strains. So, there is high requirement for novel and improved alternative therapeutic and prophylactic strategies to manage several diseases which are flaring at alarming pace because of the increase in international traffic, globalization and changing food habbits. Immunomodulation is focused on manipulation of immune system to control the infections and other adverse health effects with precise regulation to avoid any complications while suppressive or potentiating efforts are made to benefit the animal and human health. The main aim of this review is to give a closer insight into the potential immunomodulatory molecules, synthetic and natural, that are capable of modifying the immune responses including conventional and novel immunomodulators like adjuvants, cytokines, hormones, glucocorticoids, host defense peptides, microbial products, toll like receptors, synthetic compounds, probiotics, nutrients, vitamins, minerals, herbs, panchgavya, polysaccharides, helminths, vaccines and others. These immunomodulatory regimens could successfully offer the health industries with the most natural methods for enhancement of disease resistance, boosting vaccination immunity and prevention of various infections, disorders, cancer and stress related diseases. The updated information will be highly useful for scientists, veterinary/medical professionals, pharmaceutical industries, livestock and poultry industry to create a healthier future for people and their companion animals.
PDF Abstract XML References Citation
Received: January 25, 2015;
Accepted: April 03, 2015;
Published: April 18, 2015
How to cite this article
Kuldeep Dhama, Mani Saminathan, Siju Susan Jacob, Mithilesh Singh, K. Karthik, Amarpal ., Ruchi Tiwari, Lakshmi Tulasi Sunkara, Yashpal Singh Malik and Raj Kumar Singh, 2015. Effect of Immunomodulation and Immunomodulatory Agents on Health with some Bioactive Principles, Modes of Action and Potent Biomedical Applications. International Journal of Pharmacology, 11: 253-290.
DOI: 10.3923/ijp.2015.253.290
URL: https://scialert.net/abstract/?doi=ijp.2015.253.290
DOI: 10.3923/ijp.2015.253.290
URL: https://scialert.net/abstract/?doi=ijp.2015.253.290
INTRODUCTION
At present chemotherapy is the most common and widely acceptable approach to control the microbial infections of veterinary and medical importance. According to the World Health Organization the antibiotics will not be effective beyond 2020 because of the rapid emergence of drug resistant strains (Hancock and Sahl, 2006; Hamill et al., 2008; Dhama et al., 2013a). Species evolution theory is so true that all the microorganisms try to evade antibiotics which are intended to kill them. This makes things worse for the animal and human community since only a handful of antibiotics are left with to protect from the dreadful pathogens (Karthik et al., 2012; Tiwari et al., 2013). Sooner these few antibiotics may also not act against those pathogens against which they are targeted. Antibiotic resistance is the hot topic of this decade and in order to survive there is need to search alternatives to counteract the pathogens and also to safeguard human and animal health. Novel treatments include phage therapy, cytokine therapy, herbal medicines, essential oils extracts and cowpathy (Dhama et al., 2013a-c, 2014; Karthik et al., 2014; Gopi et al., 2014; Verma et al., 2014). In that instance immunomodulation with an aim to increase the immune potential of the body remains the sole alternative to control the infection. During the course of time, so many immunomodulators are introduced and it was found that herbal immunomodulators based on ancient Indian ayurvedic system of medicine were most effective without any side effects (Tan and Vanitha, 2004; Spelman et al., 2006; Borsuk et al., 2011; Hashemi and Davoodi, 2012; Mahima et al., 2012a). In animals, immunomodulation is defined as the ability to regulate an immune response to enhance the production efficiency in food-producing animals and the substances that put forth this control are called immunomodulators (Chauhan, 2010; Fraile et al., 2012; Zhao et al., 2012). Immunomodulation refers to the manipulation of immune system and all therapeutic interruptions targeted for modulating the immune system (Mahima et al., 2013a). The augmentation of immune response, known as immunostimulation or immunopotentiation, can be utilized to prevent infection by vaccination through activation of humoral immunity, to fight against an already established infection by shifting the immune response to cell mediated type and to fight against cancer by the use of cytokines, tumour specific antibodies and tumour infiltrating lymphocytes. The suppression of immune responsiveness, known as immune suppression, is usually practiced in allergy, autoimmunity and organ transplantation. Immunomodulation can be either specific or non-specific. Specific immunomodulation is limited to a single antigen such as vaccination. Non-specific immunomodulation implies for a more generalized change in immune responsiveness both in innate and adaptive immunity, leading to altered host reactivity to many different antigens.
An immunomodulator may be defined as any biological or synthetic substance that can stimulate/suppress either innate or adaptive or both arms of the immune system (Agarwal and Singh, 1999). A potent immunomodulator have got many advantages over antimicrobials. Since immunomodulators do not affect directly microbes, they may avoid the problem of rapid emergence of resistance. Traditional antimicrobials in immuno compromised patients often work poorly and the importance of immunomodulators then can be realized. The antimicrobials are specific in nature whereas immunomodulators provide broad spectrum capability against bacterial, viral and fungal diseases and thereby provide non-specific emergency-therapeutic approaches in the event of emergence of a strange pathogen or biowarfare agents (Gallois and Oswald, 2008). In the era of one world one health one medicine concept, the potent applications of useful immunomodulators needs to be encouraged (Dhama et al., 2013d). Recently, immunomodulators like helminths and mesenchymal stem cells have been proposed to be promising therapeutics for autoimmune disorders including systemic sclerosis, systemic lupus erythematosus, Crohn’s disease and allergic diseases (Reddy, 2010; Sanchez-Berna et al., 2014).
This review mainly aims at providing a close insight into the various kinds of potential immunomodulatory substances and regimens covering both synthetic and natural as well as conventional and novel immunomodulators including adjuvants, cytokines, thymic products, hormones, glucocorticoids, host defense peptides, microbial products, probiotics, synthetic chemical compounds, toll like receptors, nutrients, vitamins, minerals, herbs, panchgavya elements, polysaccharides, helminths, vaccines and others. The updated information will be highly useful for researchers, scientists, scholars, pharmacists, veterinary/medical professionals, pharmaceutical industries, animal producers/owners and livestock and poultry industry to create a healthier future for people and their companion animals.
HISTORY
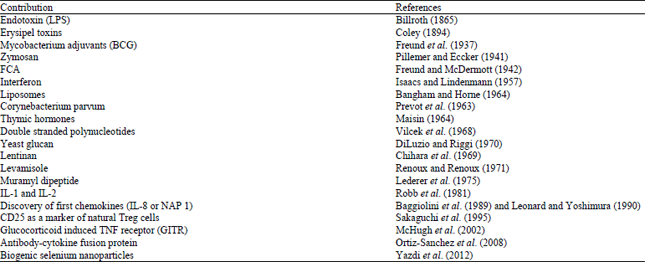
The concept of immunomodulation was first emerged in 1796. It was introduced by Edward Jenner, who carried out the first "vaccination" against small pox. Thereafter, several efforts have been made to modulate the immune system to encounter the external as well as internal attacks by pathogens. A brief history of immunomodulators is summarized in Table 1.
Objectives of immunomodulation: The primary objective of immunomodulation is to improve host resistance to external as well as internal attacks by the microbes or other infectious agents. The basic objectives of immunomodulation in domestic animals include:
| • | To activate powerful and prolonged immune response against disease causing microorganism |
| • | To speed up maturation of non specific and specific immunity during neonatal period and in young susceptible animals |
| • | To enhance local protective immune reactions at vulnerable sites such as mammary gland in dairy cattle or gastro intestinal tract in neonatal ruminants |
| • | To surmount the immunosuppressive effects of stress and environmental pollution |
| • | To enhance level and duration of immune response following vaccination |
| • | To maintain immune surveillance |
| Table 1: | History of Immunomodulators |
 | |
Characteristics of an ideal immunomodulator:
| • | It must have short withdrawal period and low tissue residues |
| • | It should stimulate both specific and non specific immune response |
| • | It should exert an adjuvant effect when administered along with a vaccine |
| • | It should be active by oral route after it enters into host system and should be stable both in its native state and upon incorporation into food and water |
| • | It should be compatible with wide range of drugs including antibiotics and anthelmintics |
| • | It should amplify primary and secondary immune response to infectious agents |
| • | It should have a defined chemical composition and biological activity |
| • | It should be inexpensive |
| • | It should be nontoxic even at high dose rates for animals and humans |
| • | It should neither be antigenic nor pyrogenic |
| • | It should not be teratogenic, carcinogenic or have long-term side effects in animals |
| • | It should not be excreted in milk or eggs |
Purpose of immunomodulation: The modification of immune response in animals may be intended to the following:
| • | Immuno-stimulation of normal animals |
| • | Immunosuppression of normal animals |
| • | Immuno-restoration of immune suppressed animals |
CLASSIFICATION AND APPLICATIONS OF IMMUNOMODULATORS
Physiological products
Neuroendocrine hormones: There exists a close interaction between the immune and nervous systems which has been extensively studied and documented (Ohira et al., 2013). The immuno competent cells not only express receptors to neuroendocrine mediators but also secrete many of them. The interaction between neuroendocrine and immune system is very essential to maintain the homeostasis of the body (Blalock, 1989; Dan and Lall, 1998). Any alteration in this interaction can lead to enhanced susceptibility of the body to various infectious, inflammatory or autoimmune diseases (Eskandari et al., 2003). High vagal tone may regulate immune and physiological activity of brain (Ohira et al., 2013). There are two main mechanisms by which the Central Nervous System (CNS) controls the immune system. The first mechanism is Hypothalamic-Pituitary-Adrenal (HPA) axis mediated hormonal response, the second is via the release of nor-epinephrine (nor-adrenaline) from sympathetic and acetylcholine from parasympathetic nerves supplied to most of the immune organs (bone marrow, thymus, lymph nodes, spleen, etc) (Shepherd et al., 2005). Furthermore, neuropeptides such as substance P, somatostatin etc., are released into tissues from the peripheral sensory nerves by the antidromic activation mechanisms, where these molecules also modulate lymphoid cell activities. In the regulation of immune response, the role of hypothalamic hormones such as Thyrotropin-releasing hormone (TRH), Corticotropin-releasing hormone (CRH) and Gonadotropin-releasing hormone (GnRH) and major stress hormones viz., cortisol and dehydroepiandrosterone (DHEA) and growth hormone are important.
| Table 2: | Neuroendocrine hormones having Immunomodulatory properties |
 | |
| *VIP: Vasoactive intestinal peptide, **NGF: Nerve growth factor, ↑: Increase and ↓: Decrease | |
All these hormones and specific neurotransmitters and neuropeptides are recognized by specific receptors of lymphoid cells resulting in production of intracellular signals which subsequently influence immune cell function (Quintanar and Guzman-Soto, 2013). Ghrelin, an orexigenic stomach hormone blocks the lipopolysaccharide (LPS)-induced secretion of pro-inflammatory cytokines (IL-6) from immune cells (Beynon et al., 2013). Conversely, the immune system can also control the CNS through cytokines and change the individual’s behavior (Prieto-Moreno and Rosenstein, 2006; Uchakin et al., 2007). For both the immune and the neuroendocrine systems, cytokines are important endogenous modulators which along with their receptors are expressed in the neuroendocrine system thereby exerting effects both centrally as well as peripherally (Benveniste, 1998). To date, around 30 neuroendocrine mediators have been reported to be active upon the immune cells. The important neuroendocrine hormones, neurotransmitters and neuropeptides having immunomodulatory properties are summarized in Table 2.
Thymic products: Thymus plays a pivitol role in the formation of the lymphoid structures in the prenatal and early postnatal life and orchestrating the lymphoid system throughout the life. More than 20 thymic products have been discovered, of which thymosin-α 1, thymopoietin, thymulin, thymosin fraction 5, Thymic Humoral Factor (THF), THF-gamma 2, splenopentin and thymopentin are particularly important in regulation of immune system (Huang et al., 1981; Zatz et al., 1984; Singh et al., 1998). Among these major thymic hormones, thymopoietin (TP), was mainly isolated from the bovine thymus. It consists of 49 amino acids and has a molecular weight of 5 kDa. This protein mainly acts on neuromuscular system and has a role in T cell differentiation and its physiological function.
| Table 3: | Thymic products and their Immunomodulatory activities |
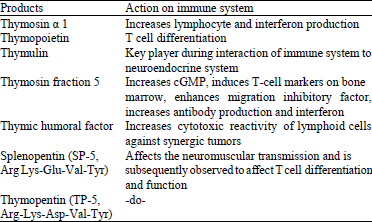 | |
It has two closely related polypeptides known as thymostimulin (TP-1) and TP-II. The thymopoietin shows cross reactivity with a protein found in lymph node and spleen known as splenin which differs from thymopoietin by aspartic acid at position 34. Interestingly, all these thymic products showed enhancement of immune efficiency in immunodeficient patients, cancer cases, bacterial and viral infections and can potentially be used for immunotherapy (Singh et al., 1998; Ben-Efraim et al., 1999). The immunomodulatory activity of each of them is summarized in Table 3.
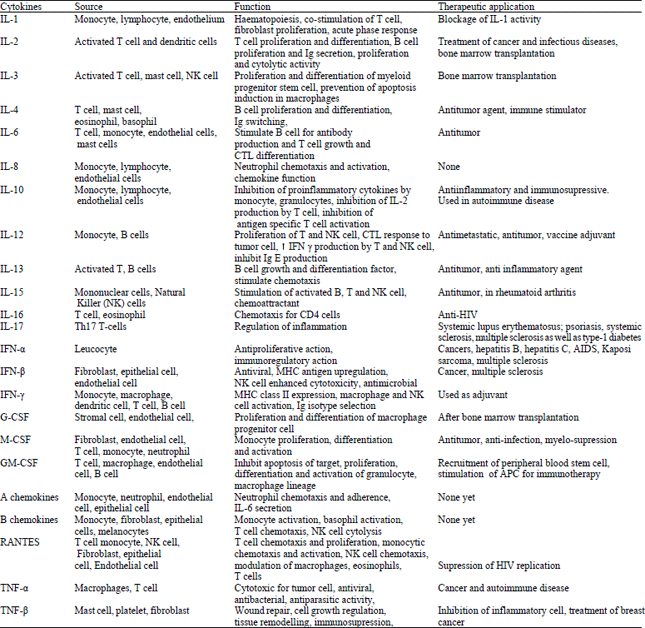
Cytokines: Cytokines are hormone like low molecular weight glycoproteins which are essential for proper functioning of the immune system (Dhama et al., 2008b, 2013c). A variety of immune cells produce them to communicate and orchestrate immune attacks (Pollard and Earnshaw, 2004). There may be initiation and perpetuation of autoimmune and infectious diseases along with tumour growth due to imbalanced network or abnormal production of cytokines (Adorini, 2003). The role of cytokines as immunomodulators is considered important for cure in future. They are autocrine, paracrine or endocrine in action and may exert either synergistic or antagonistic effect on their own production. Cytokines interestingly represent an improved and alternative treatment strategy against emerging pathogens. Cytokines can be used to enhance immunity and treat diseased or immune compromised individuals and to augment or induce desired immune responses against vaccines (Schijns and Horzinek, 1997). Cytokine secretion, when modulated, may offer novel approaches to treat a wide variety of diseases (Spelman et al., 2006). Modulation or intervention of immune response can essentially be affected by administering anti-inflammatory cytokines viz., interferon (IFN)-β; growth factors (transforming growth factor β, interleukin (IL)-4 and IL-10); or by neutralizing pro-inflammatory cytokines (IL-2, IFN-γ, IL-12, tumor necrosis factor (TNF)-α, IL-1β and IL-17) (Adorini, 2003). Daptomycin antibiotics have immunomodulatory properties and suppress the expression of inflammatory cytokines after host immune response stimulation by Methicillin Resistant Staphylococcus aureus (Tirilomis, 2014). Honey and its different components have the capacity to stimulate or inhibit the secretion of cytokines viz., TNF-α, IL-1β, IL-6 from monocytes and macrophages. It can also regulate the generation of reactive oxygen species from neutrophils (Majtan, 2014).
Cytokines as additional immunomodulators against infectious diseases: The concept that enhancement of the antimicrobial action of host immune mechanisms and/or antimicrobial agents is done by replacement or enhancement of natural mediators of host resistance, forms the basis of the rationale for using cytokines as adjunctive immunomodulators for infectious diseases. It is difficult to harness clinically the potential antimicrobial power of these agents despite the logical basis for this concept. A few examples are available, currently regarding the use of adjunctive immunomodulatory cytokines against infectious diseases. Examples are pegylated interferons and ribavirin for hepatitis C virus (HCV), use of recombinant nucleoside analogs and α-interferons for hepatitis B virus (HBV) (Forton and Karayiannis, 2006). Induction of Th1 immunity increases the efficacy of interferons against HCV (Trapero-Marugan et al., 2006). Immunotherapy with adjuvants in combination of recombinant cytokines is helpful in case of candidiasis (Van de Veerdonk et al., 2012). Depression is a common adverse effect of interferon-based therapies limiting their use in certain cases (Asnis and de la Garza, 2006). As an adjunctive agent for HIV-associated cryptococcal meningitis, adjunctive interferons hold promise in light of the confirmed benefits of interferon treatment for the prevention and cure of patients with chronic granulomatous disease associated bacterial infection (Gallin et al., 1991). Adjunctive cytokines like interferon-gamma moreover, can be utilized against invasive fungal infections as it can exhibit a protective response against fungi through Th1 helper T-cell responses (Armstrong-James et al., 2010; Jarvis et al., 2012; Delsing et al., 2014). IL-10 is effective against Leptospira infection (Rizvi et al., 2012). The ability to alleviate neutropenia forms the basis of the rationale for using colony stimulating factors (CSFs) derived from macrophages (GM-CSF) or granulocytes (G-CSF) (Antachopoulos and Roilides, 2005).
Cytokines as novel immunomodulators for vaccines: Right formulation of an appropriate vaccine adjuvant or immunomodulator is required in order to achieve the protective immunity in host following immunization. This can be achieved by the correct selection of vaccine adjuvants like cytokines. The vaccine adjuvants cytokines can be administered by two basic methods as genes encoded by DNA vaccines or as soluble proteins. In cattle, sheep, pigs and poultry soluble cytokines have been successfully used to modulate the immune system. Since most of the cytokines have a very short half-life and so large doses are needed in vivo to alter the immune responses to the co-administered antigen which often leads to development of adverse effects. This problem can be avoided by use of slow and sustained release preparations that release the cytokine for a prolonged period (Nicholls et al., 2010).
In order to ensure an appropriate immune response, new vaccines and their development necessitate production of new generation adjuvants (Nicholls et al., 2010); e.g., use of cytokines viz., mucosal vaccine adjuvants such as IL-7 and IL-12 (Stevceva et al., 2006). Cytokines and chemokines viz., monocyte chemotactic proteins (MCPs), granulocyte-macrophage colony-stimulating factor (GM-CSF) and macrophage inflammatory proteins (MIPs) have been incorporated into vaccines most recently. This can be done in lieu of their (cytokines’) potential to enhance recruitment of dendritic cells and monocytes from blood into the interstitial sites of vaccine delivery (Klavinskis et al., 2010). Killer cells, induced by cytokines like IL-2 and 15, can be used as vaccine against cancer (Lee and Margolin, 2011; Thanendrarajan et al., 2011). The important cytokines with their source, function along with their therapeutic application are listed in Table 4.
In general, classification of cytokines has been arbitrary and it depends upon the individuals. But most often, cytokines have been classified on the basis of similarity in their functional properties in controlling immunity and inflammation i.e., colony stimulating factors, interferons and pro-inflammatory and anti-inflammatory cytokines (Kogut, 2000; Reefman et al., 2010) (Table 5).
Importance of cytokines in veterinary practice: In the past, the usage of cytokines was restricted due to the problem of production of the protein in bulk. But recent developments in recombinant DNA technologies have solved this problem (Tossing, 2001). Different cytokines like interferons, interleukins etc., have been used successfully for treating various diseases. Minagawa et al. (1999) reported successful treatment of canine parvovirus in Europe by using alpha type 1 interferon of feline origin.
| Table 4: | Cytokines having Immunomodulatory activities |
 | |
| Table 5: | List of important immunomodulatory cytokines |
 | |
In the poultry industry interferons and other cytokines have been significantly used to reduce the use of vaccines and antibiotics in-feed (Lowenthal et al., 1998, 1999; Bedford, 2000). IFN-γ of poultry origin appears to have a significant role in the treatment of circovirus infection in young grey parrots (Stanford, 2004).
In Staphylococcus aureus mastitis, recombinant bovine interleukin-1 beta and interleukin-2 are found to be effective immunomodulators to augment natural resistant mechanisms similar to the normal response to pathogens. These may prove to be suitable alternatives to antibiotics or may be used along with them as effective therapeutic agents for mastitis (Daley et al., 1991). It is found that IL-11 is useful in treating Pseudomonas aeruginosa induced sepsis in immunocompromised animals (Opal et al., 1998). IL-2, IFN-gamma and TNF-alpha are recombinant bovine cytokines and all of them can be used as adjuvants in normal mammary gland to mobilize innate and acquired immunity (Alluwaimi, 2004). Immunostimulant cytokines have therapeutic potential against recurrent canine pyoderma which is a major problem for veterinarians. The helper T cell (Th1) cytokines (TNF α, IFN-γ and IL-1β) have profound effect on cell-mediated responses against bacterial infections (DeBoer et al., 2009).
For invasive fungal infections, adjunctive cytokines can be administered along with antifungal agents resulting in T-cell immune responses mainly of Th1 type in protection against fungi in experimental models. This has given rise to the rationale for using Colony Stimulating Factors (CSFs) which was derived from granulocytes (G-CSF) and macrophages (GM-CSF); partly based on their property to mitigate neutropenia (Antachopoulos and Roilides, 2005). It is assumed that adjunctive interferon holds promise as an adjunctive agent for HIV associated cryptococcal meningitis in the light of the established benefits of interferon therapy in preventing further bacterial complications in chronic granulomatous diseases (Datta and Pirofski, 2006; Pirofski and Casadevall, 2006). The recombinant cytokines such as GM-CSF, TNF, IL-1 and IFN-γ are used as adjunctive immunotherapy for the cure of disseminated candidiasis. These cytokines cause increased expression of dectin-1 receptors on phagocytic cell macrophages. It also enhances the activity of chitotriosidase which causes degradation of the chitin from the internal cell wall of C. albicans (Van de Veerdonk et al., 2012). IL-17 has got a major role to play for treating diseases like systemic lupus erythematosus, rheumatoid arthritis, psoriasis, multiple sclerosis, systemic sclerosis and type-1 diabetes (Kunz and Ibrahim, 2009). The extensive drug-resistant tuberculosis (XDR-TB) emergence has rendered chemotherapy futile due to which cytokine therapy may become one of the possible useful alternative. The efficacy of therapeutic strategies employing a single cytokine has not been reported adequately; thus basic research would be necessary to attain a better understanding of the mechanisms and interactions of cyokines involved in successful promotion of immune response in order to generate efficient cytokine-based XDR TB therapies. New research projects concerning cytokines based treatment for XDR TB should be appreciated and liberally funded. Chemokines may increase both mycobacterial killing activity of the effector cells as well as defense against intracellular multiplication of the bacteria and thus their role must also be explored (Rivero-Lezcano, 2008). Tumor necrosis factor-α and interleukin-6 have enormous roles to play in conditions like obesity (German et al., 2010; Zoran, 2010).
Glucocorticoids: Most frequently glucocorticoids are used for treating chronic inflammatory diseases that involve lymphocytes but in diseases mediated by macrophages, they are effective to a lesser extent (Kugelberg, 2014). The effect of glucocorticoids on immune system is biphasic as there is an initial inhibition during initial release of corticosterone but prolonged release of glucocorticoids cause neutrophilia which results in increased phagocytosis and stimulation of immune system (Hall and Goldstein, 1984; Wilckens and de Rijk, 1997). The suppression of immune function and feedback repression of the Hypothalamo-Pituitary-Adrenal axis (HPA axis) by glucocorticoids are mediated through repression of gene transcription (Philips et al., 1997) results in inhibition of synthesis of various cytokines such as IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-8 and IFN-γ, significantly the IL-2 and their receptors enhance the production of other molecules such as lipocortin-1 and type II IL-1 receptor (Nelson et al., 2003). In addition, glucocorticoids exert their anti-inflammatory mechanism by synthesizing lipocortin-1. Lipocortin-1 blocks phospholipase-A2. This blocks eicosanoid synthesis and suppresses many leukocyte inflammatory events (e.g., epithelial adhesion, emigration and chemotaxis, followed by phagocytosis and respiratory burst) (Van de Garde et al., 2014). Apart from suppressing immune response, glucocorticoids inhibit prostaglandins and leukotrienes (the two main products of inflammation).
