
Mayada R. Farag
Department of Forensic Medicine and Toxicology, Faculty of Veterinary Medicine, Zagazig University, Zagazig, 44111, Egypt
Mahmoud M. Alagawany
Department of Poultry, Faculty of Agriculture, Zagazig University, Zagazig, 44111, Egypt
Kuldeep Dhama
Division of Pathology, Indian Veterinary Research Institute, Izatnagar, Bareilly, 243122, Uttar Pradesh, India
International Journal of Pharmacology
Year: 2014 | Volume: 10 | Issue: 8 | Page No.: 429-439







ABSTRACT
This study was conducted to verify whether Turmeric (Curcuma longa) has the potential to modulate the cytogenotoxic and immunotoxic effects resulting from exposure to endosulfan in broiler chicks. For this purpose, 180 one day old chicks were divided randomly into 6 groups of 30 chicks each, divided into 3 replicates. First group was fed basal diet, while the other five groups were fed basal diet supplemented with 5 g turmeric, 10 g turmeric, 30 mg endosulfan, 30 mg endosulfan plus 5 g turmeric and 30 mg endosulfan plus 10