Glucocorticoids primarily influence the cellular immunity but also suppress the humoral immunity. At physiological concentrations, glucocorticoids favor the Th2 type immunity (Franchimont et al., 2000). Glucocorticoids regulate transcription factors, characteristic of a steroid. Another important feature of glucocorticoids is that they down-regulate the Fc receptors expression on macrophages resulting in reduced phagocytosis of opsonised cells. Glucocorticoids show bi-directional positive and negative actions on the immune system. They act positively on the innate immune response by priming and modulating the genes and by promoting the humoral (Th2) immune response. Glucocorticoids can also act negatively by suppressing the cellular (Th1) immunity on the adaptive immune response. In addition, glucocorticoids provoke the tolerance to specific antigens by controlling maturation of dendritic cells and their function; and cause up regulation of activity of regulatory T cells which is main source of IL-10. Glucocorticoids can be used as an adjuvant in cell therapy for the treatment of autoimmune diseases (Franchimont, 2004). Inhalation of glucocorticoids may be better to dampen eosinophil induced inflammation (especially in cats) thereby normalizing the dysregulated immune system (Chang et al., 2013).
Host defense peptides: Antimicrobial peptides or Host Defense Peptides (HDPs) are considered in the recent times for their immunomodulating properties owing to their both anti-inflammatory and immunostimulatory activities. The HDPs are an evolutionarily conserved molecule of the innate immune system and short cationic peptides of most multicellular organisms. These host defense peptides are produced by all mammals and are mainly produced from their lymph nodes (Hancock and Sahl, 2006; Hamill et al., 2008). Cathelicidins, defensins and histatins are the classical example of the peptides produced by mammals during infection. The antimicrobial actions of HDPs are owing to their amphipathic nature which enables the peptides to act on the lipid bilayers of pathogens and cause their membrane damage, translocate the membrane and inhibit nuclear and protein synthesis machinery of pathogens. As the interactions between the HDPs and the anionic lipids of membranes of microbes are relatively nonspecific, many peptides have showed broad spectrum of antimicrobial activities, targeting both Gram-negative and Gram-positive bacteria, some viruses, protozoa and fungi (Zasloff, 2002; Powers and Hancock, 2003; Nijnik and Hancock, 2009; Sunkara et al., 2012). Furthermore, these peptides have antimicrobial activity by either damaging the membrane of the microbe or by entering the microbes and act inside especially in phagosomes of the host (Powers and Hancock, 2003). The HDPs are greatly effective against multidrug-resistant (MDR) bacterial strains by acting as immunomodulators. The HDPs have capability to modify gene expression of the host, enhance the chemokine production by acting like chemokines, prevent LPS induced pro inflammatory cytokine secretion, alter the dendritic cell responses and cells of the adaptive immune system and induce wound healing (Hunter et al., 2002; Nijnik and Hancock, 2009). Host defense peptides have been synthesized and research has been conducted for their efficacy to possess immuno-modulatory actions. Innate defense regulator 1 (IDR-1), is the first synthesized peptide that boosted host disease resistance against bacterial challenge. The correct mechanism by which this activity is accomplished is not clear, though it has been postulated that it increases cytokine production and also modulate cytokines like TNFα (Scott et al., 2007). RDP58, another peptide that was obtained from Human leukocyte antigen (HLA) 1 molecule, has activity similar to IDR-1 peptide, like decreasing the synthesis of pro-inflammatory cytokines (Easton et al., 2009). The postulated mechanism is that these peptides affect the signaling pathways but the exact picture remains unclear (Travis et al., 2005). There is a list of peptides which are available in the market as immunostimulatory agents and the list include pexiganan (MSI-78), HB-1345, hLF1-11, XOMA-629, iseganan (IB-367) and omiganan (MX-226). Glutoxim a tripeptide which holds anti-infective properties, is used along with traditional medicine for the treatment of Mycobacterium tuberculosis infection (Fimiani et al., 2002). Fowlicidin-3 has the property of inhibiting lipopolysaccharide-induced expression of pro-inflammatory genes in mouse macrophage RAW264.7 cells (Bommineni et al., 2007). In addition, chicken fowlicidin 1 analog showed strong Immunomodulatory activities and protected mice against methicillin-resistant Staphylococcus aureus infections (Bommineni et al., 2014), suggesting development of chicken fowlicidins as a unique antisepsis and antimicrobial agent, especially against antibiotic-resistant microbes. In recent years, proteins isolated from Bacillus cereus have been found to have antifungal activity and are immunomodulatory in nature (Salmen et al., 2013).
MICROBIAL PRODUCTS
Louis Pasteur first suggested in 1885, that the human immune system can be affected by microorganisms. Bacterial immunomodulators are a heterogeneous group of drugs usually composed of standardized lysates or extracts of different bacterial strains used for a non-specific activation of the immune system. They are administrated to increase mucosal immunity and to prevent recurrent infection of the respiratory tract (Spisek et al., 2004). Following exposure to Gram-positive and Gram-negative bacteria (or to the biologically active components of these microorganisms), the functioning of lymphocytes and macrophages may be significantly altered. There is either enhancement or inhibition of the functioning of immune cells by immmunomodulating substances related to bacteria. This increases probability of influencing specific immune resistance of host to infection. Substances derived from such bacteria may exert a continuing influence on the immune response by both specific and non-specific mechanisms due to continual exposure and colonization by bacteria (Friedman et al., 1984).
The microbial products are known for their potential for non-specific activation of macrophages and stimulation of NK cell activity. A wide variety of bacteria have been employed as immunostimulants as most of their cellular components act as ligands for different Toll-Like Receptors (TLRs). As a result, they activate macrophages and dendritic cells which in turn stimulate the release of a mixture of cytokines. The most potent cytokine synthesis enhancer is Bacillus Calmette-Guerin (BCG) which is derived from Mycobacterium bovis the attenuated and live vaccine strain. BCG generally enhances B and T cell mediated responses, phagocytosis, graft rejection and resistance to infection (Barakat et al., 1981; Vetskova et al., 2013). Muramyl dipeptide (MDP) (adjuvant active) is a simple glycopeptide and minimal active structure of whole Mycobacteria endowed with numerous biological activities. By activation of macrophages and enhancement of humoral immunity, MDP increases non-specific resistance against infectious challenges and increases resistance against tumor grafts under certain conditions. The biological activity of MDP is greatly enhanced by incorporation into the lysosomes because of its rapid rate of excretion in urine (Souvannavong et al., 1988). Phosphoglycolipids (PGLs) viz., PGL1 and PGL2 are new class of glycolipids (first isolated in 2006) from the thermophilic bacteria: Thermus oshimai and T. thermophilus, Meiothermus ruber and M. taiwanensis. PGL1 from M. taiwanensis and T. oshimai process the activity to induce proIL-1 in human THP-1 monocytes and blood-isolated primary monocytes but the same is not true in case of T. thermophilus and M. rubber (Yang et al., 2006, 2008).
It is well reported that bacteria can modulate the immune response to non-related antigens; classical example being Propionibacteria which are amongst the most potent immunomodulators stimulating cell populations involved in non-specific resistance. Through mechanisms of recognition and elimination, the activated immune system generally provides protection from infectious pathogens and prevents spread/growth of malignant cells. In experimental and clinical settings, Propionibacteria and its defined low molecular weight substances could be effective in the treatment of infections and neoplastic diseases (Pulverer et al., 1985). Heat killed or formaldehyde treated anaerobic suspension of corynebacteria, such as Propionibacterium acnes is used for immunotherapy. It activates macrophages and clears particulate material from the circulation and presumably stimulates cytokine synthesis through TLRs. It can enhance humoral and cell mediated immune responses and can cause tumor regression. It enhances defense to bacterial, viral and protozoan infections. These organisms have a general immunostimulatory action, leading to enhanced antibacterial and antitumor activity (Becker et al., 1989). Propionibacterium avidum KP-40 is preferably introduced for clinical evaluation on practical ground although its immuno-active capacity is absolutely identical to P. granulosum KP-45. The efficiency of propionibacterial immunomodulation is related not only to the type of tumour involved but also to the bacterial strain used along with route and timing of administration (Szmigielski et al., 1982). Treatment with propionibacteria proves to be of considerable clinical benefit, inducing potent immunostimulation and is essential in colorectal carcinoma wherein Propionibacterium avidum KP-40 (having stimulatory effect on non-specific immunity (Pulverer et al., 1985; Isenberg et al., 1995). Propionibacterium avidum KP-40 also enhances thymus weight and at the same time accelerates maturation of thymocytes. It also enhances significantly absolute counts of peripheral blood lymphocytes and monocytes by inducing expression of activation markers on monocytes viz., interleukin (IL)-2 receptors (on lymphocytes) and macrophage 3 antigen (MAC-3) (Lefrancois and Puddington, 1995).
Staphylococcal cell walls (especially staphylococcal phage lysates), some Streptococcal components and components of Bordetella pertussis, Brucella abortus, Bacillus subtilis and Klebsiella pneumoniae all have immunostimulative activity (Bessler et al., 1991).
Bacterial derived unmethylated cytosine-guanosine nucleotide is the specific ligand to the pattern recognition receptor TLR9 present over the antigen-presenting cells and thereby triggers a potent Th1 cytokine response. If given alone, the nucleotides can act as immunostimulant and can greatly enhance the innate immunity while if given with an antigen these nucleotides act as potent adjuvants (Wang et al., 2006). Certain complex carbohydrates obtained from yeasts viz., zymosan, glucans, aminated polyglucose and lentinans can also activate the macrophages. These may function as adjuvants and potentiate resistance to infectious agents.
Acemannan is obtained from the Aloe vera plant which is a complex carbohydrate and is a potent cytokine synthesis enhancer with antitumor and antiviral activities. Lentinan is a neutral polysaccharide isolated from mycelia of an edible mushroom Lentinus edodes. It can augment antigen specific cellular immune response and also has antitumor action (Scaringi et al., 1988). For various in vitro assays as standards, synthetic lipopeptide analogues derived from bacterial lipoprotein can be used as potent mitogens for lymphocyte activation and also as polyclonal activators to induce immunoglobulin synthesis. They moreover induce tumor cytotoxicity apart from stimulating the secretion of IL-1, IL-6 as well as Tumor Necrosis Factor (TNF) and Nitric Oxide (NO) in monocytes and macrophages; constitute potent immuno-adjuvants in vitro and in vivo. Added advantage is their non-toxicity and long time storage capability even at room temperature thereby making them to meet the requirements for effective standards used in various biological assays (Bessler, 1992). Gram-negative bacterial cell wall primarily consists of lipopolysaccharide made up of Monophosphoryl Lipid A (MLA) which is proved to be a safe and effective compound in inducing immune responses to heterologous proteins in animal and human vaccines (Persing et al., 2002; Bohannon et al., 2013). As TLR4 agonist, it has strong immunomodulatory effects when used both as stand-alone products and vaccine adjuvants and importantly, unique approaches to vaccine manufacturing could be benefitted by both innate and adaptive responses.
Probiotics: Microorganisms serve as an important functioning constituent of the mammalian gastro-intestinal tract. The antibiotic and immunosuppressive therapy can alter the normal flora of the gastrointestinal tract. The introduction of beneficial flora into the system will not only help to reestablish the normal flora but also help to prevent the disease. The word "probiotic" was first introduced by Lilly and Stillwell (1965) in order to define substances produced by one organism which enhance the growth of the other organism. The word "probiotics" was obtained from the Greek word meaning "for life". According to the definition by FAO/WHO probiotics are defined as "Live microorganisms which when supplemented in adequate amounts provide a health benefit on the host".
Probiotics or Direct Feed Microbials (DFM) are naturally existing live microorganisms that provide a positive effect on the physiological status of the host (Behnsen et al., 2013; Hormannsperger et al., 2013; Rask et al., 2013). This is often accomplished by their ability to alter the intestinal microbial balance in a beneficial manner which in turn will improve the health and well being of animals, birds or human beings (Fuller, 2001; Dhama et al., 2008a; Behnsen et al., 2013). Probiotics include bacteria, fungi and yeast. Commonly, apathogenic live bacterial strains consisting of the genus Enterococcus, Streptococcus and Lactobacillus are mainly utilized in poultry and livestock. As growth promoters, probiotics have been found to increase feed conversion efficiency, improve growth performance and improve immune responses in poultry and livestock (Balevi et al., 2001; Dhama et al., 2008a). They are capable of stimulating the immune system of human, animals and birds to fight against infectious agents, tumors and other stress factors (Fuller, 2001; Dalloul et al., 2005; Lee et al., 2007a; Ohashi and Ushida, 2009; Corcionivoschi et al., 2010; Dhama et al., 2011; Behnsen et al., 2013; Jacquet, 2013; Rask et al., 2013; Serban, 2014).
Probiotics play a crucial role in the induction of immune competence in neonates (Balevi et al., 2001). They exert immunostimulatory action by stimulating cell mediated immunity, increasing immunoglobulin and interferon production, activating macrophages, lymphocytes and Natural Killer (NK) cells and by regulating oxidative burst and degranulation of heterophils (Koenen et al., 2004; Dhama et al., 2008a). These probiotic bacteria stimulate the immune response in a strain-specific manner (Rask et al., 2013; Ho et al., 2013). Thus, a combination of strains should be used to provide all the beneficial effect of probiotics (Hsieh et al., 2013). Lactobacillus species have been reported to increase the intra-epithelial lymphocytes of intestinal lymphoid tissue which responds to microbes by secreting immunoglobulin A (IgA) and thereby providing local immunity (Balevi et al., 2001; Haghighi et al., 2006).
Various strains of probiotic bacteria such as Lactobacillus casei, Lactobacillus delbrueckii subsp. bulgaricus, Lactobacillus acidophilus, Lactobacillus plantarum, Lactobacillus paracasei, L. fermentum, L. rhamnosus, L. brevis, L. helveticus, L. reuteri and Bifidobacterium animals vary in their mechanisms of action for immunomodulation (Ho et al., 2013; Hsieh et al., 2013; Rask et al., 2013; Lahteinen et al., 2014). The immunomodulation for these probiotics is reflected in increased secretion of antibodies; induction of cell mediated immunity; increased phagocytic activity of granulocytes, increased expression of activation marker (CD25 and CD45RO) on T cells and natural killer cells and cytokine production etc (Kosaka et al., 2012; Lamprecht et al., 2012; Mokrozub et al., 2012; Tsai et al., 2012; Behnsen et al., 2013; Deng et al., 2013; Dongarra et al., 2013; Lahteinen et al., 2014; Hsieh et al., 2013; Rask et al., 2013). Strains of Leukonostoc and Streptococcus are known potent enhancers of Th1 cytokine. Propionibacterium and Bifidobacterium genera are the potent anti-inflammatory probiotics that induce IL-10 secretion. Lactocepin produced by Lactobacillus casei and Lactobacillus paracasei selectively degrade pro-inflammatory chemokines thus may have therapeutic application in control and treatment of inflammatory diseases like IBD, allergic skin inflammation and psoriasis (Hormannsperger et al., 2013). Bacillus subtilis natto (Sun et al., 2010) or other probiotics have shown to stimulate immune system. Lactococcus lactis activated human plasmacytoid dendritic cells and induced production of interferon thus can play a crucial role in anti-viral immunity (Sugimura et al., 2013).
Dietary oligosaccharide such as galacto-oligosaccharides, fructo-oligosaccharides and pectin-derived acidic oligosaccharides may be helpful in prevention of atopic dermatitis, food allergy and allergic asthma by alteration of the intestinal microbiota or by direct interaction on immune cells or both (Jeurink et al., 2013). Lactoferrin which is an iron binding protein in milk has immune modulating properties and can help in modifying the course of methicillin Resistant Staphylococcus aureus infection in udder (Hwang et al., 2014). Microbes with high CpG motif-rich DNA could better support mucosal functions of intestine in healthy individuals and improve the T-helper 1 (Th1)/Th2 imbalance in allergic diseases (Kant et al., 2014). Interaction of prebiotic like Agave fructans (Agave salmiana) with probiotics Lactobacillus casei and Bifidobacterium lactis are helpful in activation and selective differentiation of immune cells (Moreno-Vilet et al., 2014). Fructans can bind to TLR 2 and also TLR 4 hence can modulate the immunity (Peshev and van den Ende, 2014).
Irrespective of their origin and structure, the common mechanisms involved in probiotic action include: Increased production of immunoglobulins (IgG, IgM and IgA); absorption of antigens released from dead microbes; stimulation of cell mediated immunity; elevated production of vital immune factors (interferons etc.). Further, probiotics can increase macrophage as well as lymphocyte and NK cells activity and up regulate oxidative burst and degranulation of heterophils (Isolauri et al., 2001; Trebichavsky and Splichal, 2006). Saccharomyces boulardii has stimulatory effect on secretory IgA and other secretory components of immunoglobulins in small intestine of rats (Buts et al., 1990). Kaila et al. (1992) reported that human Lactobacillus strain has potentiating effect on the circulating antibody producing cell response against diarrhea cases in humans. Park et al. (2002) reported that encapsulated Bifidobacterium bifidum enhances intestinal IgA secretion. Modulation of humoral immunity by probiotics and its effects on cytokine production are depicted in Table 6 and 7.
| Table 6: | Probiotic modulation of humoral immunity |
 | |
| Table 7: | Probiotic effect on cytokine production |
 | |
SYNTHETIC CHEMICAL COMPOUNDS
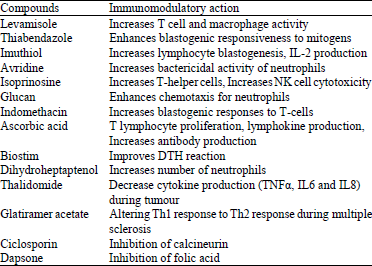
Various chemicals having immunomodulatory properties have been reported in literature (Sultana and Saify, 2012). These compounds offer the advantage of oral administration. It is important, in this regard, to note that cell surface carbohydrates such as glycoconjugates and oligosaccharides are responsible for several vital life processes. These include; immune response, cell-cell interactions, cell growth and proliferation and combating numerous bacterial and viral infections. Recognition of structurally well-characterized and chemically pure carbohydrate antigens is necessary to understand the molecular mechanisms and to study the carbohydrate recognition and subsequent immunomodulation in various diseases. Lipid A analogues, such as monophosphoryl lipid A, have been developed as an immunomodulatory compound with reduced pro-inflammatory activities, thus can be used as an adjuvant in vaccine preparation (Bohannon et al., 2013). Chemically synthesized glycoconjugates are thus valuable to understand the role of cell surface carbohydrates in the pathogenesis of a disease. The important immunomodulatory chemicals which are being used widely are summarized in Table 8.
Various drugs which are chemically synthesized have immunomodulatory actions. The important drugs in this group of chemical agents include thalidomide and its versions namely lenalidomide and pomalidomide which are commonly used in the field of oncology (Zimmerman, 2009). These immunomodulatory agents are usually given orally. They act on cytokines involved in inflammation, molecules involved in expression and modulation of immune cells and finally in angiogenesis (Anderson, 2005). Thalidomide is the first among this group which was available in the market early in 1960s as a sedative and anti emetic but was soon withdrawn because of its side effects like teratogenicity (Eisen et al., 2000; Kumar and Anderson, 2005). Its usefulness as an immunomodulatory agent was understood late in 1990s and hence it was brought back into the market not as an anti emetic or sedative drug but as a drug against erythema nodosum leprosum. Thalidomide is the parent compound derivative of glutamic acid. Lenalidomide and pomalidomide are the derivatives of thalidomide but have higher activity as compared to the parent compound (Chanan-Khan et al., 2013). Thalidomide became a promising drug for the treatment of multiple myeloma condition after a series of clinical trials.
| Table 8: | Synthetic chemical compounds having Immunomodulatory activities |
 | |
Several studies have also been conducted for its effect on other tumorous conditions like breast cancer, renal carcinoma, Kaposi’s sarcoma, malignant melanoma, CNS malignancies and prostrate carcinoma (Fife et al., 1998; Escudier et al., 2002). But its activity was found to be more against multiple melanoma as compared to the other conditions (Garcia-Sanz, 2006). Mechanism of action of thalidomide and its derivatives is of two modes, one by directly acting on the tumor cells and another by down regulating their functions (Figg et al., 2001; Short et al., 2001). The first mode of action of thalidomide is done by decreasing the production of various cytokines which are involved in survival of the tumor cells like TNF-α, IL6 and IL8. TNF-α production is decreased by two ways, one by degradation of the mRNA which is intended to produce TNF-α. Alpha-1-acid glycoproteins are produced in excess which has anti TNF-α activity and hence it also helps to reduce the production TNF-α (Baidas et al., 2000). The second mode by which thalidomide helps to keep tumor under control is an indirect way where it makes the interaction possible between the tumor cells and the nearby micro environment. Thalidomide group of drugs can also activate Caspase 8 which can cause destruction of myeloma cells through Fas mediated pathway (Latif et al., 2012).
Multiple sclerosis is a common demyelinating condition of the central nervous system (CNS) which occurs throughout the globe. Various drugs have been used against this condition and only a few drugs including immunomodulatory agents have yielded good results. Glatiramer acetate is a chemical having structural similarity with the myelin basic protein and it is formed by alanine, lysine, glutamate and tyrosine amino acids arranged randomly. Mode of action of this chemical is by altering Th1 response to Th2 response during multiple sclerosis condition (Duda et al., 2000). Natalizumab is another chemical which is synthesized to α4 portion of the α4β1 integrin and its mechanism of action is preventing the migration of lymphocytes during inflammation along the Blood Brain Barrier (BBB) to the CNS (Steinman, 2005). Fingolimod is a drug intended for oral use acting on the sphingosine 1-phosphate (S1P) receptors preventing the movement of lymphocytes (Hanel et al., 2007). Teriflunomide, Laquinimod and Cladribine are the other drugs that can be used against multiple sclerosis owing to their immunomodulatory action (Nicholas et al., 2011).
Certain chemicals and drugs that are either naturally obtained or synthesized artificially have been found to have immunosuppressive property that are used in various conditions including organ transplantation (Georgakopoulou and Scully, 2013). Cyclosporin obtained from the fungus Tolypocladium inflatum has immunosuppressive properties which changed the course of transplantation biology. Mechanism of action is by inhibition of calcineurin which occur by binding of cyclosporin to immunophilin, an intracytoplasmic protein (Tedesco and Haragsim, 2012). Tacrolimus, a macrolide obtained from Streptomyces tsukubaensis, has similar mode of action to cyclosporin and has no antibiotic activities (Jacobson et al., 1998). Tacrolimus is more potent immunosuppressive agent compared to cyclosporin but the bioavailability is less, hence dosing of tacrolimus is 4 fold higher than cyclosporin (Hoorn et al., 2012). There are some adverse effects of these two drugs like hepatotoxicity, nephrotoxicity and hypertension, so patients need complete monitoring. Azathioprine is a purine derivative anti proliferative cytotoxic drug having action on purine synthesis thereby causing severe damage to the DNA (Saroj et al., 2012). Several steroid drugs are used to suppress immunity hence preventing graft rejection and Prednisolone is one such important drug (Saroj et al., 2012). Dapsone a well known drug used for the treatment of leprosy has immunomodulatory action and hence it is used for conditions like dermatitis herpetiformis (Cardones and Hall, 2012). Its mode of action is by inhibition of folic acid synthesis. Colchicine, an alkaloid derived from Colchicum autumnaleit blocks mitosis by binding tubulin in the microtubules (Molad, 2002). It is commonly used in auto immune disorders, gout, Behcet’s disease and recurrent aphthous stomatitis (Brocklehurst et al., 2012). Cyclophosphamide is another agent which also has immunosuppressive action and used mostly in neoplastic conditions that are malignant in nature (Mukhtyar et al., 2009). An antiparasitic drug, Levamisole has been mostly studied for its immune stimulant properties. It has various potential immune stimulating properties and to name some; it activates macrophages and monocytes thereby increasing the phagocytosis in the body, T cells are activated, proliferated and also enhances antibody production to various antigens (Hegde et al., 2012). Levamisole can show its action in diseased as well as normal immune system hence it has both immune stimulant and immune regulatory actions. Its action on T cells is comparatively higher than on B cells. Levamisole has been used to stimulate immune cells in various cancers in human patients (Stevenson et al., 1991; Friis et al., 2005).
HERBAL PRODUCTS
Since long time, modulation of immune response to alleviate diseases using herbal medicines has been a form of therapy for livestock, particularly among the resource poor marginal farmers and in this aspect the traditional medicinal plants are especially important (Hasani-Ranjbar et al., 2012; Mahima et al., 2012a; Mirzaei-Aghsaghali, 2012; Tiwari et al., 2014a, b). In different parts of the world, plant extracts have been widely investigated for their possible immunomodulatory properties (Alamgir and Uddin, 2010). In Indian traditional medicine, several medicinal plants called ‘Rasayanas’ are known to increase the resistance of the body against a variety of infections and thus have attracted the attention of many scientists. Macrophages facilitating generation of an immune response are the main target of the immunomodulatory plant products. The activated macrophages cause increased phagocytosis and generate effector molecules like free radicals, nitric oxide and cytokines that facilitate intracellular killing of pathogens. These cytokines may have direct function or may affect the function of other immune cell population such as induction of natural killer cell mediated cytotoxicity or production of cytotoxic T lymphocytes. The plant derived immunomodulators have tremendous potential for generation of new pharmaceutical products (Tan and Vanitha, 2004; Chen et al., 2014; Mahima et al., 2013b). Extract of fresh ajoene (Allium sativum) protects CD8+cells from attack by HIV at low concentrations early in the viral life and has little toxicity with 45 times more powerful effect than dextran sulfate. A. sativum causes impairment of the activity of the liver enzymes (that process protease inhibitors) raising their level. Allium sativum extract (GE) has dose-dependent inhibitory effect on human cytomegalovirus (HCMV) in vitro (Guo et al., 1993).
Progress on the use of ethnomedicinal plants as immunomodulatory agents has been more pronounced recently as plant extracts have widely been explored during last few decades in various parts of the world for their possible immunomodulatory properties. Many studies have demonstrated the isolation of potential bioactive molecules which have been patented and have been tested as herbal formulations (Carrio et al., 2012). Sunila and Kuttan (2004) reported immunomodulatory and anti-tumour activity of Piper longum Linn and piperine. Botanicals like C. versicolor etc., are rich in glucans, having potent immunomodulatory action. Polysaccharides isolated from Artemisia apiacea can inhibit growth of hepatoma cells by stimulating apoptosis and immuno-defense (Chen et al., 2013). Morinda citrifolia (Noni) fruit juice contains a polysaccharide-rich substance which enhances the IFN-γ production from thymocytes. Noni (M. citrifolia) potentiates the immune system by activating the macrophages to secrete TNF-α, IFN-γ, IL-1β, IL-10, IL-12 and nitric oxide but it suppresses the release of IL-4. Hence, Noni reduces the tumour growth by potentiating the host immune system (Hirazumi and Furusawa, 1999). Noni juice potentiates the immune system by increasing the weight of the thymus resulting in protection from degenerative disease and aging (Pansuebchue et al., 2002). Feeding of Noni fruit juice to neonatal Holstein calves potentiates the immune system by increasing the expression of CD25 on CD4+, CD8+ and γδ T cells. Noni also increases the secretion of IL-1β, TNF-α and IFN-γ in bovine colostrums and results in enhancement of natural innate cell-mediated immunity (Brooks et al., 2009). The fermented Noni Exudate (fNE) treated dendritic cells cause immunoglobulin class switching to produce IgG and IgM, proliferation of splenocytes and B cells and promotes their differentiation (Zhang et al., 2009). The extract of M. citrifolia fruits enhances the activity of T and B lymphocytes which potentiates the both arms of the adaptive immune system such as humoral and cell mediated responses (Nayak and Mengi, 2010). Aqueous extracts of Withania somnifera show potent immunomodulatory properties (AbdEIslam et al., 2013). Mostly secondary metabolites of plants exert beneficial immunomodulatory effects many of which have been widely studied in mouse as well as chicken and human cell lines. Ginseng and steroidal saponin present in it, for instance, has immune-stimulating properties that include; Cytokine production (IL-2, IL-6,TNF-α and IFN-α); macrophage activation and lymphocyte stimulation (Lee et al., 2007b). Flavonoids and terpenes from Ginko biloba (Ginseng) conversely can mediate production of inflammatory cytokines. For stimulating cell-mediated immunity and to enhance antibody production, saponins are useful and can induce production of cytokines viz., interleukins and interferons. Meyer saponins and Quillaja saponins; butanol extract of Lonicera japonica and de-acetylated saponin-1 can be administered to the nasal mucosa to stimulate the immune response in vivo. Considerable improvement in antibody titre is caused by herbal plant polysaccharides obtained from four Chinese herbs viz., roots of Astragalus, Isatis and Achyranthes; and Chinese Yam (Hashemi and Davoodi, 2012; Chakraborty and Pal, 2012). Aquous extract of C. nuda (an edible mushroom) induces dendritic cell maturation through TLR-4 and/or TLR-2 suggesting its role in cancer vaccine immunotherapy (Chen et al., 2014). Eupatorium adenophorum polysaccharide increases the production of IL-6, TNF-α and IFN-γ suggesting its immunomodulatory properties for prophylaxis of H5N1 influenza infection (Jin et al., 2013). Naringenin and flavones are certain flavonoids which are found to be effective against filarial worms like Brugia malayi (Lakshmi et al., 2010). Nigella sativa has anti inflammatory effects and its active principle thymoquinone limits production of 5-lipooxygenase and 5-hydroxyeicosatetraenoic acid (Hajhashemi et al., 2004; Elkamel and Mosaad, 2012). Herbs have been found useful in amelioration of the immunosuppressive effects of chicken infectious anaemia virus in poultry (Bhatt et al., 2013). Various herbal products which can be used as immunomodulators and herbal preparations containing them are depicted in Table 9 and 10.
| Table 9: | Herbal products and their immunomodulatory activities |
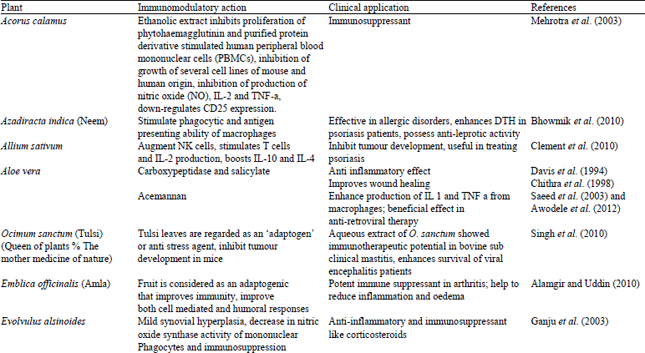  | |
| Table 10: | Herbal preparation for immunomodulation |
 | |
Essential oils from various herbal extracts possess the unique property to modulate immunity (Gopi et al., 2014). Carvacrol and thymol are the two main such extracts obtained from oregano and thyme. These oils in equal composition together have shown to protect pigs from pathogenic diseases (Walter and Bilkei, 2004). There was increase in CD4 and CD8 cells after giving these oils to the pigs which indicates its immune modulating potential. Thymol has anti inflammatory properties locally which are evident by the reduction of TNFα and it also increases the immunoglobulins like IgA and IgM (Trevisi et al., 2007). Cinnamon when extracted produces cinnamaldehyde which do have immune modulatory properties like other essential oils. Echinacea purpurea extract from plants of Echinacea shows to improve feed efficacy in animals (Maass et al., 2005). Immunity of animal was increased in pigs which were vaccinated against Erysipelothrix rhusiopathiae after administering extracts of Echinacea purpurea. Several Chinese herbs were also studied for their immune modulatory properties and one such Astragalus membranaceus showed to increase leucocyte count especially CD4 cells (Yuan et al., 2006). Soyabean extracts has two isoflavones namely genistein and daidzein have immune modulatory activities (Greiner et al., 2001). Auraptene extracted from a citrus fruit peel has property of enhancing macrophage and lymphocyte activities. Ethanolic extract of Allium hirtifolium Boiss has shown to reduce acquired immunity in a concentration dependent manner (Sharma et al., 2011). Extracts of Randia dumetorum Lamk increases both humoral and cell mediated immunity in mice model. Extract of Cleome gynandra Linn. has immune suppressive activity (Gaur et al., 2009). Apart from oils from herbs, fish oil with emulsion of lipids also has anti inflammatory properties (Hecker et al., 2014).
An illustration depicting an overview of immunomodulation, immune mechanisms and modes of action of various immunomodulatory agents is presented as Fig. 1.
ADJUVANTS
Adjuvant is a term derived from the Latin word Adjuvare, meaning to help or to aid or to enhance (Kumar et al., 2013) which was coined by Ramon (1926), who reported that horses that developed abscesses following injection of diphtheria toxoid produced higher antitoxin titers than animals without abscesses (Ramon, 1926). Jolles and Paraff (1973) defined adjuvants as any substances which act on a hapten or antigen enhancing its antigenic properties or which act on the cells involved in the immune response. Vaccine adjuvants are a group of structurally heterogeneous compounds that may stimulate or modulate the immune system thereby enhance specific immune responses against co-administered antigens without having any specific antigenic effect itself (Petrovsky and Aguilar, 2004; Singh and O'Hagan, 2003). Enhancing host defenses is a highly efficient approach to counteract infectious diseases either through vaccination as a means of prevention or therapeutically by the use of immunomodulators. Different vaccines currently in use are effective partly due to molecules that possess low immunogenicity by themselves but which may help to increase and modulate the immune response to an antigen which ischaracteristic feature of adjuvants.
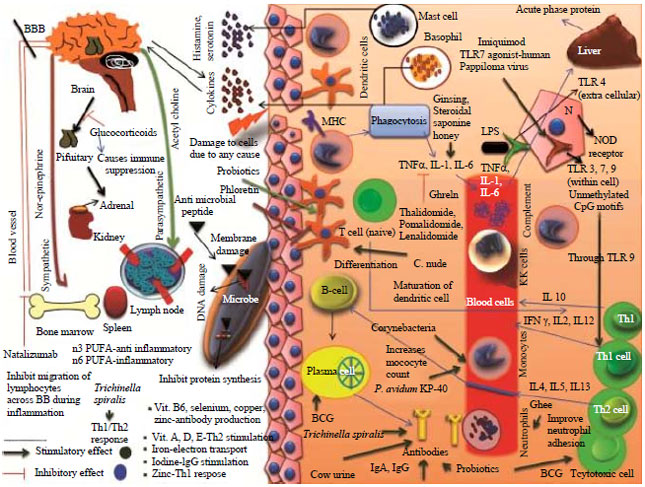 | |
| Fig. 1: | An overview of immunomodulation and modes of action of various immunomodulatory agents |
There is a wide array of adjuvants that are being developed and used, from a variety of sources (Sun et al., 2009). Despite a plethora of options, only aluminium salts have gained wide acceptance as human and veterinary vaccine adjuvants (Edelman, 2002). Adjuvants increase the humoral as well as T cell mediated immune responses by enhancing antigen presentation, activating dendritic cells, increasing the inflammatory cytokine production etc (McElhaney et al., 2013).
The immunostimulatory property of aluminium salts is dendritic cell (DC)-dependent (Kool et al., 2008a); whereas the T-cell response partially depends on Nalp3 inflammasome activation (Eisenbarth et al., 2008). Nalp3 is an intracellular recognition receptor and is a member of the NOD-like receptor (NLRs) family. Nalp3 can recognize stimuli from microbial entry and origin and endogenous molecules produced during the cellular damage. Along with Apoptosis-associated Speck-like Protein (ASC) and caspase-1, it forms the inflammasome and controls the cleavage and secretion of the strong pro-inflammatory cytokines like interleukin (IL)-1b, IL-18 and IL-33 (Sutterwala et al., 2006, 2007). Induction of Nalp3 inflammasome is done by alum thereby inducing the secretion of pro-inflammatory cytokines IL-1b and IL-18 leading to strong production of uric acid. Crystals of monosodium urate and aluminium containing adjuvants activate the differentiation and recruitment of inflammatory monocytes (F4/80int CD11b+Ly6G-Ly6C+) that differentiate into inflammatory dendritic cells and induce CD4+T-cell activation and multiplication results in antibody secretion (Kool et al., 2008b). Saponin-based adjuvants are obtained from the bark of the quillaja tree and possess immunomodulatory properties of activation of a balanced TH1/TH2 response with antibody (Maloy et al., 1995) and cytotoxic CD8+lymphocytes production (Lipford et al., 1994). Cationic liposome formulations (CAF) having positive surface charge with lipid bilayer vesicles cannot be adequately immunostimulatory thus can be administered along with immunostimulators viz., α, α-trehalose 6, 6'-dibehenate (TDB). TDB (adjuvant CAF01) is a synthetic mycobacterial cord factor analogue and act as a stabilizing agent on liposomes and prevents precipitation and aggregation in suspension (Agger et al., 2008). It also has potential adjuvant action in animal model in tuberculosis (TB) vaccine. The CAF01 adjuvant is considered to be safe and possesses short immunogenicity in Phase I studies. The experimental studies showed that the CAF01 adjuvanted TB vaccine enhances depot formation and prolonging antigen release while targeting the antigens and immunomodulator to the same activated antigen presenting cells (APCs) (Kamath et al., 2009). Along with efficient polyfunctional memory T cells both humoral and cellular immune responses are stimulated by such preparation (Lindenstrom et al., 2009). In order to obtain well-defined non-toxic adjuvants and antigens which can be used for human vaccination and immunostimulation, efforts have been made for several years. One of the most studied family of compounds is Muramyl dipeptide (MDP) (synthetic adjuvant) that has got other pharmacological properties viz., pyrogenic and somnogenic activities. Several hundreds of MDP derivatives have been synthetized and some of the biological ineactivities have been dissociated. One MDP derivative presently under clinical trials has been shown to be active adjuvant but devoid of pyrogenicity (Leclerc and Vogel, 1985).
A widely used vector is adenovirus serotype (Ad) 5 as an adjuvant and has been investigated for the potential to deliver human immunodeficiency virus (HIV-1) gene products and stimulating HIV-specific immune responses in human clinical trials (Cheng et al., 2010). In some instances, vector immunity can lead to reduction in CD4 and CD8 immunity and decrease in vaccine efficacy. To avoid pre-existing immunity due to Ad5 vaccination, diverse vaccination strategies are used such as prime-boost approaches and increased vaccine doses. An encouraging approach is the use of replication-defective recombinant lymphocytic choriomeningitis virus (rLCMV) vectors wherein vaccine gene replaces the LCMV glycoprotein gene to generate replication-defective vaccine vectors. For producing tetramer specific CD8+T-cells response in mice rLCMV is 1000-fold more potent than rAd5 (Flatz et al., 2010).
F3, a high molecular weight polysaccharide extract of Ganoderma lucidum, has been comprehensively analysed for adjuvant and immunomodulatory activities both in vitro and in vivo studies. F3-treated mice have shown more number of dendritic cells and other cells of the immune system during in vivo experiments. These cells viz., CD4+ and CD8+ cells, regulatory lymphocytes and plasma cells, NK and NKT cells elevate the quantity of multiple chemokines and cytokines in the spleen and blood of mice. F3, in addition, possesses strong adjuvant activity in tetanus toxoid in the lack of alum and enhances antibody responses to alum-formulated tetanus toxoid in mice and boost Th1 and Th2 response. F3 in vitro can induce dendritic cell maturation derived from human monocytes by up regulating CD40, CD54, CD80, CD83, CD86 and HLA-DR; enhance mixed lymphocyte reaction and stimulate cytokine and chemokine production (Lai et al., 2010). Appreciable success has been possible by using many peptide antigen based formulations. Nowadays peptide-based vaccines are arising as the next generation of prophylactic and corrective immunotherapy. But the major disadvantage is that peptide- based vaccines are poorly immunogenic and require strong immunostimulatory adjuvants which can make active both innate and adaptive immune systems. Studies have thus been conducted to create ideal peptide antigen delivery systems by incorporating multiple pattern-recognition receptor (PRR) agonists. Nowadays peptide antigen delivery systems are designed to stimulate Toll-like receptors (TLRs) on dendritic cells (DCs) and evaluation of the profiles of various cytokines (viz., IL-4, IL-6, IL-10, IL-12, IL-23 etc. of DCs) induced by individual TLR agonists. TLR agonists are commonly used as immunostimulatory agents, for example imiquimod an agonist of TLR 7 is approved for use against Human pappiloma virus topically (Romagne, 2007; Kanzler et al., 2007; Malik et al., 2013). In predicting the influence of specific TLR agonists on a particular T helper cell (Th) response such studies are helpful (Mitchell et al., 2011). Innate immunity also known as non-specific or semi specific immunity because it differentiates self from microbes through toll-like receptors (TLRs) which are evolutionarily conserved receptors. Recent research evidences have shown that TLRs can act as adjuvant receptors that act as bridge between innate and adaptive immunity, resulting in stimulation of adaptive immunity (Mutwiri et al., 2007; Akira, 2011). TLR 7 agaonist Imiquimod, has been approved by FAO for use against Herpes sarcoma virus-2 (HSV-2) (Prins et al., 2006). Similarly LPS which act against agonist of TLR 4 are also used against glioblastoma multiforme tumours in mice models (Chicoine et al., 2007; Deng et al., 2014). TLR 9 agonist has been used in mice models against brain tumours which ftlineresulted in apoptosis of glioblastoma cell (El Andaloussi et al., 2006; Rookhuizen, 2012). Stils (2005) described adjuvants and antibody production that has dispelled the myths in relation to Freund’s complete as well as other adjuvants. Conjugation of monophosphoryl lipid A and oligodeoxynucleotides to nanolipoprotein significantly increased/activated the production of cytokine, cell surface expression of activation markers and upregulation of immunoregulatory genes (Weilhammer et al., 2013). Curdlan sulfates i.e., synthetic product of curdlan with sulfur trioxide-pyridine complex was found effective in increasing the Nitric Oxide (NO) release and the cytokines TNF-α, IL-6 and IL-1β production by macrophages in mice model and can be developed as a new immunotherapy agent and anti-viral vaccine adjuvant (Li et al., 2014). Recently, high mobility group box 1 (HMGB1) protein has been found to act as a novel immunoadjuvant generating highly protective immune responses during DNA vaccine trial of chicken infectious anemia virus in poultry. The most common adjuvants which are used for veterinary vaccine production are depicted in Table 11.
Role of adjuvants in immunomodulation/ veterinary vaccine development: Adjuvants enhance the host immune response against vaccine antigens in many different mechanisms as mentioned below:
| • | Enhance the immunogenicity of weak antigens |
| • | Increase the duration and speed of the immune response |
| • | Modulate avidity as well as specificity and isotype or subclass distribution of antibodies |
| • | Enhance strong Cell Mediated Immunity (CMI) |
| • | Stimulates the mucosal immunity |
| • | Augment immune responses in immunologically immature (newborn) or senescent and compromised individuals |
| • | Reduce the antigen dose or the number of immunizations needed for protective immunity to reduce the vaccine costs |
| • | Help to avoid antigen competition in combination vaccines |
| • | Increase the overall antibody titer or functional titers |
| • | Induce broader immune response (cross-protection) (Vogel and Hem, 2004) |
| Table 11: | Adjuvants for veterinary vaccine production |
 | |
Mode of action of adjuvants: The identification of adjuvants has largely been factual for decades but a series of new adjuvants and novel formulations are upcoming nowadays that can act through immunomodulatory mechanisms. They direct immune responses against disease causing organisms and enhance memory responses. It is thus beneficial for vaccine design and to build new model to assess adjuvant safety at regulatory level and at development stages (Mastelic et al., 2010). Adjuvants enhance the immune response by various mechanisms that include antigen presentation and targeting, "depot" effect, immunomodulation as well as cytotoxic lymphocyte induction (Cox and Coulter, 1997). "Depot" effect is a classical mechanism of action of adjuvant that helps the adjuvant to protect the antigen from rapid degradation and removal by the host due to slow release of intact antigen, thereby, permitting a prolonged and slow exposure of the cells of the immune system to a low level of antigen. Furthermore, adjuvants protect the antigen from proteolytic destruction by carrying the vaccinal antigens. Adjuvant can also preserve and maintain the conformational integrity of antigen and enables to present the antigen to professional antigen-presenting cells such as dendritic cells and macrophages (Stils, 2005; Leroux-Roels, 2010; Noe et al., 2010; De Gregorio et al., 2013). For designing vaccine, it is necessary to understand mechanism of action of adjuvant which helps in immune responses directly towards effector mechanisms which should be efficacious as well as disease-specific in nature and could provide appropriate memory (Mastelic et al., 2010).
Risk of adjuvants: From the past 70 years various kinds of adjuvants have been made but they were not used in routine vaccination programmes due to their immediate toxicity and delayed effects which include the following:
| • | Localized acute or chronic inflammation that may lead to painful abscess formation |
| • | Formation of persistent ulcers, nodules and draining lymphadenopathy |
| • | Fever with influenza like illness |
| • | Toxicity to organs and tissues due to chemicals of adjuvants |
| • | Hypersensitivity reactions to host tissue that produces anterior uveitis, amyloidosis and autoimmune arthritis |
| • | Cross reaction with host tissue antigens results in meningoencephalitis and glomerulonephritis in humans |
| • | Oral tolerance and immune depression |
| • | Carcinogenic potential |
| • | Teratogenic potential |
These adverse effects can be reduced by imparting regulatory control over use of adjuvant, developing relatively non-toxic synthetic immunoregulators and by standardizing safety and potency tests (Edelman, 2002).
NUTRIENTS
It is well known that nutrition plays a significant role in immunomodulation and malnutrition is the most common cause of immunodeficiency worldwide. Thus, for the proper functioning of the immune system, adequate supply of nutrients is must and both deficiencies and excesses of nutrients adversely affect the various components of the immune system. This is because of the fact that optimal functioning of immune system involves a variety of biological activities which include cell growth, energy metabolism, production of proteins and antioxidants (El-Gamal et al., 2011; Mahima et al., 2013a; Rahal et al., 2014a, b). Apart from normal nutrition, various proteins and peptides derived from hen’s egg have biological properties like antimicrobial, antioxidant and immunomodulatory that regulate different functional systems of the body (Yu et al., 2014). Regarding the amino acids role for the oxygen dependant killing of microbes in the phagocytic cells, arginine (a direct precursor of nitric oxide and a effective killer) is important (Duff and Daly, 2002). Glutamine is essential for appropriate functioning of the lymphocytes and macrophages and induction of the immune system during the inflammation. It has also been reported that the utilization of glutamine by macrophages and lymphocytes during inflammation is high. Glutamine is also necessary for secretion of cytokines and antibodies as well as for the cell division (Calder and Field, 2002). In the pregnant animals both low and high protein: {softlinecarbohydrate ratios in the diet had significant effect on the immune system of the offsprings (Tuchscherer et al., 2012).
Essential fatty acids in the food may enhance the immune system and maintains health (Pond, 2005). In the diet n-6 and n-3 are two major classes of polyunsaturated fatty acids (PUFA). Linoleic acid is precursor of n-6 and present in the vegetable oils, corn and soybean. In the plasma membrane of immune cells linoleic acid is converted into arachidonic acid. So, the n-6 PUFA is inflammatory in action which is opposite to n-3. These findings denote that diet more in n-3 PUFA reduces inflammation by enhancing the docosahexanoic acid and eicosapentanoic acid levels in the plasma membrane by inhibiting arachidonic acid (Mantzioris et al., 2000; Calder, 2001). n-3 PUFA reduces inflammatory and autoimmune disorders (Miles and Calder, 2012; Pae et al., 2012). Fatty acids and amino acids along with other secondary plant substances viz., carotenoids, flavonoids and spices inhibit the release of prestored mast cell mediators like histamine or de novo expression of mast cell mediators such as cytokines and eicosanoids thus can be used in prevention of allergic diseases (Hagenlocher and Lorentz, 2015). Micronutrient deficiency inhibits immune functions by affecting the innate and adaptive immunity which finally leads to dysregulation of the optimal host responses. This increases the susceptibility to various microbial infections, with increased morbidity and mortality (Wintergerst et al., 2007). The micronutrients which are important for immune function comprise of vitamins A, C, E and B6, copper, folate, iron, selenium and zinc. Other nutrients such as beta-carotene which is precursor of vitamin A, vitamin B12 and vitamin D play a significant role in immune function. Generally, they contribute to the body’s natural defenses on three levels by influencing physical barriers (skin/mucosa) and other innate immune cells, cell mediated immunity and humoral immunity. To enhance skin barrier function nutrients like vitamins A, C, E and the trace element zinc are essential. Vitamins A, C, D, E, B6, B12, folic acid and trace elements like copper, iron, selenium and zinc act synergistically to maintain the defensive activities of the immune cells. For antibody production all these micronutrients (except iron and vitamin C) are necessary. Copper, selenium, zinc and vitamin B6, have significant role in B-cell proliferation and antibody production. Vitamins A, D and E potentiates Th2 response which leads to augmentation of humoral immunity and other micronutrients indirectly involved in protein synthesis and cell growth (Chandra, 2002; Maggini et al., 2007). In addition, several studies have shown that a four carbon short chain fatty acid, butyrate and its analog phenylbutyrate enhance disease resistance against shigellosis in humans and rabbits, respectively, by enhancing host defense peptide synthesis (Raqib et al., 2006; Sarker et al., 2011). Moreover, vitamin D and phenylbutyrate are effective in controlling Mycobacterium tuberculosis in human studies by enhancing host defense peptide, cathelicidin expression in macropahges (Mily et al., 2013). Furthermore, butyrate and other fatty acids enhance disease resistance of chicken against Salmonella Enteritidis by enhancing host defense peptide synthesis (Sunkara et al., 2011, 2012). Recently, short chain fatty acids have also shown their ability in enhancing host defense peptide synthesis in porcine intestinal epithelial cells and alveolar macrophages, suggesting the importance of fatty acids as immunostimulators both in human and animals (Zeng et al., 2013).
As a matter of fact, these micronutrients also act as antioxidants and cofactors in the cytokine production and regulation. Micronutrient deficiency and chronic under nutrition nevertheless can suppresses cytokine production and disturb immune cell trafficking and in association with infection further impairs the immune response. Malnutrition at an early age might have long-term effects on health and results in modified immune cell populations and a generalized production of more inflammatory mediators as the immune system is immature at birth. Zinc deficiency and protein calorie malnutrition stimulates the hypothalamic-pituitary-adrenal axis resulting in increased production of glucocorticoids. Increased levels of circulating glucocorticoids result in thymic atrophy which inturn affects hematopoiesis. Leptin is a potent cytokine-like immune regulator that exerts complex effects like inflammatory response in both malnutrition and over nutrition (Cunningham-Rundles et al., 2005). A water-soluble polysaccharide (JS-MP-1) obtained from Korean mulberry fruits Oddi (Morus alba) has immunomodulation property by stimulating macrophage to release chemokines and proinflammatory cytokines (TNF-α and IL-6), thus can be used as immunotherapeutic adjuvant (Lee et al., 2013). Phloretin, a natural dihydrochalcone found in many fruits has ability to suppress the activation and function of dendritic cells in mice thus may be useful in inflammatory and autoimmune disorders (Lin et al., 2014).
Vitamins and immune function: Vitamins are vital amines and have crucial role as nutrients. Apart from this, these play important role in regulation of the host immune defense and balancing the physiologic and pathologic conditions (Kunisawa and Kiyono, 2013). Vitamin A plays an important role in maintaining the integrity of mucosal surfaces in the respiratory and gastrointestinal tracts and in the regulation of innate and adaptive immune response (Villamor and Fawzi, 2005). Vitamin A and its metabolites like trans retinoic acid and retinol; retinal and retinoic acid play essential roles in both cellular and humoral immunity. They also enhance phagocytic activity and regulate immune homeostasis via the peripheral induction of regulatory T-cells. Further, they can control CD4 T-cell differentiation; mucosal immunity and immune tolerance and induces regulatory and inflammatory responses (Prabhala et al., 1991; DePaolo et al., 2011; Cassani et al., 2012). Vitamin A deficiency results in decreased phagocytic activity and diminished oxidative burst activity of activated macrophages during inflammation (Ramakrishnan et al., 2004) and a decreased number of natural killer (NK) cells and their activity (Dawson et al., 1999). Vitamin A deficiency strongly impairs the humoral immunity (Long and Santos, 1999) and provokes an inflammatory state in the body due to enhanced secretion of TNF-α and IL-12. But Vitamin A supplementation can restore these actions (Aukrust et al., 2000).
Vitamin D, especially 1,25-dihydroxycholecalciferol (1,25(OH) 2D3)) is biologically active and is considered to be an important immunoregulator besides its role in calcium metabolism. Most cells of the immune system like T lymphocytes and macrophages (except B cells) express vitamin D receptors in significant concentrations (Veldman et al., 2000; Mocanu et al., 2013). Vitamin D performs its immunomodulatory action by inhibiting excessive production of inflammatory cytokines and enhancing the oxidative burst activity of macrophages. Additionally, it also induces the secretion of potent anti-microbial peptides in most of the immune competent cells (Cannell et al., 2006). Deficiency of vitamin D leads to development of autoimmune diseases and other inflammatory diseases like multiple sclerosis (MS), chronic rhinitis, tonsillar hypertrophy due to increase in inflammatory substances (tumor necrosis factor alpha, IL-1 and prostaglandin D2) (McCarty et al., 2013; Mocanu et al., 2013). Vitamin D also has got significant effect on immune as well as neural cells (Fernandes de Abreu et al., 2009). It also plays an important anti-neoplastic role and is used for anticancer immunotherapy. Vitamin D binding protein (DBP) is an essential precursor for macrophage activation and is crucial for providing innate defense against cancer (Karbasi and Saburi, 2012; Vuolo et al., 2012).
Vitamin E has a strong lipid soluble antioxidant activity that scavenges free radicals and can enhance both the cell division and cytokine secretion from naive T cells but not by memory T cell. Vitamin E can also suppress a variety of inflammatory processes by blocking the activity of transcription factor NFkB which is essential for transcription of many proteins, especially pro-inflammatory cytokines. Vitamin E favors the immune response which is biased towards Th1 immunity (Meydani et al., 2005).
Leukocytes are enriched in Vitamin C which is a major water-soluble antioxidant in cells and plasma. Vitamin C performs its antioxidant activity by scavenging Reactive Oxygen Species (ROS) generated during the process of phagocytosis by activated immune cells (Puertollano et al., 2011). Supplementation of vitamin C results in enhancement of various components of immune response and therefore besides its metabolic functions, its significant role in immune homeostasis cannot be overlooked (Jacob et al., 1991; Hartel et al., 2004). Vitamin C is essential in order to safeguard the host cell from free radicals produced during the respiratory burst from phagocytes. Lack of sufficient quantity of this vitamin in diet results in altered phagocytosis and more free radical mediated injury to the host cells. After the administration of the ascorbic acid the condition changes as there are enhanced sensitivity of B cells to the mitogen as well as it is responsible for DTH and prevention of T cell apoptosis. An improvement in the immune function of juvenile cobia, Rachycentron canadum has been recorded after dietary supplementation of vitamin C (Hartel et al., 2007; Zhou et al., 2012). Vitamin C has also got significant impact as an immunomodulator in case of infection caused by Pasteurella monodon (Kanagu et al., 2010).
Induction of vitamin B complex deficiency in pregnant rats resulted in defect in the formation of lymphoid organs especially thymus and spleen in their progeny. The study indicates a role of vitamin B in the development of these vital organs and thus on the level of circulating lymphocyte (Dhur et al., 1991). Further, vitamin B complex deficiency leads to reduced number of plasma cells; suppressed lymphocyte proliferation and diminished CT cell activity etc., resulting in lowered DTH response (Fawzi et al., 1999; Liu et al., 2011b). Vitamin B6 is essentially required as a cofactor in the biosynthesis of nucleic acids and proteins and hence its role in immune function can be appreciated, since most of the immune key performers like antibodies and cytokines built up from these biomolecules (Leklem, 2001). The deficiency of vitamin B6 favors the Th2 mediated immunity (Long and Santos, 1999) and thus affects cellular immunity. Deficiency of biotinase may lead to cellular immunity abnormalities in mice (Pindolia et al., 2014). Similar to Vitamin B6, folate also plays a significant role in protein and nucleic acid synthesis and therefore folate deficiency significantly impairs the immune response. Folate deficiency also alters immune competence and protection against microbes and impairs cell-mediated immunity by decreasing the proportion of circulating T lymphocytes and their ftlinemultiplication in response to mitogen activation (Dhur et al., 1991). Folate, moreover, increases biochemical constituents and enhances total IgG generation; in poultry it exhibits pleiotropic effects in inflammatory responses (Munyaka et al., 2012). In Helicobacter-associated gastric cancer in mice, folic acid supplementation can be chemo-preventive as it prevents global loss of methylation and suppresses inflammation (Gonda et al., 2012). Vitamin B12 may have an immunomodulatory influence on cellular immunity (Tamura et al., 1999). Vitamin B12 deficiency may influence the purine and thymidine synthesis and ultimately RNA and DNA synthesis that may lead to alterations in immunological secretions (Bailey and Gregory, 2006).
Trace elements and immune function: Essentially trace elements are nutrients which are required in small quantities for supporting an organism’s cellular functions (McClung and Peterson, 2010). The immune system is a sensitive indicator of the nutritional state with regard to trace elements (Girish, 2011). The list of micronutrients that have significant influence on immune function is ever increasing. It essentially includes zinc, copper, iron, selenium etc. Deficiency of the trace elements would increase one’s susceptibility to various infections (Goswami et al., 2005).
Selenium is crucial for maximal immune response and modulates both arms of immunity as it is a potent nutritional antioxidant. Selenium is required as a trace mineral even in individuals who are not suffering from overt deficiency of the mineral (Moeini et al., 2011; Zhang et al., 2012; Safaralizadeh et al., 2013). It carries out biological effects through Selenium-dependent enzyme or selenoproteins (Hoffmann and Berry, 2008) and glutathione peroxidase (Chaudhary et al., 2010). It further protects cellular membranes and organelles from peroxidative damage (Mahima et al., 2012b) and oxidative stress generated by the microbial effect of macrophages that could contribute to its anti-inflammatory effects (Huang et al., 2012). It plays an important role in balancing the redox state and help to protect the host from oxidative stress through glutathione peroxidases. The thioredoxin reductase which is a selenoenzyme, impairs the redox regulation of various key enzymes, transcription receptors and factors, including glucocorticoid receptors, ribonucleotide reductase, nuclear factor-kappa B (NFκB) and anti-inflammatory protein AP-1 which binds to DNA and potentiates the expression of genes responsible for coding of proteins involved in immune response such as cytokines and adhesion molecules (Wintergerst et al., 2007; Arthur et al., 2003; Ryan-Harshman and Aldoori, 2005). Selenium in addition also promotes TR1 expression and enhances antioxidative capacity in porcine splenocytes causing TCR or ConA-induced T-cell activation enhancement (Ren et al., 2012). Deficiency of Selenium causes poor development of immune organs due to oxidative stress (Zhang et al., 2012). It ultimately influences the innate and non-adaptive; acquired and adaptive immune responses (Beckett et al., 2003; Carlson et al., 2010).
Zinc is a trace mineral that is required as a cofactor for the activities of more than 300 enzymes besides its involvement in various metabolic processes (Cuevas and Koyanagi, 2005). Zinc is crucial for highly multiplying cells of the immune system and potentiates both arms of the immunity. Its superoxide dismutase activity results in protective action against oxidative stress and it is a crucial cofactor for thymulin production which regulates cytokine secretion and enhances proliferation. Sufficient amount of zinc intake favors the Th1 type immunity and enables to manage skin and mucosal membrane integrity (Wintergerst et al., 2006; Prasad, 2000; Fraker and King, 2004; Haase and Rink, 2009).
Copper plays a significant role in the formation and maintenance of the immune system and a very small quantity of it is required for individual’s well being (Josko, 2011). Catalase and glutathione peroxidase are important cytosolic antioxidant agents and have synergistic defense activity against ROS. Copper is necessary for the dismutation of superoxide anion into H2O2 and oxygen. Moreover, it minimizes damage to lipids as well as proteins and DNA. Both deficiency and chronic excessive intakes of copper can alter various components of the immune response (Minatel and Carfagnini, 2000; Bonham et al., 2002; Klotz et al., 2003). Copper deficiency leads to decrease in thymus weight and ceruloplasmin activity; T-helper cells and immunoglobulins; responsiveness of lymphocytes to mitogens (T and B-cell mitogens); antibody response with increased susceptibility to infection and cytokine production (Solaiman et al., 2007). Natural Killer (NK) cells and antimicrobial activity of phagocytes are also decreased in copper deficiency (Klurfeld, 1993).
Iron plays a crucial role in gene regulation and electron transfer reactions and is an important component of nitric oxide and peroxide producing enzymes. Iron is also important in the control of cytokine production. It activates protein kinase C which is necessary for phosphorylation of factors responsible for regulating cell proliferation. Furthermore, iron is essential for myeloperoxidase activity which is responsible for the killing of bacteria by neutrophils via the generation of highly toxic hydroxyl radicals (Food and Nutrition Board, 2001; Weiss, 2004). Any disturbance in the homeostasis of cellular iron such as either deficiency or excess has detrimental effects on the immune system. Because pathogens need iron and other micronutrients for their replication and survival, it is essential to prevent the access of iron to the infecting microorganism. It is also necessary to maintain an appropriate concentration of iron so that the host can maintain optimal immune response and avoid the overload of iron which may cause free radical mediated injury to the host cells (Openheimer, 2001; Schaible and Kaufmann, 2004; Weiss, 2002). Iron deficiency leads to anemia making individuals susceptible to infection. Iron deficiency also causes lymphopenia due to decrease in IgM-positive lymphocyte counts (in piglets); impairment or delay in the development of cell mediated immunity; decrease in myeloperoxidase and phagocytic activity of neutrophils. Eosinophil counts and immunoglobulin concentration become low; there may be delay in cutaneous hypersensitivity and Natural Killer (NK) cell activity (Svoboda et al., 2007; Nairz et al., 2010; Cherayil, 2011).
Iodine acts with the myeloperoxidase in vitro in the phagocytic cells to destroy the bacteria; increases production of IgG by the humans B lymphocytes. Iodine has antioxidant activity in stomach, breast and thyroid and immunodeficiency results in increased risk for cancers (Venturi et al., 2000). Overload of iodine is responsible for hypo or hyperthyroidism characterized by impairment of natural killer cell activity (Wenzel et al., 1998). Iodine acts as an inducing factor in autoimmunity of thyroid gland as with the increase in iodine concentration in rats, the antigen presentation activity of macrophages is increased (Zhao et al., 2008).
Polysaccharides: Polysaccharides and oligosaccharides from various plants mimic pathogen associated molecular patterns (PAMPs) and bind to Toll Like Receptor (TLR) so these modulates immunity. TLRs are thus gaining importance as targets to modulate immunity (Liu et al., 2011a). Polysaccharides namely β-D-glucans and α-D-mannans are the two components found in the cell wall of yeast. Yeast extracts are mostly used in poultry and swine industry as growth promoters. These two polysaccharides act on macrophages and other immune cells thereby modulating the immunity of these animals (Tzianabos, 2000). Other polysaccharides from different sources can also have similar immunomodulatory functions. Anti inflammatory properties are possessed by one of the polysaccharide component present in the yeast which is β-D-glucan. Yeast fed to pigs and challenged with lipopolysaccharide (LPS) of bacterial origin has shown that β-D-glucans has reduced the production of pro inflammatory cytokines and increased the production of anti inflammatory cytokines (Li et al., 2005, 2006). Due to the reduction in proinflammatory cytokines like IL 1, IL 6 and Tumor necrosis factor α (TNF α) it controls the release of acute phase proteins (Baumann and Gauldie, 1994). β-glucans can bind with many receptors of the immune system which include scavenging receptors, complement receptor 3, TLR 4 and also TLR 7 (Tsoni and Brown, 2008). β-glucans from Candida albicans when administered to laboratory animal model showed increase in neutrophils, macrophages, eosinophils and CD80+ and CD86+ activated leukocytes in the alveolar tissues (Suzuki et al., 2001). Studies in pigs with β-glucans showed that there is reduction in the heptaglobulin content of blood which usually increases post weaning in piglets. There is increase of both pro-and anti-inflammatory response in pigs that are challenged with LPS after giving β-glucans (Eicher et al., 2006). These β-glucans also have different activity against different class of antibodies in a dose dependent manner. At higher concentration they suppress IgG class of antibodies and at lower concentration they increase IgA class antibody release. β-glucans at times act on different pathways in the cells like NF-kappa B pathway and this is dependent on the concentration of the glucan.
Another polysaccharide mannan has the fascinating property to adsorb the pathogens in the intestinal tract there by protecting the animals from infection (Sohn et al., 2000). Mannan fed through diet to animals has shown increased phagocytic activity of macrophages especially in the lamina propria of the intestine (Davis et al., 2004). Systemically there is a shift towards B cells than T cells after animals are fed with mannans (Davis et al., 2004). There is no alteration in the proliferation of blood lymphocytes in vitro after administrating mannan. Mannan from C. albicans possesses immune suppressive activities which are evident after administering this agent. Mannan of C. albicans also raises pro inflammatory cytokines like TNFα, IL 1 and IL 6 (Garner et al., 1990). This increase of pro inflammatory cytokines is in the control of the dose of the mannan being administered and this increase can be blocked by using antibodies specific to the mannan. This phenomenon also reflects the pathogenesis of C. albicans (Garner and Hudson, 1996). Similarly, mannan from Cryptococcus neoformans has immune suppressive action which is regulated by T suppressor cells (Blackstock and Casadevall, 1997).
HELMINTHS
During helminth infection, there would be a mutual interaction between the helminth and host immune system which favors survival of both the host and the parasite (Gause et al., 2013). In this process helminth can control the development of autoimmune disease in the host (Maizels et al., 2009; Ashour, 2013; Shor et al., 2013; Dalton et al., 2013; Osada et al., 2013). There has been an increased demand for the development of novel therapy for treating as well as preventing a range of debilitating and life threatening conditions. Identification of parasite mediated key pathways involved in regulation of immune system is helpful for further development of novel therapies (Zaccone and Cooke, 2013). Th2 events which are evoked by helminths have been found to improve Th1 mediated immune disorders (El-Malky et al., 2011). Nutman and Kumaraswami (2001) reported that during filarial infection there will be decreased multiplication of lymphocytes which leads to reduced secretion of IFN-γ and enhanced production of IL-4, IL-10 and antibody isotype IgG4. As Th2 cells secretes important cytokine IL-4 and anti-inflammatory cytokine IL-10, so an immune response is predominantly of Th2 type. Thus autoimmune diseases considered as Th1 mediated disease would not be able to flourish with helminth infection in the same host. Host Defense Peptides (HDPs) are produced by various helminth worms such as Clonorchisin , Fasciola, Opisthorcis, Paragonimus and Schistosoma species have potential immunomodulating property through action of molecular mimicry resulting in anti-inflammatory effect against helminth diseases (Cotton et al., 2012). Infections of schistosomes and filariae have immunomodulatory capacity that is activated by different stages of life-cycle of parasites (Hubner et al., 2013). Heligmosomoides polygyrus, a gastrointestinal nematode prevents the decrease in pancreatic islet cells via Th2 polarization-independent mechanisms (Osada et al., 2013). Helminths exhibit the anti-inflammatory activities in various diseases like rheumatoid arthritis, type-1 diabetes, inflammatory bowel disease etc., through inhibition of IL-17 and IFN-γ secretion, promotion of IL-4, IL-10 and TGF-β production, induction of FoxP3(+) expression, CD4(+) T cell and generation of regulatory macrophages, dendritic cells and B cells (Shor et al., 2013). Trichinella spiralis can have the immunomodulatory activity through Th1/Th2 phenotype with predominance of Th2 response, macrophage activation, regulation of B cells (Ashour, 2013). By establishing the mechanism in which helminths regulate the immune response and inhibit autoimmune disease, it is feasible to formulate modern therapeutic options that may not need infection with live worms. To achieve this, molecules are prepared that mimic the activity of modulators derived from helminths to develop worm based drugs (Reddy, 2010).
| Table 12: | Vitamins and trace minerals as immunomodulator for livestock health |
 | |
Helminthic parasites also help to prevent allergies like asthma. Ascaris suum adult worm antigen and products of Toxascaris leonina prevents allergy during challenge studies (McSorley et al.,2012). Litomosoides sigmodontis and Schistosoma mansoni can prevent auto immune disorders (McSorley et al., 2013). Trichuris suis can be used in treatment of multiple sclerosis. Parasitic products suppress Th1 response while induce Th2 response thereby reduce inflammation (Fleming, 2011). Based on this concept various therapeutic trials have been conducted which are summarized in Table 12.
NON-SPECIFIC IMMUNOMODULATION BY VACCINES
Apart from all the above mentioned immunomodulators, researches have proved that vaccination of an individual against some infectious diseases could increase the host defense to counter other infectious agent. For example, tuberculosis and measles vaccine are linked with decrease in overall child mortality. The probable reason of this phenomenon might be the cross-reactivity of host immune system with some other infectious agent (Benn et al., 2013). Vaccination against vaccinia virus showed para-immune effects such as more than 40% reduction in adult mortality in low income countries (Aaby et al., 2006), whereas it reduces risk of asthma, malignant melanoma and infectious disease hospitalizations in high income countries (Pfahlberg et al., 2002; Sorup et al., 2011). The cancer vaccination or immunotherapy enhances the host body’s immune system nonspecifically and provides innate resistance to cancer. This method of immunomodulation in cancer therapies offers specific cytotoxicity to cancer cells and negligible cytotoxicity to the rest of the cells of the body (Kuhn and Hanke, 1997). It has been found in recent years that certain vaccines (viz., sipuleucel-T against prostate cancer in advanced stage) that provides non-specific immunity against cancer have got approval in the market (Winter et al., 2011). One of the most commonly administered vaccines against tuberculosis (TB) is Bacille Calmette-Guerin (BCG). Apart from TB, it can also reduce overall mortality of children which is attributed to non-specific effects (Ritz et al., 2013). Vaccine against Streptococcus pneumoniae also provides non-specific immunity (Flasche et al., 2013). Protection against orthopox diseases can also be provided non-specifically by vaccinia based vaccines (Rice et al., 2014).
Monoclonal antibodies can also have immunomodulation. Muromonab-CD3, a monoclonal antibody targeted against CD 3, led to decrease of T lymphocyte population in the blood circulation, this helps to prevent graft rejection (Patil et al., 2012).
PANCHGAVYA (COW THERAPY)
Cow therapy or cowpathy is made up of five constituents derived from Indian Zebu cow (panchagavya), namely milk, curd, ghee, urine and dung extract. Panchagavya has significant role in Ayurveda system of medicine and it has been mentioned in ancient Indian literature that the panchagavya increases body’s resistance and makes body refractory to infections (Dhama et al., 2005; Mathivanan et al., 2006a, b; Joseph and Sankarganesh, 2011; Mathivanan and Edwin, 2012). By enhancement of both cellular as well as humoral immune responses cowpathy upregulates the lymphocyte proliferation activity; secretion of cytokines and macrophage activity; decreases apoptosis of lymphocytes thereby helping them to survive and fight infection; acts as anti-aging factor by preventing formation of the free radicals and efficiently repairing the damaged DNA (Dhama et al., 2005). Ameliorative effect on certain viral diseases (e.g., New Castle disease in layer chicken) is an additional benefit of panchgavya (Sumithra et al., 2013). Recently, cow urine has been patented (U.S. Patents No. 6896907 and 6,410,059) for its medicinal properties especially fight against cancers and in controlling bacterial infection (Randhawa, 2010). Cow urine was found to potentiate the humoral and cell mediated immune response in mice, to increase B and T lymphocyte blastogenesis and to increase IgG and IgA antibody titers in mice. Based on these findings a preparation called Kamadhenu Ark was manufactured by Govigyan Anusandhan Kendra, Nagpur from the urine of Indian cows and was tested for its immunomodulatory properties in mice. There was an increase in T and B lymphocyte blastogenesis and IgG, IgM and IgA antibody titers in mice. The therapy also protected the DNA from oxidative damage, responsible for ageing, cancer etc. Cow ghee is traditionally believed to improve resistance of body to infections. It possesses immunostimulant activity that has been indicated by increase in neutrophil adhesion as well as haemagglutination (HA) titre and delayed type hypersensitivity (DTH) responses in rats (Fulzele et al., 2001). Both Panchagavya and Ark can help to reduce multiplication of the viruses such as the one causing swine flu (Chauhan et al., 2001; Dhama et al., 2013b). Anticonvulsant as well as hypoglycemic effects are exerted by Cow Urine Concoction (CUC) and has been found to be useful against liver disorders as well as fever; inflammation and anemia (Dhama et al., 2013b). Gauloka Peya which translates as ‘drink from the land of cow’ is another beverage prepared by incorporation of distilled and sterile cow urine with water and traditional Indian herbs and medicinal plants viz., Brahmi and basil (1: 7 ratio) and thus has medicinal properties (Rahman, 2010; Kulkarni, 2009; Ganguly and Prasad, 2011).
CONCLUSION
Nowadays, in natural health industry, immunomodulators are becoming very popular due to realization of people about the significance of a healthy immune system in the preservation of health as well as prevention and recovery from diseases. The issue of helping the body to help itself by optimizing the immune system is of vital significance. In this context immunomodulation holds great value in veterinary and medical sciences and needs to be explored extensively for the development of compounds that can increase the immunity in rapid changing environmental scenario. An ideal immunomodulator is yet to be discovered, developed and validated. Depending upon the situation, an immunomodulator can be used either to stimulate the immunity or to suppress the immunity to help the individual patient to maintain homeostasis. Lots of invalidated natural compounds, claimed to possess immunostimulating effects, circulate in the market that may mislead many physicians about the action and use of these compounds. Many of them are put out by companies for mere financial gain, it is therefore essential for a physician to have a thorough knowledge of their effects and side effects and idea regarding the techniques involved in comparing them. In the context of the affected individual’s immune response a good understanding of pathogenesis of microbes and the relative need for immune modulation and immune activation is required. This will be essential for the future use of adjunctive immunomodulators for infectious diseases. Moreover, it needs utmost care while testing the immunomodulatory properties of herbs or the materials from cow for their stability, effectiveness, duration of action and effect of temperature or moisture and pesticide and heavy metal residues present in them. Efforts should also be directed towards patenting effective plant products or preparations so that we can utilize our rich resources for the benefit of mankind and animal welfare. Due emphasis need to be given for exploring fully the mechanisms and modes of actions of potent and novel immunomodulatory molecules like cytokines, hormones, host defense peptides, TLRs, probiotics, nutrients, herbs, panchgavya, polysaccharides and others which would help in developing effective immunoadjuvants and prophylactics, boosting vaccinal and general immunity, enhancement of disease resistance ability and safeguarding health of humans and their companion animals from various diseases, disorders and stressful conditions. Strengthening of the field of immunomodulatory substances could equip the health industries with the powerful pharmaceuticals, biologicals and medicines for nurturing a healthier future in the era of one world, one health and one medicine concept.
ACKNOWLEDGMENT
All the authors of the manuscript thank and acknowledge their respective Universities and Institutes.
REFERENCES
- Aaby, P., P. Gustafson, A. Roth, A. Rodrigues and M. Fernandes et al., 2006. Vaccinia scars associated with better survival for adults: An observational study from Guinea-Bissau. Vaccine, 24: 5718-5725.
CrossRefDirect Link - AbdEIslam, N.M., S. Ahmad, R. Ullah, A. Bibi, I. Ahmad, M.S. Mostafa and J.A. Khader, 2013. Antibacterial assay of Withania somnifera. J. Pure Applied Microbiol., 7: 791-794.
Direct Link - Adorini, L., 2003. Cytokine-based immunointervention in the treatment of autoimmune diseases. Clin. Exp. Immunol., 132: 185-192.
CrossRefDirect Link - Agarwal, S.S. and V.K. Singh, 1999. Immunomodulators: A review of studies on Indian medicinal plants and synthetic peptides. Part 1: Medicinal plants. Proc. Indian Natl. Sci. Acad. Part B: Biol. Sci., B65: 179-204.
Direct Link - Agger, E.M., I. Rosenkrands, J. Hansen, K. Brahimi and B.S. Vandahl et al., 2008. Cationic liposomes formulated with synthetic mycobacterial cordfactor (CAF01): A versatile adjuvant for vaccines with different immunological requirements. PloS ONE, Vol. 3.
CrossRefDirect Link - Aher, V.D. and A. Wahi, 2010. Pharmacological study of Tinospora cordifolia as an immunomodulator. Int. J. Curr. Pharmaceut. Res., 2: 52-54.
Direct Link - Akira, S., 2011. Innate immunity and adjuvants. Philos. Trans. R. Soc. Lond. B: Biol. Sci., 366: 2748-2755.
CrossRefDirect Link - Alluwaimi, A.M., 2004. The cytokines of bovine mammary gland: Prospects for diagnosis and therapy. Res. Vet. Sci., 77: 211-222.
CrossRefPubMedDirect Link - Anderson, K.C., 2005. Lenalidomide and thalidomide: Mechanisms of action-similarities and differences. Semin. Hematol., 42: S3-S8.
CrossRefDirect Link - Armstrong-James, D., I.A. Teo, S. Shrivastava, M.A. Petrou, D. Taube, A. Dorling and S. Shaunak, 2010. Exogenous interferon-γ immunotherapy for invasive fungal infections in kidney transplant patients. Am. J. Transplantation, 10: 1796-1803.
CrossRefDirect Link - Antachopoulos, C. and E. Roilides, 2005. Cytokines and fungal infections. Br. J. Haematol., 129: 583-596.
CrossRefDirect Link - Arthur, J.R., R.C. McKenzie and G.J. Beckett, 2003. Selenium in the immune system. J. Nutr., 133: 1457S-1459S.
CrossRefDirect Link - Ashour, D.S., 2013. Trichinella spiralis immunomodulation: An interactive multifactorial process. Expert Rev. Clin. Immunol., 9: 669-675.
CrossRefDirect Link - Asnis, G.M. and G.R. de la Garza, 2006. Interferon-induced depression in chronic hepatitis C: A review of its prevalence, risk factors, biology and treatment approaches. J. Clin. Gastroenterol., 40: 322-335.
Direct Link - Aukrust, P., F. Mueller, T. Ueland, A. Svardal, R. Berge and S.S. Froland, 2000. Decreased vitamin A levels in common variable immunodeficiency: Vitamin A supplementation in vivo enhances immunoglobulin production and downregulates inflammatory responses. Eur. J. Clin. Invest., 30: 252-259.
PubMed - Awodele, O., S.O. Olayemi, T.A. Adeyemo, T.A. Sanya and D.C. Dolapo, 2012. Use of complementary medicine amongst patients on antiretroviral drugs in an HIV treatment centre in Lagos, Nigeria. Curr. Drug Saf., 7: 120-125.
CrossRefPubMedDirect Link - Baggiolini, M., A. Walz and S.L. Kunkel, 1989. Neutrophil-activating peptide-1/interleukin 8, a novel cytokine that activates neutrophils. J. Clin. Invest., 84: 1045-1049.
CrossRefPubMedDirect Link - Baidas, S.M., E.P. Winer, G.F. Fleming, L. Harris and J.M. Pluda et al., 2000. Phase II evaluation of thalidomide in patients with metastatic breast cancer. J. Clin. Oncol., 18: 2710-2717.
Direct Link - Balevi, T., U.S. Ucan, B. Cokun, V. Kurtolu and I.S. Cetingul, 2001. Effect of dietary probiotic on performance and humoral immune response in layer hens. Br. Poult. Sci., 42: 456-461.
CrossRefPubMedDirect Link - Bangham, A.D. and R.W. Horne, 1964. Negative staining of phospholipids and their structural modification by Surface-active agents as observed in the electron microscope. J. Mol. Biol., 8: 660-668.
CrossRefDirect Link - Baumann, H. and J. Gauldie, 1994. The acute phase response. Immunol. Today, 15: 74-80.
CrossRefPubMedDirect Link - Becker, A.M., A. Janik, E.K. Smith, C.A. Sousa and B.A. Peters, 1989. Propionibacterium acnes immunotherapy in chronic recurrent canine pyoderma: An adjunct to antibiotic therapy. J. Vet. Internal Med., 3: 26-30.
CrossRefDirect Link - Bedford, M., 2000. Removal of antibiotic growth promoters from poultry diets: Implications and strategies to minimise subsequent problems. World's Poult. Sci. J., 56: 347-365.
CrossRefDirect Link - Behnsen, J., E. Deriu, M. Sassone-Corsi and M. Raffatellu, 2013. Probiotics: Properties, examples and specific applications. Cold Spring Harbor Perspect. Med., Vol. 3.
CrossRef - Shor, D.B.A., M. Harel, R. Eliakim and Y. Shoenfeld, 2013. The hygiene theory harnessing helminths and their ova to treat autoimmunity. Clin. Rev. Allergy Immunol., 45: 211-216.
CrossRefDirect Link - Ben-Efraim, S., Y. Burstein, Y. Keisari, R. Ophir, M. Pecht and N. Trainin, 1999. Immunopotentiating and immunotherapeutic effects of thymic hormones and factors with special emphasis on thymic humoral factor THF-γ2. Crit. Rev. Immunol., 19: 261-284.
Direct Link - Benn, C.S., M.G. Netea, L.K. Selin and P. Aaby, 2013. A small jab-a big effect: Nonspecific immunomodulation by vaccines. Trends Immunol., 34: 431-439.
CrossRefDirect Link - Tan, B.K.H. and J. Vanitha, 2004. Immunomodulatory and antimicrobial effects of some traditional Chinese medicinal herbs: A review. Curr. Med. Chem., 11: 1423-1430.
CrossRefPubMedDirect Link - Benveniste, E.N., 1998. Cytokine actions in the central nervous system. Cytokine Growth Factor Rev., 9: 259-275.
CrossRefDirect Link - Bessler, W.G., 1992. Synthetic lipopeptide immunomodulators derived from bacterial lipoprotein: Tools for the standardization of in vitro assays. Dev. Biol. Stand., 77: 49-56.
PubMed - Bessler, W.G., P. Beck, U. Konetznick, M. Loleit and E. Sedelmeier et al., 1991. Biological activity of bacterial surface components. Immunogenicity and immunomodulatory properties of a bacterial extract from Escherichia coli. Arzneimittelforschung., 41: 274-279.
PubMed - Beynon, A.L., M.R. Brown, R. Wright, M.I. Rees, I.M. Sheldon and J.S. Davies, 2013. Ghrelin inhibits LPS-induced release of IL-6 from mouse dopaminergic neurones. J. Neuroinflammation, Vol. 10.
CrossRefDirect Link - Bhatt, P., S.K. Shukla, M.Y. Wani, R. Tiwari and K. Dhama, 2013. Amelioration of chicken infectious anaemia virus induced immunosuppression by immunomodulator and haematinic supplementation in chicks. Veterinarski Arhiv, 83: 639-652.
Direct Link - Bhowmik, D., Chiranjib, J. Yadav, K.K. Tripathi and K.P.S. Kumar, 2010. Herbal remedies of Azadirachta indica and its medicinal application. J. Chem. Pharm. Res., 2: 62-72.
Direct Link - Blackstock, R. and A. Casadevall, 1997. Presentation of cryptococcal capsular polysaccharide (GXM) on activated antigen-presenting cells inhibits the T-suppressor response and enhances delayed-type hypersensitivity and survival. Immunology, 92: 334-339.
CrossRefDirect Link - Blalock, J.E., 1989. A molecular basis for bidirectional communication between the immune and neuroendocrine systems. Physiol. Rev., 69: 1-32.
PubMed - Blodgett, D.J., G.G. Schuring and E.T. Kornegay, 1986. Immunomodulation in weanling swine with dietary selenium. Am. J. Vet. Res., 47: 1517-1519.
PubMed - Bohannon, J.K., A. Hernandez, P. Enkhbaatar, W.L. Adams and E.R. Sherwood, 2013. The immunobiology of Toll-like receptor 4 agonists: From endotoxin tolerance to immunoadjuvants. Shock, 40: 451-462.
CrossRefDirect Link - Bommineni, Y.R., H. Dai, Y.X. Gong, J.L. Soulages and S.C. Fernando et al., 2007. Fowlicidin-3 is an α-helical cationic host defense peptide with potent antibacterial and lipopolysaccharide-neutralizing activities. FEBS J., 274: 418-428.
CrossRefDirect Link - Bommineni, Y.R., G.H. Pham, L.T. Sunkara, M. Achanta and G. Zhang, 2014. Immune regulatory activities of fowlicidin-1, a cathelicidin host defense peptide. Mol. Immunol., 59: 55-63.
CrossRefPubMedDirect Link - Bonham, M., J.M. O'Connor, B.M. Hannigan and J.J. Strain, 2002. The immune system as a physiological indicator of marginal copper status? Br. J. Nutr., 87: 393-403.
CrossRefDirect Link - Borsuk, O.S., N.V. Masnaya, E.Y. Sherstoboev, N.V. Isaykina, G.I. Kalinkina and D.V. Reihart, 2011. Effects of drugs of plant origin on the development of the immune response. Bull. Exp. Biol. Med., 151: 194-196.
CrossRefDirect Link - Brocklehurst, P., M. Tickle, A.M. Glenny, M.A. Lewis and M.N. Pemberton et al., 2012. Systemic interventions for recurrent aphthous stomatitis (mouth ulcers). Cochrane Database Syst. Rev., Vol. 2012.
CrossRefDirect Link - Brooks, V.J., M. Schafer, P. Sharp, J. Xu and J. Cai et al., 2009. Effects of Morinda citrifolia (noni) on CD4+ and CD8+ T-cell activation in neonatal calves. Prof. Anim. Scientist, 25: 262-265.
Direct Link - Buts, J.P., P. Bernasconi, J.P. Vaerman and C. Dive, 1990. Stimulation of secretory IgA and secretory component of immunoglobulins in small intestine of rats treated with Saccharomyces boulardii. Dig. Dis. Sci., 35: 251-256.
CrossRefDirect Link - Calder, P.C., 2001. Polyunsaturated fatty acids, inflammation and immunity. Lipids, 36: 1007-1024.
PubMedDirect Link - Cannell, J.J., R. Vieth, J.C. Umhau, M.F. Holick and W.B. Grant et al., 2006. Epidemic influenza and vitamin D. Epidemiol. Infect., 134: 1129-1140.
CrossRefPubMedDirect Link - Cardones, A.R.G. and R.P. Hall, 2012. Management of dermatitis herpetiformis. Immunol. Allergy Clin. North Am., 32: 275-281.
CrossRefDirect Link - Carlson, B.A., M.H. Yoo, R.K. Shrimali, R. Irons, V.N. Gladyshev, D.L. Hatfield and J.M. Park, 2010. Role of selenium-containing proteins in T-cell and macrophage function. Proc. Nutr. Soc., 69: 300-310.
CrossRefDirect Link - Carrio, E., M. Rigat, T. Garnatje, M. Mayans, M. Parada and J. Valles, 2012. Plant ethnoveterinary practices in two pyrenean territories of Catalonia (Iberian Peninsula) and in two areas of the Balearic Islands and comparison with ethno-botanical uses in human medicine. Evidence-Based Complem. Altern. Med.
CrossRefDirect Link - Cassani, B., E.J. Villablanca, J. de Calisto, S. Wang and J.R. Mora, 2012. Vitamin A and immune regulation: Role of retinoic acid in gut-associated dendritic cell education, immune protection and tolerance. Mol. Aspects Med., 33: 63-76.
CrossRefDirect Link - Chakraborty, S. and S.K. Pal, 2012. Plants for cattle health: A review of ethno-veterinary herbs in veterinary health care. Ann. Ayurvedic Med., 1: 144-152.
Direct Link - Chanan-Khan, A.A., A. Swaika, A. Paulus, S.K. Kumar and J.R. Mikhael et al., 2013. Pomalidomide: The new immunomodulatory agent for the treatment of multiple myeloma. Blood Cancer J., Vol. 3.
CrossRefDirect Link - Chandra, R.K., 2002. Nutrition and the immune system from birth to old age. Eur. J. Clin. Nutr., 56: S73-S76.
CrossRefDirect Link - Chang, C.H., L.A. Cohn, A.E. DeClue, H. Liu and C.R. Reinero, 2013. Oral glucocorticoids diminish the efficacy of allergen-specific immunotherapy in experimental feline asthma. Vet. J., 197: 268-272.
CrossRefDirect Link - Chauhan, R.S., B.P. Singh and L.K. Singhal, 2001. Immunomodulation with kamdhenu ark in mice. J. Immunol. Immunopathol., 3: 74-77.
Direct Link - Chauhan, R.S., 2010. Nutrition, immunity and livestock health. Indian Cow: Scient. Econ. J., 7: 2-13.
Direct Link - Chen, J., X. Wang and C. Liu, 2014. Anti-tumour effects of polysaccharides isolated from Artemisia annua L by inducing cell apoptosis and immunomodulatory anti-hepatoma effects of polysaccharides. Afr. J. Tradit. Complem. Altern. Med., 11: 15-22.
Direct Link - Chen, M.H., W.S. Li, Y.S. Lue, C.L. Chu and I. Pan et al., 2013. Clitocybe nuda activates dendritic cells and acts as a DNA vaccine adjuvant. Evidence-Based Complem. Altern. Med.
CrossRefDirect Link - Cheng, C., J.G. Gall, M. Nason, C.R. King and R.A. Koup et al., 2010. Differential specificity and immunogenicity of adenovirus type 5 neutralizing antibodies elicited by natural infection or immunization. J. Virol., 84: 630-638.
Direct Link - Cherayil, B.J., 2011. The role of iron in the immune response to bacterial infection. Immun. Res., 50: 1-9.
CrossRefDirect Link - Chicoine, M.R., M. Zahner, E.K. Won, R.R. Kalra, T. Kitamura, A. Perry and R. Higashikubo, 2007. The in vivo antitumoral effects of lipopolysaccharide against glioblastoma multiforme are mediated in part by Toll-like receptor 4. Neurosurgery, 60: 372-380.
PubMed - Chihara, G., Y. Maeda, J. Hamuro, T. Sasakia and F. Fukuoka, 1969. Inhibition of mouse sarcoma 180 by polysaccharides from Lentinus edodes (Berk.) sing. Nature, 222: 687-688.
PubMedDirect Link - Chithra, P., G.B. Sajithlal and G. Chandrakasan, 1998. Influence of Aloe vera on the glycosaminoglycans in the matrix of healing dermal wounds in rats. J. Ethnopharmacol., 59: 179-186.
CrossRefDirect Link - Clement, F., S.N. Pramod and Y.P. Venkatesh, 2010. Identity of the immunomodulatory proteins from garlic (Allium sativum) with the major garlic lectins or agglutinins. Int. Immunopharmacol., 10: 316-324.
CrossRefDirect Link - Coley, W.B., 1894. Treatment of inoperable malignant tumors with the toxines of erysipelas and the Bacillus prodigiosus. Am. J. Med. Sci., 108: 50-66.
Direct Link - Corcionivoschi, N., D. Drinceanu, I.M. Pop, D. Stack, L. Stef, C. Julean and B. Bourke, 2010. The effect of probiotics on animal health. Anim. Sci. Biotechnol., 43: 35-41.
Direct Link - Cotton, S., S. Donnelly, M.W. Robinson, J.P. Dalton and K. Thivierge, 2012. Defense peptides secreted by helminth pathogens: Antimicrobial and/or immunomodulator molecules? Front. Immunol., Vol. 3.
CrossRefDirect Link - Cox, J.C. and A.R. Coulter, 1997. Adjuvants-a classification and review of their modes of action. Vaccine, 15: 248-256.
CrossRefDirect Link - Cuevas, L.E. and A. Koyanagi, 2005. Zinc and infection: A review. Ann. Trop. Paediatr., 25: 149-160.
CrossRefDirect Link - Cunningham-Rundles, S., D.F. McNeeley and A. Moon, 2005. Mechanisms of nutrient modulation of the immune response. J. Allergy Clin. Immunol., 115: 1119-1128.
CrossRefDirect Link - Daley, M.J., P.A. Coyle, T.J. Williams, G. Furda, R. Dougherty and P.W. Hayes, 1991. Staphylococcus aureus mastitis: Pathogenesis and treatment with bovine interleukin-1 β and interleukin-2. J. Dairy Sci., 74: 4413-4424.
CrossRefPubMedDirect Link - Dalloul, R.A., H.S. Lillehoj, T.A. Shellem and J.A. Doerr, 2003. Intestinal immunomodulation by vitamin a deficiency and Lactobacillus-based probiotic in Eimeria acervulina-infected broiler chickens. Avian Dis., 47: 1313-1320.
PubMedDirect Link - Dalloul, R.A., H.S. Lillehoj, N.M. Tamim, T.A. Shellem and J.A. Doerr, 2005. Induction of local protective immunity to Eimeria acervulina by a Lactobacillus-based probiotic. Comp. Immunol. Microbiol. Infect. Dis., 28: 351-361.
CrossRefDirect Link - Dalton, J.P., M.W. Robinson, G. Mulcahy, S.M. O'Neill and S. Donnelly, 2013. Immunomodulatory molecules of Fasciola hepatica: Candidates for both vaccine and immunotherapeutic development. Vet. Parasitol., 195: 272-285.
CrossRefDirect Link - Dan, G. and S.B. Lall, 1998. Neuroendocrine modulation of immune system. Indian J. Phamacol., 30: 129-140.
Direct Link - Datta, K. and L. Pirofski, 2006. Towards a vaccine for Cryptococcus neoformans: Principles and caveats. FEM Yeast Res., 6: 525-536.
CrossRefDirect Link - Davis, R.H., J.J. Donato, G.M. Hartman and R.C. Haas, 1994. Anti-inflammatory and wound healing activity of a growth substance in Aloe vera. J. Am. Podiatr. Med. Assoc., 84: 77-81.
Direct Link - Davis, M.E., C.V. Maxwell, G.F. Erf, D.C. Brown and T.J. Wistuba, 2004. Dietary supplementation with phosphorylated mannans improves growth response and modulates immune function of weanling pigs. J. Anim. Sci., 82: 1882-1891.
PubMed - Dawson, H.D., N.Q. Li, K.L. DeCicco, J.A. Nibert and A.C. Ross, 1999. Chronic marginal vitamin A status reduces natural killer cell number and function in aging Lewis rats. J. Nutr., 129: 1510-1517.
Direct Link - Delsing, C.E., M.S. Gresnigt, J. Leentjens, F. Preijers and F.A. Frager et al., 2014. Interferon-gamma as adjunctive immunotherapy for invasive fungal infections: A case series. BMC Infect. Dis., Vol. 14.
CrossRefDirect Link - DePaolo, R.W., V. Abadie, F. Tang, H. Fehlner-Peach and J.A. Hall et al., 2011. Co-adjuvant effects of retinoic acid and IL-15 induce inflammatory immunity to dietary antigens. Nature, 471: 220-224.
CrossRefDirect Link - De Gregorio, E., E. Caproni and J.B. Ulmer, 2013. Vaccine adjuvants: Mode of action. Front. Immunol., Vol. 4.
CrossRefDirect Link - Deng, J., Y. Li, J. Zhang and Q. Yang, 2013. Co-administration of Bacillus subtilis RJGP16 and Lactobacillus salivarius B1 strongly enhances the intestinal mucosal immunity of piglets. Res. Vet. Sci., 94: 62-68.
CrossRefDirect Link - Deng, S., S. Zhu, Y. Qiao, Y.J. Liu, W. Chen, G. Zhao and J. Chen, 2014. Recent advances in the role of toll-like receptors and TLR agonists in immunotherapy for human glioma. Protein Cell, 5: 899-911.
CrossRefDirect Link - Dhama, K., M. Mahendran, S. Tomar and R.S. Chauhan, 2008. Beneficial effects of probiotics and prebiotics in livestock and poultry: The current perspectives. Intas Polivet, 9: 1-12.
Direct Link - Dhama, K., M. Mahendran, R.S. Chauhan and S. Tomar, 2008. Cytokines-Their functional roles and prospective applications in Veterinary practice: A review. J. Immunol. Immunopathol., 10: 79-89.
Direct Link - Dhama, K., V. Verma, P.M. Sawant, R. Tiwari, R.K. Vaid and R.S. Chauhan, 2011. Applications of probiotics in poultry: Enhancing immunity and beneficial effects on production performances and health: A review. J. Immunol. Immunopathol., 13: 1-19.
Direct Link - Dhama, K., S. Chakraborty, Mahima, M.Y. Wani and A.K. Verma et al., 2013. Novel and emerging therapies safeguarding health of humans and their companion animals: A review. Pak. J. Biol. Sci., 16: 101-111.
CrossRefDirect Link - Dhama, K., S. Chakraborty and R. Tiwari, 2013. Panchgavya therapy (Cowpathy) in safeguarding health of animals and humans: A review. Res. Opin. Anim. Vet. Sci., 3: 170-178.
Direct Link - Dhama, K., S. Chakraborty, M.Y. Wani, R. Tiwari and R. Barathidasan, 2013. Cytokine therapy for combating animal and human diseases-A review. Res. Opin. Anim. Vet. Sci., 3: 195-208.
Direct Link - Dhama, K., S. Chakraborty, S. Kapoor, R. Tiwari and A. Kumar et al., 2013. One world, one health-veterinary perspectives. Adv. Anim. Vet. Sci., 1: 5-13.
Direct Link - Dhama, K., S. Chakraborty, R. Tiwari, A.K. Verma and M. Saminathan et al., 2014. A concept paper on novel technologies boosting production and safeguarding health of humans and animals. Res. Opin. Anim. Vet. Sci., 4: 353-370.
Direct Link - Dhur, A., P. Galan and S. Hercberg, 1991. Folate status and the immune system. Prog. Food Nutr. Sci., 15: 43-60.
Direct Link - Dongarra, M.L., V. Rizzello, L. Muccio, W. Fries, A. Cascio, I. Bonaccorsi and G. Ferlazzo, 2013. Mucosal immunology and probiotics. Curr. Allergy Asthma Rep., 13: 19-26.
CrossRefDirect Link - Easton, D.M., A. Nijnik, M.L. Mayer and R.E. Hancock, 2009. Potential of immunomodulatory host defense peptides as novel anti-infectives. Trends Biotechnol., 27: 582-590.
CrossRefDirect Link - Duda, P.W., M.C. Schmied, S.L. Cook, J.I. Krieger and D.A. Hafler, 2000. Glatiramer acetate (Copaxone®) induces degenerate, Th2-polarized immune responses in patients with multiple sclerosis. J. Clin. Invest., 105: 967-976.
CrossRefDirect Link - Edelman, R., 2002. The development and use of vaccine adjuvants. Mol. Biotechnol., 21: 129-148.
CrossRefDirect Link - Eicher, S.D., C.A. McKee, J.A. Carroll and E.A. Pajor, 2006. Supplemental vitamin C and yeast cell wall β-glucan as growth enhancers in newborn pigs and as immunomodulators after an endotoxin challenge after weaning. J. Anim. Sci., 84: 2352-2360.
CrossRefPubMedDirect Link - Eisen, T., C. Boshoff, I. Mak, F. Sapunar and M.M. Vaughan et al., 2000. Continuous low dose thalidomide: A phase II study in advanced melanoma, renal cell, ovarian and breast cancer. Br. J. Cancer, 82: 812-817.
CrossRefDirect Link - Eisenbarth, S.C., O.R. Colegio, W. O'Connor, F.S. Sutterwala and R.A. Flavell, 2008. Crucial role for the Nalp3 inflammasome in the immunostimulatory properties of aluminium adjuvants. Nature, 453: 1122-1126.
CrossRefDirect Link - El Andaloussi, A., A.M. Sonabend, Y. Han and M.S. Lesniak, 2006. Stimulation of TLR9 with CpG ODN enhances apoptosis of glioma and prolongs the survival of mice with experimental brain tumors. Glia, 54: 526-535.
CrossRefDirect Link - Elkamel, A.A. and G.M. Mosaad, 2012. Immunomodulation of Nile Tilapia, Oreochromis niloticus, by Nigella sativa and Bacillus subtilis. J. Aquacult. Res. Dev., Vol. 3.
CrossRefDirect Link - El-Malky, M., N. Nabih, M. Heder, N. Saudy and M. El-Mahdy, 2011. Helminth infections: Therapeutic potential in autoimmune disorders. Parasite Immunol., 33: 589-593.
CrossRefDirect Link - Escudier, B., N. Lassau, D. Couanet, E. Angevin and F. Mesrati et al., 2002. Phase II trial of thalidomide in renal-cell carcinoma. Ann. Oncol., 13: 1029-1035.
PubMed - Eskandari, F., J.I. Webster and E.M. Sternberg, 2003. Neural immune pathways and their connection to inflammatory diseases. Arthritis Res. Ther., 5: 251-265.
Direct Link - Fawzi, W.W., R.L. Mbise, E. Hertzmark, M.R. Fataki, M.G. Herrera, G. Ndossi and D. Spiegelman, 1999. A randomized trial of vitamin A supplements in relation to mortality among human immunodeficiency virus-infected and uninfected children in Tanzania. Pediatr. Infect. Dis. J., 18: 127-133.
PubMed - Fernandes de Abreu, D.A., D. Eyles and F. Feron, 2009. Vitamin D, a neuro-immunomodulator: Implications for neurodegenerative and autoimmune diseases. Psychoneuroendocrinology, 34: S265-S277.
CrossRefDirect Link - Fife, K., R.H. Phillips, M.R. Howard, M. Bower and F. Gracie, 1998. Activity of thalidomide in AIDS-related Kaposi's sarcoma and correlation with HHV8 titre. Int. J. STD AIDS, 9: 751-755.
PubMed - Figg, W.D., W. Dahut, P. Duray, M. Hamilton and A. Tompkins et al., 2001. A randomized phase II trial of thalidomide, an angiogenesis inhibitor, in patients with androgen-independent prostate cancer. Clin. Cancer Res., 7: 1888-1893.
PubMed - Fimiani, V., A. Cavallaro, T. Ainis, G. Baranovskaia, O. Ketlinskaya and L. Kozhemyakin, 2002. Immunomodulatory effect of glutoxim on some activities of isolated human neutrophils and in whole blood. Immunopharmacol. Immunotoxicol., 24: 627-638.
Direct Link - Flasche, S., W.J. Edmunds, E. Miller, D. Goldblatt, C. Robertson and Y.H. Choi, 2013. The impact of specific and non-specific immunity on the ecology of Streptococcus pneumonia and the implications for vaccination. Proc. R. Soc. Biol. Sci., Vol. 280.
CrossRefDirect Link - Flatz, L., A.N. Hegazy, A. Bergthaler, A. Verschoor and C. Claus et al., 2010. Development of replication-defective lymphocytic choriomeningitis virus vectors for the induction of potent CD8+T cell immunity. Nat. Med., 16: 339-345.
CrossRefDirect Link - Fleming, J.O., 2011. Helminths and multiple sclerosis: Will old friends give us new treatments for MS? J. Neuroimmunol., 233: 3-5.
CrossRefPubMedDirect Link - Forton, D. and P. Karayiannis, 2006. Established and emerging therapies for the treatment of viral hepatitis. Dig. Dis., 24: 160-173.
CrossRefDirect Link - Fraile, L., E. Crisci, L. Cordoba, M.A. Navarro, J. Osada and M. Ontoya, 2012. Immunomodulatory properties of β-sitosterol in pig immune responses. Int. Immunopharmacol., 13: 316-321.
CrossRefDirect Link - Fraker, P.J. and L.E. King, 2004. Reprogramming of the immune system during zinc deficiency. Annu. Rev. Nutr., 24: 277-298.
CrossRefDirect Link - Franchimont, D., 2004. Overview of the actions of glucocorticoids on the immune response: A good model to characterize new pathways of immunosuppression for new treatment strategies. Ann. N. Y. Acad. Sci., 1024: 124-137.
CrossRefDirect Link - Franchimont, D., J. Galon, M. Gadina, R. Visconti and Y.J. Zhou et al., 2000. Inhibition of Th1 immune response by glucocorticoids: Dexamethasone selectively inhibits IL-12-induced Stat4 phosphorylation in T lymphocytes. J. Immunol., 164: 1768-1774.
PubMed - Freund, J. and K. McDermott, 1942. Sensitization to horse serum by means of adjuvants. Exp. Biol. Med., 49: 548-553.
Direct Link - Friis, T., A.M. Engel, B.M. Klein, J. Rygaard and G. Houen, 2005. Levamisole inhibits angiogenesis in vitro and tumor growth in vivo. Angiogenesis, 8: 25-34.
CrossRefDirect Link - Fuller, R., 2001. The chicken gut microflora and probiotic supplements. J. Poult. Sci., 38: 189-196.
CrossRefDirect Link - Gallois, M. and I.P. Oswald, 2008. Immunomodulatoes as efficient alternatives to in-feed antimicrobials in pig production. Archiva Zootechnica, 11: 15-32.
Direct Link - Ganguly, S. and A. Prasad, 2011. Role of plant extracts and cow urine distillate as immunomodulators: A review. J. Med. Plant Res., 5: 649-651.
Direct Link - Ganju, L., D. Karan, S. Chanda, K.K. Srivastava, R.C. Sawhney and W. Selvamurthy, 2003. Immunomodulatory effects of agents of plant origin. Biomed. Pharmacother., 57: 296-300.
CrossRefDirect Link - Garcia-Sanz, R., 2006. Thalidomide in multiple myeloma. Expert Opin. Pharmacother., 7: 195-213.
CrossRefDirect Link - Garner, R.E., A.M. Childress, L.G. Human and J.E. Domer, 1990. Characterization of Candida albicans mannan-induced, mannan-specific delayed hypersensitivity suppressor cells. Infect. Immun., 58: 2613-2620.
Direct Link - Garner, R.E. and J.A. Hudson, 1996. Intravenous injection of Candida-derived mannan results in elevated tumor necrosis factor alpha levels in serum. Infect. Immun., 64: 4561-4566.
PubMed - Gause, W.C., T.A. Wynn and J.E. Allen, 2013. Type 2 immunity and wound healing: Evolutionary refinement of adaptive immunity by helminths. Nat. Rev. Immunol., 13: 607-614.
CrossRefDirect Link - Georgakopoulou, E.A. and C. Scully, 2013. Systemic use of non-biologic agents in orofacial diseases: Other immunomodulatory agents. Oral Dis.
CrossRefDirect Link - German, A.J., V.H. Ryan, A.C. German, I.S. Wood and P. Trayhurn, 2010. Obesity, its associated disorders and the role of inflammatory adipokines in companion animals. Vet. J., 185: 4-9.
CrossRefPubMedDirect Link - Gonda, T.A., Y.I. Kim, M.C. Salas, M.V. Gamble and W. Shibata et al., 2012. Folic acid increases global DNA methylation and reduces inflammation to prevent Helicobacter-associated gastric cancer in mice. Gastroenterology, 142: 824-833.
CrossRefDirect Link - Gopi, M., K. Karthik, H.V. Manjunathachar, P. Tamilmahan and M. Kesavan et al., 2014. Essential oils as a feed additive in poultry nutrition. Adv. Anim. Vet. Sci., 2: 1-7.
CrossRefDirect Link - Goswami, T.K., R. Bhar, S.E. Jadhav, S.N. Joardar and G.C. Ram, 2005. Role of dietary zinc as a nutritional immunomodulator. Asian-Aust. J. Anim. Sci., 18: 439-452.
Direct Link - Greiner, L.L., T.S. Stahly and T.J. Stabel, 2001. The effect of dietary soy daidzein on pig growth and viral replication during a viral challenge. J. Anim. Sci., 79: 3113-3119.
PubMedDirect Link - Guo, N.L., D.P. Lu, G.L. Woods, E. Reed, G.Z. Zhou, L.B. Zhang and R.H. Waldman, 1993. Demonstration of the anti-viral activity of garlic extract against human cytomegalovirus in vitro. Chinese Med. J., 106: 93-96.
PubMedDirect Link - Haase, H. and L. Rink, 2009. The immune system and the impact of zinc during aging. Immunity Ageing, Vol. 6.
CrossRefDirect Link - Hartel, C., T. Strunk, P. Bucsky and C. Schultz, 2004. Effects of vitamin C on intracytoplasmic cytokine production in human whole blood monocytes and lymphocytes. Cytokine, 27: 101-106.
CrossRefDirect Link - Hagenlocher, Y. and A. Lorentz, 2015. Immunomodulation of mast cells by nutrients. Mol. Immunol., 63: 25-31.
CrossRefDirect Link - Haghighi, H.R., J. Gong, C.L. Gyles, M.A. Hayes and H. Zhou et al., 2006. Probiotics stimulate production of natural antibodies in chickens. Clin. Vaccine Immunol., 13: 975-980.
CrossRefDirect Link - Hajhashemi, V., A. Ghannadi and H. Jafarabadi, 2004. Black cumin seed essential oil, as a potent analgesic and antiinflammatory drug. Phytother. Res., 18: 195-199.
CrossRefDirect Link - Hamill, P., K. Brown, H. Jenssen and R.E.W. Hancock, 2008. Novel anti-infectives: Is host defence the answer? Curr. Opin. Biotechnol., 19: 628-636.
CrossRef - Hancock, R.E.W. and H.G. Sahl, 2006. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol., 24: 1551-1557.
CrossRefDirect Link - Hanel, P., P. Andreani and M.H. Graler, 2007. Erythrocytes store and release sphingosine 1-phosphate in blood. Federat. Am. Soc. Exp. Biol. J., 21: 1202-1209.
CrossRefDirect Link - Hartel, C., A. Puzik, W. Gopel, P. Temming, P. Bucsky and C. Schultz, 2007. Immunomodulatory effect of vitamin C on intracytoplasmic cytokine production in neonatal cord blood cells. Neonatology, 91: 54-60.
CrossRef - Hasani-Ranjbar, S., S. Khosravi, N. Nayebi, B. Larijani and M. Abdollahi, 2012. Systematic review of the efficacy and safety of anti-aging herbs in animals and human. Asian J. Anim. Vet. Adv., 7: 621-640.
CrossRefDirect Link - Hashemi, S.R. and H. Davoodi, 2012. Herbal plants as new immuno-stimulator in poultry industry: A review. Asian J. Anim. Vet. Adv., 7: 105-116.
CrossRefDirect Link - Hecker, M., J. Ott, C. Sondermann, M.B. Schaefer and M. Obert et al., 2014. Immunomodulation by fish-oil containing lipid emulsions in murine acute respiratory distress syndrome. Crit. Care, Vol. 18.
CrossRefDirect Link - Hegde, M., S.S. Karki, E. Thomas, S. Kumar and K. Panjamurthy et al., 2012. Novel levamisole derivative induces extrinsic pathway of apoptosis in cancer cells and inhibits tumor progression in mice. PLoS ONE, Vol. 7.
CrossRefDirect Link - Hirazumi, A. and E. Furusawa, 1999. An immunomodulatory polysaccharide‐rich substance from the fruit juice of Morinda citrifolia (Noni) with antitumour activity. Phytother. Res., 13: 380-387.
CrossRefPubMedDirect Link - Ho, N.K., S.P. Hawley, J.C. Ossa, O. Mathieu, T.A. Tompkins, K.C. Johnson-Henry and P.M. Sherman, 2013. Immune signalling responses in intestinal epithelial cells exposed to pathogenic Escherichia coli and lactic acid-producing probiotics. Benef. Microbes, 4: 195-209.
CrossRefDirect Link - Hoffmann, P.R. and M.J. Berry, 2008. The influence of selenium on immune responses. Mol. Nutr. Food Res., 52: 1273-1280.
CrossRefDirect Link - Hoorn, E.J., S.B. Walsh, J.A. McCormick, R. Zietse, R.J. Unwin and D.H. Ellison, 2012. Pathogenesis of calcineurin inhibitor-induced hypertension. J. Nephrol., 25: 269-275.
Direct Link - Hormannsperger, G., M.A. von Schillde and D. Haller, 2013. Lactocepin as a protective microbial structure in the context of IBD. Gut Microbes, 4: 152-157.
CrossRefDirect Link - Hsieh, P.S., Y. An, Y.C. Tsai, Y.C. Chen and C.J. Chuang et al., 2013. Potential of probiotic strains to modulate the inflammatory responses of epithelial and immune cells in vitro New Microbiol., 36: 167-179.
PubMed - Huang, K.Y., P.D. Kind, E.J. Jagoda and A.L. Goldstein, 1981. Thymosin treatment modulates production of interferons. J. Interferon Res., 1: 411-420.
CrossRefDirect Link - Huang, Z., A.H. Rose and P.R. Hoffmann, 2012. The role of selenium in inflammation and immunity: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signaling, 16: 705-743.
CrossRefDirect Link - Hubner, M.P., L.E. Layland and A. Hoerauf, 2013. Helminths and their implication in sepsis: A new branch of their immunomodulatory behaviour? Pathog. Dis., 69: 127-141.
CrossRefDirect Link - Huff, G.R., W.E. Huff and N.C. Rath, 2009. Nutritional immunomodulation as an approach to decreasing then effects of stress in poultry production. J. Arkansas Acad. Sci., 63: 87-92.
Direct Link - Hunter, H.N., D.B. Fulton, T. Ganz and H.J. Vogel, 2002. The solution structure of human hepcidin, a peptide hormone with antimicrobial activity that is involved in iron uptake and hereditary hemochromatosis. J. Biol. Chem., 277: 37597-37603.
CrossRefDirect Link - Hwang, S.A., M.L. Kruzel and J.K. Actor, 2014. Immunomodulatory effects of recombinant lactoferrin during MRSA infection. Int. Immunopharmacol., 20: 157-163.
CrossRefDirect Link - Ichikawa, H., Y. Takada, S. Shishodia, B. Jayaprakasam, M.G. Nair and B.B. Aggarwal, 2006. Withanolides potentiate apoptosis, inhibit invasion and abolish osteoclastogenesis through suppression of nuclear factor-κB (NF-κB) activation and NF-κB-regulated gene expression. Mol. Cancer Ther., 5: 1434-1445.
CrossRefPubMedDirect Link - Isaacs, A. and J. Lindenmann, 1957. Virus interference. I. The interferon. Proc. R. Soc. Lond., B, Biol. Sci., 147: 258-267.
CrossRefDirect Link - Isenberg, J., B. Stoffel, U. Wolters, J. Beuth, H. Stuetzer, H.L. Ko and H. Pichimaier, 1995. Immunostimulation by propionibacteria: Effects on immune status and antineoplastic treatment. Anticancer Res., 15: 2363-2368.
Direct Link - Isolauri, E., Y. Sutas, P. Kankaanpaa, H. Arvilommi and S. Salminen, 2001. Probiotics: Effects on immunity. Am. J. Clin. Nutr., 73: 444s-450s.
PubMedDirect Link - Jacob, R.A., D.S. Kelley, F.S. Pianalto, M.E. Swendseid and S.M. Henning et al., 1991. Immunocompetence and oxidant defense during ascorbate depletion of healthy men. Am. J. Clin. Nutr., 54: 1302-1309.
Direct Link - Jacobson, P., J. Uberti, W. Davis and V. Ratanatharathorn, 1998. Tacrolimus: A new agent for the prevention of graft-versus-host disease in hematopoietic stem cell transplantation. Bone Marrow Transplant., 22: 217-225.
PubMed - Cho, J.Y., A.R. Kim, E.S. Yoo, K.U. Baik and M.H. Park, 2002. Ginsenosides from Panax ginseng differentially regulate lymphocyte proliferation. Planta Med., 68: 497-500.
CrossRefPubMedDirect Link - Jarvis, J.N., G. Meintjes, K. Rebe, G.N. Williams and T. Bicanic et al., 2012. Adjunctive interferon-gamma immunotherapy for the treatment of HIV-associated cryptococcal meningitis: A randomized controlled trial. Aids, 26: 1105-1113.
Direct Link - Jeurink, P.V., B.C. van Esch, A. Rijnierse, J. Garssen and L.M. Knippels, 2013. Mechanisms underlying immune effects of dietary oligosaccharides. Am. J. Clin. Nutr., 98: 572S-577S.
Direct Link - Jin, Y., Y. Zhang, C. Wan, H. Wang and L. Hou et al., 2013. Immunomodulatory activity and protective effects of polysaccharide from Eupatorium adenophorum leaf extract on highly pathogenic H5N1 influenza infection. Evid Based Complement Alternat. Med., Vol. 2013.
CrossRefDirect Link - Joseph, B. and P. Sankarganesh, 2011. Antifungal efficacy of panchagavya. Int. J. PharmTech Res., 3: 585-588.
Direct Link - Kaila, M., E. Isolauri, E. Soppi, E. Virtanen, S. Laine and H. Arvilommi, 1992. Enhancement of the circulating antibody secreting cell response in human diarrhea by a human Lactobacillus strain. Pediatr. Res., 32: 141-144.
PubMed - Gaur, K., M.L. Kori and R.K. Nema, 2009. Comparative screening of immunomodulatory activity of hydro-alcoholic extract of Hibiscus rosa sinensis Linn. and ethanolic extract of Cleome gynandra Linn. Global J. Pharmacol., 3: 85-89.
Direct Link - Kamath, A.T., A.F. Rochat, D. Christensen, E.M. Agger, P. Andersen and P.H. Lambert, 2009. A liposome-based mycobacterial vaccine induces potent adult and neonatal multifunctional T cells through the exquisite targeting of dendriticcells. PLoS One, Vol. 4.
CrossRefDirect Link - Kanzler, H., F.J. Barrat, E.M. Hessel and R.L. Coffman, 2007. Therapeutic targeting of innate immunity with toll-like receptor agonists and antagonists. Nat. Med., 13: 552-559.
CrossRefPubMedDirect Link - Karbasi, A. and A. Saburi, 2012. Role of vitamin D on cancer immunotherapy. Front. Endocrinol., Vol. 3.
Direct Link - Karthik, K., I. Sophia, V. Rekha, M. Bhardwaj, N. Singh and A. Jain, 2012. Antibiotic resistance-counter attack by Salmonella. Int. J. Livestock Res., 2: 42-47.
Direct Link - Karthik, K., N.S. Muneeswaran, H.V. Manjunathachar, M. Gopi, A. Elamurugan and S. Kalaiyarasu, 2014. Bacteriophages: Effective alternative to antibiotics. Adv. Anim. Vet. Sci., Vol. 2.
Direct Link - Klotz, L.O., K.D. Kroncke, D.P. Buchczyk and H. Sies, 2003. Role of copper, zinc, selenium and tellurium in the cellular defense against oxidative and nitrosative stress. J. Nutr., 133: 1448-1451.
PubMed - Koenen, M.E., J. Kramer, R. van der Hulst, L. Heres, S.H.M. Jeurissen and W.J.A. Boersma, 2004. Immunomodulation by probiotic lactobacilli in layer- and meat-type chickens. Br. Poult. Sci., 45: 355-366.
CrossRefPubMedDirect Link - Kogut, M.H., 2000. Cytokines and prevention of infectious diseases in poultry: A review. Avian Pathol., 29: 395-404.
CrossRef - Kosaka, A., H. Yan, S. Ohashi, Y. Gotoh and A. Sato et al., 2012. Lactococcus lactis subsp. cremoris FC triggers IFN-γ production from NK and T cells via IL-12 and IL-18. Int. Immunopharmacol., 14: 729-733.
CrossRefPubMedDirect Link - Kugelberg, E., 2014. Glucocorticoids shift response in macrophages. Nat. Rev. Immunol., Vol. 14.
CrossRef - Kuhn, C.A. and C.W. Hanke, 1997. Current status of melanoma vaccines. Dermatol. Surg., 23: 649-654.
CrossRef - Kumar, A., A.K. Verma, A. Rahal, P.K. Panwar and K. Dhama, 2013. Recent trends in development of adjuvant of vaccine. Trends Med. Res., 8: 32-35.
CrossRefDirect Link - Kumar, S. and K.C. Anderson, 2005. Drug insight: Thalidomide as a treatment for multiple myeloma. Nat. Clin. Pract. Oncol., 2: 262-270.
CrossRef - Kunisawa, J. and H. Kiyono, 2013. Vitamin-mediated regulation of intestinal immunity. Frontiers Immunol., 4: 189-189.
Direct Link - Kunz, M. and S.M. Ibrahim, 2009. Cytokines and cytokine profiles in human autoimmune diseases and animal models of autoimmunity. Mediators Inflammation.
CrossRefDirect Link - Sunila, E.S. and G. Kuttan, 2004. Immunomodulatory and antitumor activity of Piper longum Linn. and piperine. J. Ethnopharmacol., 90: 339-346.
CrossRefDirect Link - Leclerc, C. and F.R. Vogel, 1985. Synthetic immunomodulators and synthetic vaccines. Crit. Rev. Therapeutic Drug Carrier Syst., 2: 353-406.
Direct Link - Lahteinen, T., A. Lindholm, T. Rinttila, S. Junnikkala and R. Kant et al., 2014. Effect of Lactobacillus brevis ATCC 8287 as a feeding supplement on the performance and immune function of piglets. Vet. Immunol. Immunopathol., 158: 14-25.
CrossRef - Lai, C.Y., J.T. Hung, H.H. Lin, A.L. Yu and S.H. Chen et al., 2010. Immunomodulatory and adjuvant activities of a polysaccharide extract of Ganoderma lucidum in vivo and in vitro. Vaccine, 28: 4945-4954.
CrossRef - Lakshmi, V., S.K. Joseph, S. Srivastava, S.K. Verma and M.K. Sahoo et al., 2010. Antifilarial activity in vitro and in vivo of some flavonoids tested against Brugia malayi. Acta Trop., 116: 127-133.
CrossRef - Lamprecht, M., S. Bogner, G. Schippinger, K. Steinbauer and F. Fankhauser et al., 2012. Probiotic supplementation affects markers of intestinal barrier, oxidation and inflammation in trained men: A randomized, double-blinded, placebo-controlled trial. J. Int. Soc. Sports Nutr., Vol. 9.
CrossRef - Latif, T., N. Chauhan, R. Khan, A. Moran and S.Z. Usmani, 2012. Thalidomide and its analogues in the treatment of Multiple Myeloma. Exp. Hematol. Oncol., Vol. 1.
Direct Link - Lederer, E., A. Adam, R. Ciorbaru, J.F. Petit and J. Wietzerbin, 1975. Cell walls of mycobacteria and related organisms: Chemistry and immunostimulant properties. Mol. Cell. Biochem., 7: 87-104.
CrossRefDirect Link - Lee, J.S., A. Synytsya, H.B. Kim, D.J. Choi and S. Lee et al., 2013. Purification, characterization and immunomodulating activity of a pectic polysaccharide isolated from Korean mulberry fruit Oddi (Morus alba L.). Int. Immunopharmacol., 17: 858-866.
CrossRef - Lee, S., H.S. Lillehoj, D.W. Park, Y.H. Hong and J.J. Lin, 2007. Effects of Pediococcus-and Saccharomyces-based probiotic (MitoMax®) on coccidiosis in broiler chickens. Comp. Immunol. Microbiol. Infect. Dis., 30: 261-268.
CrossRef - Lefrancois, L. and L. Puddington, 1995. Extrathymic intestinal T-cell development: Virtual reality? Immunol. Today, 16: 16-21.
CrossRef - Leonard, E.J. and T. Yoshimura, 1990. Neutrophil attractant/activation protein-1 (NAP-1 [interleukin-8]). Am. J. Respir Cell. Mol. Biol., 2: 479-486.
CrossRef - Leroux-Roels, G., 2010. Unmet needs in modern vaccinology: Adjuvants to improve the immune response. Vaccine, 28: C25-C36.
CrossRef - Nelson, H.S., D.Y. Leung and J.W. Bloom, 2003. Update on glucocorticoid action and resistance. J. Allergy Clin. Immunol., 111: 3-22.
CrossRef - Li, J., D.F. Li, J.J. Xing, Z.B. Cheng and C.H. Lai, 2006. Efects of β-glucan extracted from Saccharomyces cerevisiae on growth performance and immunological and somatotropic responses of pigs challenged with Escherichia coli lipopolysaccharide. J. Anim. Sci., 84: 2374-2381.
PubMed - Li, J., J. Xing, D. Li, X. Wang, L. Zhao, S. Lv and D. Huang, 2005. Effects of β-glucan extracted from Saccharomyces cerevisiae on humoral and cellular immunity in weaned piglets. Arch. Anim. Nutr., 59: 303-312.
CrossRefDirect Link - Li, P., X. Zhang, Y. Cheng, J. Li and Y. Xiao et al., 2014. Preparation and in vitro immunomodulatory effect of curdlan sulfate. Carbohydr. Polymers, 102: 852-861.
CrossRef - Lilly, D.M. and R.H. Stillwell, 1965. Probiotics: Growth-promoting factors produced by microorganisms. Science, 147: 747-748.
CrossRefDirect Link - Lin, C.C., C.L. Chu, C.S. Ng, C.Y. Lin, D.Y. Chen, I.H. Pan and K.J. Huang, 2014. Immunomodulation of phloretin by impairing dendritic cell activation and function. Food Funct., 5: 997-1006.
CrossRef - Lindenstrom, T., E.M. Agger, K.S. Korsholm, P.A. Darrah and C. Aagaard et al., 2009. Tuberculosis subunit vaccination provides long-term protective immunity characterized by multifunctional CD4 memory T cells. J. Immunol., 182: 8047-8055.
Direct Link - Lipford, G.B., H. Wagner and K. Heeg, 1994. Vaccination with immunodominant peptides encapsulated in Quil A-containing liposomes induces peptide-specific primary CD8+ cytotoxic T cells. Vaccine, 12: 73-80.
CrossRef - Liu, X., J. Zheng and H. Zhou, 2011. TLRs as pharmacological targets for plant-derived compounds in infectious and inflammatory diseases. Int. Immunopharmacol., 11: 1451-1456.
CrossRef - Long, K.Z. and J.I. Santos, 1999. Vitamins and the regulation of the immune response. Pediatric Infect. Dis. J., 18: 283-290.
Direct Link - Lowenthal, J.W., T.E. O'Neil, M. Broadway, A.D.G. Strom, M.R. Digby, M. Andrew and J.J. York, 1998. Coadministration of IFN-γ enhances antibody responses in chickens. J. Interferon Cytokine Res., 18: 617-622.
CrossRefDirect Link - Liu, Y., H. Jing, J. Wang, R. Zhang and Y. Zhang et al., 2011. Micronutrients decrease incidence of common infections in type 2 diabetic outpatients. Asia Pac. J. Clin. Nutr., 20: 375-382.
PubMedDirect Link - Maass, N., J. Bauer, B.R. Paulicks, B.M. Bohmer and D.A. Roth-Maier, 2005. Efficiency of Echinacea purpurea on performance and immune status in pigs. J. Anim. Physiol. Anim. Nutr., 89: 244-252.
CrossRefPubMedDirect Link - Maggini, S., E.S. Wintergerst, S. Beveridge and D.H. Hornig, 2007. Selected vitamins and trace elements support immune function by strengthening epithelial barriers and cellular and humoral immune responses. Br. J. Nutr., 98: 29-35.
Direct Link - Chaudhary, M., A.K. Garg, G.K. Mittal and V. Mudgal, 2010. Effect of organic selenium supplementation on growth, se uptake and nutrient utilization in guinea pigs. Biol. Trace Elem. Res., 133: 217-226.
CrossRefDirect Link - Mahima, A. Rahal, R. Deb, S.K. Latheef and H.A. Samad et al., 2012. Immunomodulatory and therapeutic potentials of herbal, traditional/indigenous and ethnoveterinary medicines. Pak. J. Biol. Sci., 15: 754-774.
CrossRefDirect Link - Mahima, A.K. Garg and V. Mudgal, 2012. Influence of sodium selenite on growth, nutrient utilization and selenium uptake in Cavia porcellus. Pak. J. Biol. Sci., 15: 448-453.
CrossRefDirect Link - Mahima, A.M. Ingle, A.K. Verma, R. Tiwari and K. Karthik et al., 2013. Immunomodulators in day to day life: A review. Pak. J. Biol. Sci., 16: 826-843.
CrossRefDirect Link - Mahima, A.K. Verma, R. Tiwari, K. Karthik, S. Chakraborty, R. Deb and K. Dhama, 2013. Nutraceuticals from fruits and vegetables at a glance: A review. J. Biol. Sci., 13: 38-47.
CrossRefDirect Link - Maisin, J., 1964. Existence in the thymus of a factor protecting the skin of the mouse against the induction of skin cancers induced by 20-methylcholanthrene. Nature, 204: 1211-1215.
CrossRefDirect Link - Majtan, J., 2014. Honey: An immunomodulator in wound healing. Wound Repair Regener., 22: 187-192.
CrossRef - Maloy, K.J., A.M. Donachie and A.M. Mowat, 1995. Induction of Th1 and Th2 CD4+ T cell responses by oral or parenteral immunization with ISCOMS. Eur. J. Immunol., 25: 2835-2841.
CrossRef - Mantzioris, E., L.G. Cleland, R.A. Gibson, M.A. Neumann, M. Demasi and M.J. James, 2000. Biochemical effects of a diet containing foods enriched with n-3 fatty acids. Am. J. Clin. Nutr., 72: 42-48.
PubMedDirect Link - Mastelic, B., S. Ahmed, W.M. Egan, G. del Giudice and H. Golding et al., 2010. Mode of action of adjuvants: Implications for vaccine safety and design. Biologicals, 38: 594-601.
CrossRefDirect Link - Mathivanan, R. and S.C. Edwin, 2012. Effects of alternatives to antibiotic growth promoters on intestinal content characteristics, intestinal morphology and gut flora in broilers. Wudpecker J. Agric. Res., 1: 244-249.
Direct Link - Mathivanan, R., S.C. Edwin, K. Viswanathan and D. Chandrasekaran, 2006. Chemical, microbial composition and antibacterial activity of modified panchagavya. Int. J. Cow Sci., 2: 23-26.
Direct Link - Mathivanan, R., S.C. Edwin, R. Amutha and K. Viswanathan, 2006. Panchagavya and Andrographis paniculata as alternatives to antibiotic growth promoter on broiler production and carcass characteristics. Int. J. Poult. Sci., 5: 1144-1150.
CrossRefDirect Link - McCarty, D.E., A.L. Chesson Jr., S.K. Jain and A.A. Marino, 2013. The link between vitamin D metabolism and sleep medicine. Sleep Med. Rev., 18: 311-319.
CrossRefDirect Link - McElhaney, J.E., R.N. Coler and S.L. Baldwin, 2013. Immunologic correlates of protection and potential role for adjuvants to improve influenza vaccines in older adults. Exp. Rev. Vaccines, 12: 759-766.
CrossRefDirect Link - McHugh, R.S., M.J. Whitters, C.A. Piccirillo, D.A. Young, E.M. Shevach, M. Collins and M.C. Byrne, 2002. CD4+CD25+ immunoregulatory T cells: Gene expression analysis reveals a functional role for the glucocorticoid-induced TNF receptor. Immunity, 16: 311-323.
CrossRefPubMedDirect Link - McSorley, H.J., J.P. Hewitson and R.M. Maizels, 2013. Immunomodulation by helminth parasites: Defining mechanisms and mediators. Int. J. Parasitol., 43: 301-310.
CrossRefDirect Link - McSorley, H.J., M.T. O'Gorman, N. Blair, T.E. Sutherland, K.J. Filbey and R.M. Maizels, 2012. Suppression of type 2 immunity and allergic airway inflammation by secreted products of the helminth Heligmosomoides polygyrus. Eur. J. Immunol., 42: 2667-2682.
CrossRef - Mehrotra, S., K.P. Mishra, R. Maurya, R.C. Srimal, V.S. Yadav, R. Pandey and V.K. Singh, 2003. Anticellular and immunosuppressive properties of ethanolic extract of Acorus calamus rhizome. Int. Immunopharmacol., 3: 53-61.
CrossRefDirect Link - Meydani, S.N., S.N. Han and D. Wu, 2005. Vitamin E and immune response in the aged: Molecular mechanisms and clinical implications. Immunol. Rev., 205: 269-284.
CrossRef - Miles, E.A. and P.C. Calder, 2012. Influence of marine n-3 polyunsaturated fatty acids on immune function and a systematic review of their effects on clinical outcomes in rheumatoid arthritis. Br. J. Nutr., 107: S171-S184.
CrossRefDirect Link - Mily, A., R.S. Rekha, S.M. Kamal, E. Akhtar and P. Sarker et al., 2013. Oral intake of phenylbutyrate with or without vitamin D3 upregulates the cathelicidin LL-37 in human macrophages: A dose finding study for treatment of tuberculosis. BMC Pulm. Med., Vol. 13.
CrossRef - Minagawa, T., K. Ishiwata and T. Kajimoto, 1999. Feline interferon-ω treatment on canine parvovirus infection. Vet. Microbiol., 69: 51-53.
CrossRefDirect Link - Minatel, L. and J.C. Carfagnini, 2000. Copper deficiency and immune response in ruminants. Nutr. Res., 20: 1519-1529.
CrossRefDirect Link - Mirzaei-Aghsaghali, A., 2012. Importance of medical herbs in animal feeding: A review. Ann. Biol. Res., 3: 918-923.
Direct Link - Mitchell, D., M. Yong, J. Raju, N. Willemsen and M. Black et al., 2011. Toll-like receptor-mediated adjuvanticity and immunomodulation in dendritic cells: Implications for peptide vaccines. Hum. Vaccines, 7: 85-93.
CrossRefDirect Link - Mocanu, V., T. Oboroceanu and F. Zugun-Eloae, 2013. Current status in vitamin D and regulatory T cells-immunological implications. Revista Medico-Chirurgicala Societatii Medici Naturalisti Iasi, 117: 965-973.
Direct Link - Moeini, M.M., A. Kiani, E. Mikaeili and H.K. Shabankareh, 2011. Effect of prepartum supplementation of selenium and Vitamin E on serum Se, IgG concentrations and colostrum of heifers and on hematology, passive immunity and Se status of their offspring. Biol. Trace Element Res., 144: 529-537.
CrossRefDirect Link - Mokrozub, V.V., L.M. Lazarenko, L.P. Babenko, L.M. Shynkarenko-Sichel and Z.M. Olevinska et al., 2012. Effect of probiotic strains of lacto- and bifidobacteria on the activity of macrophages and other parameters of immunity in cases of staphylococcosis. Mikrobiolohichnyi Zhurnal, 74: 90-98.
PubMedDirect Link - Molad, Y., 2002. Update on colchicine and its mechanism of action. Curr. Rheumatol. Rep., 4: 252-256.
CrossRefDirect Link - Moreno-Vilet, L., M.H. Garcia-Hernandez, R.E. Delgado-Portales, N.E. Corral-Fernandez, N. Cortez-Espinosa, M.A. Ruiz-Cabrera and D.P. Portales-Perez, 2014. In vitro assessment of agave fructans (Agave salmiana) as prebiotics and immune system activators. Int. J. Biol. Macromol., 63: 181-187.
CrossRefDirect Link - Mubarak, A., A. Rashid, I.A. Khan and A. Hussain, 2009. Effect of vitamin E and selenium as immunomodulators on induced aflatoxicosis in broiler birds. Pak. J. Life Soc. Sci., 7: 31-34.
Direct Link - Muir, W.I., A.J. Husband and W.L. Bryden, 2002. Dietary supplementation with vitamin E modulates avian intestinal immunity. Br. J. Nutr., 87: 579-585.
CrossRefDirect Link - Mukhtyar, C., L. Guillevin, M.C. Cid, B. Dasgupta and K. de Groot et al., 2009. EULAR recommendations for the management of primary small and medium vessel vasculitis. Ann. Rheumatic Dis., 68: 310-317.
CrossRefDirect Link - Munyaka, P.M., G. Tactacan, M. Jing, J.D. House and J.C. Rodriguez-Lecompte, 2012. Immunomodulation in young laying hens by dietary folic acid and acute immune responses after challenge with Escherichia coli lipopolysaccharide. Poult. Sci., 91: 2454-2463.
PubMed - Mutwiri, G., V. Gerdts, M. Lopez and L.A. Babiuk, 2007. Innate immunity and new adjuvants. Rev. Sci. Tech. Off. Epiz., 26: 147-156.
Direct Link - Nairz, M., A. Schroll, T. Sonnweber and G. Weiss, 2010. The struggle for iron-a metal at the host-pathogen interface. Cell. Microbiol., 12: 1691-1702.
CrossRef - Nicholas, R., P. Giannetti, A. Alsanousi, T. Friede and P.A. Muraro, 2011. Development of oral immunomodulatory agents in the management of multiple sclerosis. Drug Des. Dev. Ther., 5: 255-274.
Direct Link - Nicholls, E.F., L. Madera and R.E.W. Hancock, 2010. Immunomodulators as adjuvants for vaccines and antimicrobial therapy. Ann. N. Y. Acad. Sci., 1213: 46-61.
CrossRef - Nijnik, A. and R.E.W. Hancock, 2009. Host defence peptides: Antimicrobial and immunomodulatory activity and potential applications for tackling antibiotic-resistant infections. Emerg. Health Threats J., Vol. 2.
CrossRefDirect Link - Noe, S.M., M.A. Green, H. HogenEsch and S.L. Hem, 2010. Mechanism of immunopotentiation by aluminum-containing adjuvants elucidated by the relationship between antigen retention at the inoculation site and the immune response. Vaccine, 28: 3588-3594.
CrossRefPubMedDirect Link - Nutman, T.B. and V. Kumaraswami, 2001. Regulation of the immune response in lymphatic filariasis: Perspectives on acute and chronic infection with Wuchereria bancrofti in South India. Parasite Immunol., 28: 389-399.
CrossRefDirect Link - Ohashi, Y. and U. Ushida, 2009. Health-beneficial effects of probiotics: Its mode of action. Anim. Sci. J., 80: 361-371.
CrossRefDirect Link - Ohira, H., M. Matsunaga, T. Osumi, S. Fukuyama, J. Shinoda, J. Yamada and Y. Gidron, 2013. Vagal nerve activity as a moderator of brain-immune relationships. J. Neuroimmunol., 260: 28-36.
CrossRefDirect Link - Openheimer, S.J., 2001. Iron and its relation to immunity and infectious disease. J. Nutr., 131: 616S-633S.
PubMedDirect Link - Ortiz-Sanchez, E., G. Helguera, T.R. Daniels and M.L. Penichet, 2008. Antibody-cytokine fusion proteins: Applications in cancer therapy. Exp. Opin. Biol. Ther., 8: 609-632.
CrossRefDirect Link - Osada, Y., S. Yamada, A. Nabeshima, Y. Yamagishi and K. Ishiwata et al., 2013. Heligmosomoides polygyrus infection reduces severity of type 1 diabetes induced by multiple low-dose streptozotocin in mice via STAT6-and IL-10-independent mechanisms. Exp. Parasitol., 135: 388-396.
CrossRefDirect Link - Osredkar, J. and N. Sustar, 2011. Copper and zinc, biological role and significance of copper/zinc imbalance. J. Clin. Toxicol., Vol. S3.
CrossRefDirect Link - Pae, M., S.N. Meydani and D. Wu, 2012. The role of nutrition in enhancing immunity in aging. Aging Dis., 3: 91-129.
Direct Link - Park, J.H., J.I. Um, B.J. Lee, J.S. Goh, S.Y. Park, W.S. Kim and P.H. Kim, 2002. Encapsulated Bifidobacterium bifidum potentiates intestinal IgA production. Cell Immunol., 219: 22-27.
CrossRef - Patil, U.S., A.V. Jaydeokar and D.D. Bandawane, 2012. Immunomodulators: A pharmacological review. Int. J. Pharm. Pharm. Sci., 4: 30-36.
Direct Link - Persing, D.H., R.N. Coler, M.J. Lacy, D.A. Johnson, J.R. Baldridge, R.M. Hershberg and S.G. Reed, 2002. Taking toll: Lipid a mimetics as adjuvants and immunomodulators. Trends Microbiol., 10: 32-37.
PubMed - Peshev, D. and W. van den Ende, 2014. Fructans: Prebiotics and immunomodulators. J. Funct. Foods, 8: 348-357.
CrossRefDirect Link - Pfahlberg, A., K.F. Kolmel, J.M. Grange, G. Mastrangelo and B. Krone et al., 2002. Inverse association between melanoma and previous vaccinations against tuberculosis and smallpox: Results of the FEBIM study. J. Invest. Dermatol., 119: 570-575.
CrossRefDirect Link - Philips, A., M. Maira, A. Mullick, M. Chamberland, S. Lesage, P. Hugo and J. Drouin, 1997. Antagonism between Nur77 and glucocorticoid receptor for control of transcription. Mol. Cell. Biol., 17: 5952-5959.
PubMed - Pillemer, L.A. and E.E. Ecker, 1941. Anticomplementary factor in fresh yeast. J. Biol. Chem., 137: 139-142.
Direct Link - Pindolia, K., H. Li, C. Cardwell and B. Wolf, 2014. Characterization and functional analysis of cellular immunity in mice with biotinidase deficiency. Mol. Gen. Metabol., 112: 49-56.
CrossRef - Pirofski, L.A. and A. Casadevall, 2006. Immunomodulators as an antimicrobial tool. Curr. Opin. Microbial., 9: 489-495.
CrossRefDirect Link - Pond, C.M., 2005. Adipose tissue and the immune system. Prostaglandins, Leukotrienes Essent. Fatty Acids, 73: 17-30.
CrossRefDirect Link - Powers, J.P.S. and R.E. Hancock, 2003. The relationship between peptide structure and antibacterial activity. Peptides, 24: 1681-1691.
CrossRefDirect Link - Prabhala, R.H., H.S. Garewal, M.J. Hicks, R.E. Sampliner and R.R. Watson, 1991. The effects of 13‐cis‐retinoic acid and beta‐carotene on cellular immunity in humans. Cancer, 67: 1556-1560.
CrossRefDirect Link - Prasad, A.S., 2000. Effects of zinc deficiency on immune functions. J. Trace Elem. Exp. Med., 13: 1-20.
CrossRefDirect Link - Prevot, A.R., B. Halpern, G. Biozzi, C. Stiffel and D. Mouton et al., 1963. [Stimulation of the reticuloendothelial system (RES) by killed microbial substances from Corynebacterium parvum]. Hebd Seances Acad. Sci., 257: 13-17, (In French).
PubMed - Prins, R.M., N. Craft, K.W. Bruhn, H. Khan-Farooqi and R.C. Koya et al., 2006. The TLR-7 agonist, imiquimod, enhances dendritic cell survival and promotes tumor antigen-specific T cell priming: Relation to central nervous system antitumor immunity. J. Immunol., 176: 157-164.
CrossRefDirect Link - Sarker, P., S. Ahmed, S. Tiash, R.S. Rekha and R. Stromberg et al., 2011. Phenylbutyrate counteracts Shigella mediated downregulation of cathelicidin in rabbit lung and intestinal epithelia: A potential therapeutic strategy. PLoS One, Vol. 6.
CrossRefDirect Link - Puertollano, M.A., E. Puertollano, G.A. de Cienfuegos and M.A. de Pablo, 2011. Dietary antioxidants: Immunity and host defense. Curr. Top. Med. Chem., 11: 1752-1766.
CrossRefDirect Link - Pulverer, G., H.L. Ko, K. Roszkowski, W. Roszkowski and J. Jeljaszewicz, 1985. Immunomodulation by propionibacteria. Clin. Immunol. Newslett., 6: 51-54.
CrossRefDirect Link - Quintanar, J.L. and I. Guzman-Soto, 2013. Hypothalamic neurohormones and immune responses. Front. Integr. Neurosci., Vol. 7.
CrossRefDirect Link - Rahal, A., Mahima, A.K. Verma, A. Kumar and R. Tiwari et al., 2014. Phytonutrients and nutraceuticals in vegetables and their multi-dimensional medicinal and health benefits for humans and their companion animals: A review. J. Biol. Sci., 14: 1-19.
CrossRefDirect Link - Rahal, A., A. Kumar, V. Singh, B. Yadav, R. Tiwari, S. Chakraborty and K. Dhama, 2014. Oxidative stress, prooxidants and antioxidants: The interplay. BioMed Res. Int., Vol. 2014.
CrossRefDirect Link - Randhawa, G.K., 2010. Cow urine distillate as bioenhancer. J. Ayurveda Integr. Med., 1: 240-241.
CrossRef - Raqib, R., P. Sarker, P. Bergman, G. Ara and M. Lindh et al., 2006. Improved outcome in shigellosis associated with butyrate induction of an endogenous peptide antibiotic. Proc. Nat. Acad. Sci. USA., 103: 9178-9183.
PubMed - Rask, C., I. Adlerberth, A. Berggren, I.L. Ahren and A.E. Wold, 2013. Differential effect on cell‐mediated immunity in human volunteers after intake of different lactobacilli. Clin. Exp. Immunol., 172: 321-332.
CrossRefDirect Link - Reddy, M.V., 2010. Immunomodulators of helminthes: Promising therapeutics for autoimmune disorders and allergic diseases x. Indian J. Clin. Biochem., 25: 109-110.
CrossRefDirect Link - Ren, F., X. Chen, J. Hesketh, F. Gan and K. Huang, 2012. Selenium promotes T-cell response to TCR-stimulation and ConA, but not PHA in primary porcine splenocytes. PloS One, Vol. 7.
CrossRefDirect Link - Renoux, G. and M. Renoux, 1971. [Immunostimulating effect of an imidotiiazole in the immunization of mice against Brucella abortus infection]. C. R. Acad. Sci. Hebd. Seances Acad. Sci. D, 272: 349-350, (In French).
PubMed - Rice, A.D., M.M. Adams, S.F. Lindsey, D.M. Swetnam and B.R. Manning et al., 2014. Protective properties of vaccinia virus-based vaccines: Skin scarification promotes a nonspecific immune response that protects against orthopoxvirus disease. J. Virol., 88: 7753-7763.
CrossRefDirect Link - Ritz, N., M. Mui, A. Balloch and N. Curtis, 2013. Non-specific effect of bacille calmette-guerin vaccine on the immune response to routine immunisations. Vaccine, 31: 3098-3103.
CrossRefDirect Link - Rivero-Lezcano, O.M., 2008. Cytokines as immunomodulators in tuberculosis therapy. Recent Patents Anti-Infect. Drug Discov., 3: 168-176.
CrossRefDirect Link - Robb, R.J., A. Munck and K.A. Smith, 1981. T cell growth factor receptors. Quantitation, specificity and biological relevance. J. Exp. Med., 154: 1455-1474.
PubMedDirect Link - Romagne, F., 2007. Current and future drugs targeting one class of innate immunity receptors: The Toll-like receptors. Drug Discovery Today, 12: 80-87.
CrossRefDirect Link - Roth, J.A. and M.L. Kaeberle, 1985. In vivo effect of ascorbic acid on neutrophil function in healthy and dexamethasone-treated cattle. Am. J. Vet. Res., 46: 2434-2436.
Direct Link - Ryan-Harshman, M. and W. Aldoori, 2005. The relevance of selenium to immunity, cancer and infectious/inflammatory diseases. Can. J. Diet. Pract. Res., 66: 98-102.
PubMedDirect Link - Saeed, M.A., I. Ahmad, U. Yaqub, S. Akbar, A. Waheed, M. Saleem and Nasir-ud-Din, 2003. Aloe vera: A plant of vital significance. Sci. Vision, 9: 1-13.
Direct Link - Safaralizadeh, R., M. Nourizadeh, A. Zare, G.A. Kardar and Z. Pourpak, 2013. Influence of selenium on mast cell mediator release. Biol. Trace Element Res., 154: 299-303.
CrossRefDirect Link - Sakaguchi, S., N. Sakaguchi, M. Asano, M. Itoh and M. Toda, 1995. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol., 155: 1151-1164.
Direct Link - Saroj, P., M. Verma and K.K. Jha, 2012. An overview on immunomodulation. J. Adv. Sci. Res., 3: 7-12.
Direct Link - Scaringi, L., P. Marconi, M. Boccanera, L. Tissi, F. Bistoni and A. Cassone, 1988. Cell wall components of Candida albicans as immunomodulators: Induction of natural killer and macrophage-mediated peritoneal cell cytotoxicity in mice by mannoprotein and glucan Fractions. Microbiology, 134: 1265-1274.
Direct Link - Schaible, U.E. and S.H.E. Kaufmann, 2004. Iron and microbial infection. Nat. Rev. Microbiol., 2: 946-953.
CrossRefDirect Link - Scott, M.G., E. Dullaghan, N. Mookherjee, N. Glavas and M. Waldbrook et al., 2007. An anti-infective peptide that selectively modulates the innate immune response. Nat. Biotechnol., 25: 465-472.
CrossRefDirect Link - Serban, D.E., 2014. Gastrointestinal cancers: Influence of gut microbiota, probiotics and prebiotics. Cancer Lett., 345: 258-270.
CrossRefDirect Link - Sharma, R., A. Rohilla and V. Arya, 2011. A short review on pharmacology of plant immunomodulators. Int. J. Pharmaceut. Sci. Rev. Res., 9: 126-131.
Direct Link - Shepherd, A.J., J.E.G. Downing and J.A. Miyan, 2005. Without nerves, immunology remains incomplete-in vivo veritas. Immunology, 116: 145-163.
CrossRefDirect Link - Short, S.C., D. Traish, A. Dowe, F. Hines, M. Gore and M. Brada, 2001. Thalidomide as an anti-angiogenic agent in relapsed gliomas. J. Neurooncol., 51: 41-45.
CrossRefDirect Link - Singh, M. and D.T. O'Hagan, 2003. Recent advances in veterinary vaccine adjuvants. Int. J. Parasitol., 33: 469-478.
CrossRefDirect Link - Singh, V.K., S. Biswas, K.B. Mathur, W. Haq, S.K. Garg and S.S. Agarwal, 1998. Thymopentin and splenopentin as immunomodulators. Immunol. Res., 17: 345-368.
CrossRefPubMedDirect Link - Singh, V., S. Amdekar and O. Verma, 2010. Ocimum sanctum (Tulsi): Bio-pharmacological activities. Pharmacology, Vol. 1, No. 10.
CrossRefDirect Link - Sohn, K.S., M.K. Kim, J.D. Kim and I.K. Han, 2000. The role of immunostimulants in monogastric animal and fish-review. Asian Austr. J. Anim. Sci., 13: 1178-1187.
Direct Link - Solaiman, S.G., T.J. Craig Jr., G. Reddy and C.E. Shoemaker, 2007. Effect of high levels of Cu supplement on growth performance, rumen fermentation and immune responses in goat kids. Small Rumin. Res., 69: 115-123.
CrossRefDirect Link - Sorup, S., M. Villumsen, H. Ravn, C.S. Benn and T.I. Sorensen et al., 2011. Smallpox vaccination and all-cause infectious disease hospitalization: A Danish register-based cohort study. Int. J. Epidemiol., 40: 955-963.
Direct Link - Souvannavong, V., S. Brown and A. Adam, 1988. The synthetic immunomodulator muramyl dipeptide (MDP) can stimulate activated B cells. Mol. Immunol., 25: 385-391.
CrossRefDirect Link - Spelman, K., J. Burns, D. Nichols, N. Winters, S. Ottersberg and M. Tenborg, 2006. Modulation of cytokine expression by traditional medicines: A review of herbal immunomodulators. Altern. Med. Rev., 11: 128-150.
PubMedDirect Link - Spisek, R., J. Brazova, D. Rozkova, K. Zapletalova, A. Sediva and J. Bartunkova, 2004. Maturation of dendritic cells by bacterial immunomodulators. Vaccine, 22: 2761-2768.
CrossRefDirect Link - Stanford, M., 2004. Interferon treatment of circovirus infection in grey parrots (Psittacuse erithacus). Vet. Rec., 154: 435-436.
Direct Link - Steinman, L., 2005. Blocking adhesion molecules as therapy for multiple sclerosis: Natalizumab. Nat. Rev. Drug Discovery, 4: 510-518.
CrossRefDirect Link - Stevceva, L., M. Moniuszko and M.G. Ferrari, 2006. Utilizing IL-12, IL-15 and IL-7 as mucosal vaccine adjuvants. Lett. Drug Des. Discovery, 3: 586-592.
CrossRefDirect Link - Stevenson, H.C., I. Green, J.M. Hamilton, B.A. Calabro and D.R. Parkinson, 1991. Levamisole: Known effects on the immune system, clinical results and future applications to the treatment of cancer. J. Clin. Oncol., 9: 2052-2066.
Direct Link - Stils, H.F., 2005. Adjuvants and antibody production: Dispelling the myths associated with Freund's complete and other adjuvants. ILAR J., 46: 280-293.
Direct Link - Sugimura, T., K. Jounai, K. Ohshio, T. Tanaka, M. Suwa and D. Fujiwara, 2013. Immunomodulatory effect of Lactococcus lactis JCM5805 on human plasmacytoid dendritic cells. Clin. Immunol., 149: 509-518.
CrossRef - Sultana, N. and Z.S. Saify, 2012. Naturally occurring and synthetic agents as potential anti-inflammatory and immunomodulants. Antiinflamm Antiallergy Agents Med. Chem., 11: 3-19.
CrossRefDirect Link - Sumithra, A., P. Srinivasan, G.A. Balasubramaniam, T.R.G. Murthy and P. Balachandran, 2013. Ameliorative effect of panchagavya on newcastle disease in layer chicken. Int. J. Agric. Biosci., 2: 60-63.
Direct Link - Sun, P., J.Q. Wang and H.T. Zhang, 2010. Effects of Bacillus subtilis natto on performance and immune function of preweaning calves. J. Dairy Sci., 93: 5851-5855.
CrossRefDirect Link - Sunkara, L.T., M. Achanta, N.B. Schreiber, Y.R. Bommineni and G. Dai et al., 2011. Butyrate enhances disease resistance of chickens by inducing antimicrobial host defense peptide gene expression. PLos One, Vol. 6.
CrossRefDirect Link - Sunkara, L.T., W. Jiang and G. Zhang, 2012. Modulation of antimicrobial host defense peptide gene expression by free fatty acids. PLoS One, Vol. 7.
CrossRefDirect Link - Sutterwala, F.S., Y. Ogura and R.A. Flavell, 2007. The inflammasome in pathogen recognition and inflammation. J. Leukocyte Biol., 82: 259-264.
CrossRefDirect Link - Sutterwala, F.S., Y. Ogura, M. Szczepanik, M. Lara-Tejero and G.S. Lichtenberger et al., 2006. Critical role for NALP3/CIAS1/Cryopyrin in innate and adaptive immunity through its regulation of caspase-1. Immunity, 24: 317-327.
CrossRefDirect Link - Suzuki, Y., Y. Adachi, N. Ohno and T. Yadomae, 2001. Th1/Th2-balancing immunomodulating activity of gel-forming (1→3)-β-glucans from Fungi. Biol. Pharmaceut. Bull., 24: 811-819.
CrossRefDirect Link - Svoboda, M., K. Nechvatalova, J. Krejci, J. Drabek, R. Ficek and M. Faldyna, 2007. The absence of iron deficiency effect on the humoral immune response of piglets to tetanus toxoid. Vet. Med., 52: 179-185.
Direct Link - Tamura, J., K. Kubota, H. Murakami, M. Sawamura and T. Matsushima et al., 1999. Immunomodulation by vitamin B12: Augmentation of CD8+ T lymphocytes and Natural Killer (NK) cell activity in vitamin B12-deficient patients by methyl-B12 treatment. Clin. Exp. Immunol., 116: 28-32.
CrossRefDirect Link - Thanendrarajan, S., M. Nowak, H. Abken and I.G.H. Schmidt-Wolf, 2011. Combining cytokine-induced killer cells with vaccination in cancer immunotherapy: More than one plus one? Leukemia Res., 35: 1136-1142.
CrossRefDirect Link - Tirilomis, T., 2014. Daptomycin and its immunomodulatory effect: Consequences for antibiotic treatment of methicillin-resistant staphylococcus aureus wound infections after heart surgery. Front Immunol., Vol. 5.
CrossRefDirect Link - Tiwari, R., S. Chakraborty, K. Dhama, S. Rajagunalan and S.V. Singh, 2013. Antibiotic resistance-an emerging health problem: Causes, worries, challenges and solutions: A review. Int. J. Curr. Res., 5: 1880-1892.
Direct Link - Tiwari, R., S. Chakraborty, M. Saminathan, K. Dhama and S.V. Singh, 2014. Ashwagandha (Withania somnifera): Role in safeguarding health, immunomodulatory effects, combating infections and therapeutic applications: A review. J. Biol. Sci., 14: 77-94.
Direct Link - Tiwari, R., A.K. Verma, S. Chakraborty, K. Dhama and S.V. Singh, 2014. Neem (Azadirachta indica) and its potential for safeguarding health of animals and humans: A review. J. Biol. Sci., 14: 110-123.
CrossRefDirect Link - Trapero-Marugan, M., L. Garcia-Buey, C. Munoz, N.E. Quintana and J.A. Moreno-Monteagudo et al., 2006. Sustained virological response to peginterferon plus ribavirin in chronic hepatitis C genotype 1 patients is associated with a persistent Th1 immune response. Aliment. Pharmacol. Ther., 24: 117-128.
CrossRefPubMedDirect Link - Travis, S., L.M. Yap, C. Hawkey, B. Warren, M. Lazarov, T. Fong and R.J. Tesi, 2005. RDP58 is a novel and potentially effective oral therapy for ulcerative colitis. Inflammatory Bowel Dis., 11: 713-719.
CrossRefDirect Link - Trebichavsky, I. and I. Splichal, 2006. Probiotics manipulate host cytokine response and induce antimicrobial peptides. Folia Microbiol., 51: 507-510.
CrossRefDirect Link - Trevisi, P., G. Merialdi, M. Mazzoni, L. Casini and C. Tittarelli et al., 2007. Effect of dietary addition of thymol on growth, salivary and gastric function, immune response and excretion of Salmonella enterica serovar typhimurium, in weaning pigs challenged with this microbe strain. Ital. J. Anim. Sci., 6: 374-376.
CrossRefDirect Link - Tsai, Y.T., P.C. Cheng and T.M. Pan, 2012. The immunomodulatory effects of lactic acid bacteria for improving immune functions and benefits. Applied Microbiol. Biotechnol., 96: 853-862.
CrossRefPubMedDirect Link - Tsoni, S.V. and G.D. Brown, 2008. β-glucans and dectin-1. Ann. New York Acad. Sci., 1143: 45-60.
CrossRefDirect Link - Tuchscherer, M., W. Otten, E. Kanitz, M. Grabner and A. Tuchscherer et al., 2012. Effects of inadequate maternal dietary protein: Carbohydrate ratios during pregnancy on offspring immunity in pigs. BMC Vet. Res., Vol. 8.
CrossRef - Tzianabos, A.O., 2000. Polysaccharide immunomodulators as therapeutic agents: Structural aspects and biologic function. Clin. Microbiol. Rev., 13: 523-533.
Direct Link - Uchakin, P.N., O.N. Uchakina, B.V. Tobin and F.I. Ershov, 2007. Neuroendocrine immunomodulation. Vestn. Ross Akad. Med. Nauk., 9: 26-32.
PubMed - Van de Veerdonk, F.L., B.J. Kullberg and M.G. Netea, 2012. Adjunctive immunotherapy with recombinant cytokines for the treatment of disseminated candidiasis. Clin. Microbiol. Infect., 18: 112-119.
CrossRefPubMedDirect Link - Van de Garde, M.D.B., F.O. Martinez, B.N. Melgert, M.N. Hylkema, R.E. Jonkers and J. Hamann, 2014. Chronic exposure to glucocorticoids shapes gene expression and modulates innate and adaptive activation pathways in macrophages with distinct changes in leukocyte attraction. J. Immunol., 192: 1196-1208.
PubMedDirect Link - Veldman, C.M., M.T. Cantorna and H.F. DeLuca, 2000. Expression of 1,25-dihydroxyvitamin D3 receptor in the immune system. Arch. Biochem. Biophys., 374: 334-338.
CrossRefPubMedDirect Link - Venturi, S., F.M. Donati, A. Venturi, M. Venturi, L. Grossi and A. Guidi, 2000. Role of iodine in evolution and carcinogenesis of thyroid, breast and stomach. Adv. Clin. Pathol., 4: 11-17.
PubMed - Verma, A.K., K. Dhama, S. Chakraborty, A. Kumar and R. Tiwari et al., 2014. Strategies for combating and eradicating important infectious diseases of animals with particular reference to India: Present and future perspectives. Asian J. Anim. Vet. Adv., 9: 77-106.
CrossRefDirect Link - Vetskova, E.K., M.N. Muhtarova, T.I. Avramov, T.R. Stefanova, I.J. Chalakov and M.H. Nikolova, 2013. Immunomodulatory effects of BCG in patients with recurrent respiratory papillomatosis. Folia Medica, 55: 49-54.
PubMed - Vilcek, J., M.H. Ng, A.E. Friedman-Kien and T. Krawciw, 1968. Induction of interferon synthesis by synthetic double-stranded polynucleotides. J. Virol., 2: 648-650.
Direct Link - Walter, B.M. and G. Bilkei, 2004. Immunostimulatory effect of dietary oregano etheric oils on lymphocytes from growth-retarded, low-weight growing-finishing pigs and productivity. Tijdschrift Voor Diergeneeskunde, 129: 178-181.
PubMedDirect Link - Wang, H., E.R. Rayburn, W. Wang, E.R. Kandimalla, S. Agrawal and R. Zhang, 2006. Immunomodulatory oligonucleotides as novel therapy for breast cancer: Pharmacokinetics, in vitro and in vivo anticancer activity and potentiation of antibody therapy. Mol. Cancer Ther., 5: 2106-2114.
PubMed - Weilhammer, D.R., C.D. Blanchette, N.O. Fischer, S. Alam and G.G. Loots et al., 2013. The use of nanolipoprotein particles to enhance the immunostimulatory properties of innate immune agonists against lethal influenza challenge. Biomaterials, 34: 10305-10318.
CrossRefPubMedDirect Link - Weiss, G., 2002. Iron and immunity: A double-edged sword. Eur. J. Clin. Investigat, 32: 70-78.
CrossRefPubMedDirect Link - Wenzel, B.E., A. Chow, R. Baur, H. Schleusener and J.R. Wall, 1998. Natural killer cell activity in patients with Graves' disease and Hashimoto's thyroiditis. Thyroid, 8: 1019-1022.
PubMedDirect Link - Wilckens, T. and R. de Rijk, 1997. Glucocorticoids and immune function: Unknown dimensions and new frontiers. Immunol. Today, 18: 418-424.
PubMedDirect Link - Wintergerst, E.S., S. Maggini and D.H. Hornig, 2007. Contribution of selected vitamins and trace elements to immune function. Ann. Nutr. Metab., 51: 301-323.
PubMed - Wintergerst, E.S., S. Maggini and D.H. Hornig, 2006. Immune-enhancing role of vitamin C and zinc and effect on clinical conditions. Ann. Nutr. Metab., 50: 85-94.
CrossRefPubMedDirect Link - Sun, H.X., Y. Xie and Y.P. Ye, 2009. Advances in saponin-based adjuvants. Vaccine, 27: 1787-1796.
CrossRefPubMedDirect Link - Yang, F.L., K.F. Hua, Y.L. Yang, W. Zou and Y.P. Chen et al., 2008. TLR-independent induction of human monocyte IL-1 by phosphoglycolipids from thermophilic bacteria. Glycoconjugate J., 25: 427-439.
CrossRefPubMedDirect Link - Yang, Y.L., Yang, F.L., S.C. Jao, M.Y. Chen, S.S. Tsay, W. Zou and S.H. Wu, 2006. Structural elucidation of phosphoglycolipids from strains of the bacterial thermophiles Thermus and Meiothermus. J. Lipid Res., 47: 1823-1832.
CrossRefPubMedDirect Link - Yazdi, M.H., M. Mahdavi, B. Varastehmoradi, M.A. Faramarzi and A.R. Shahverdi, 2012. The immunostimulatory effect of biogenic selenium nanoparticles on the 4T1 breast cancer model: An in vivo study. Biol. Trace Element Res., 149: 22-28.
CrossRefPubMedDirect Link - Yu, Z., Y. Yin, W. Zhao, F. Chen and J. Liu, 2014. Application and bioactive properties of proteins and peptides derived from hen eggs: Opportunities and challenges. J. Sci. Food Agric., 94: 2839-2845.
CrossRefPubMedDirect Link - Yuan, S.L., X.S. Piao, D.F. Li, S.W. Kim, H.S. Lee and P.F. Guo, 2006. Effects of dietary Astragalus polysaccharide on growth performance and immune function in weaned pigs. Anim. Sci., 62: 501-507.
CrossRefDirect Link - Zaccone, P. and A. Cooke, 2013. Vaccine against autoimmune disease: Can helminths or their products provide a therapy? Curr. Opin. Immunol., 25: 418-423.
CrossRefPubMedDirect Link - Zasloff, M., 2002. Antimicrobial peptides of multicellular organisms. Nature, 415: 389-395.
CrossRefDirect Link - Zatz, M.M., J. Oliver, C. Samuels, A.B. Skotnicki, M.B. Sztein and A.L. Goldstein, 1984. Thymosin increases production of T-cell growth factor by normal human peripheral blood lymphocytes. Proc. Natl. Acad. Sci. USA., 81: 2882-2885.
PubMed - Zeng, X., L.T. Sunkara, W. Jiang, M. Bible and S. Carter et al., 2013. Induction of porcine host defense peptide gene expression by short-chain fatty acids and their analogs. PLoS ONE, Vol. 8.
CrossRefDirect Link - Zhang, Z.W., Q.H. Wang, J.L. Zhang, S. Li, X.L. Wang and S.W. Xu, 2012. Effects of oxidative stress on immunosuppression induced by selenium deficiency in chickens. Biol. Trace Element Res., 149: 352-361.
CrossRefPubMedDirect Link - Zhang, X., J. Li, D.K.W. Wong, T.E. Wagner and Y. Wei, 2009. Fermented noni exudate-treated dendritic cells directly stimulate B lymphocyte proliferation and differentiation. Oncol. Rep., 21: 1147-1152.
CrossRefDirect Link - Zhao, S.J., F.J. Sun, E.J. Tian and Z.P. Chen, 2008. [The effects of iodine/selenium on the function of antigen presentation of peritoneal macrophages in rats]. Zhonghua Yu Fang Yi Xue Za Zhi, 42: 485-488, (In Chinese).
PubMed - Zhao, X., R. Li, J. Huang, J. Li, M. Hou and J. Zhong, 2012. Association of some physiological factors and milk performance in Chinese holstein. Asian J. Anim. Vet. Adv., 7: 1356-1363.
CrossRefDirect Link - Zhou, Q., L. Wang, H. Wang, F. Xie and T. Wang, 2012. Effect of dietary vitamin C on the growth performance and innate immunity of juvenile cobia (Rachycentron canadum). Fish Shellfish Immunol., 32: 969-975.
CrossRefPubMedDirect Link - Zimmerman, T., 2009. Immunomodulatory agents in oncology. Update Cancer Therapeut., 3: 170-181.
CrossRefDirect Link - Zoran, D.L., 2010. Obesity in dogs and cats: A metabolic and endocrine disorder. Vet. Clin. North Am.: Small Anim. Pract., 40: 221-239.
CrossRefDirect Link - Petrovsky, N. and J.C. Aguilar, 2004. Vaccine adjuvants: Current state and future trends. Immunol. Cell Biol., 82: 488-496.
CrossRefPubMedDirect Link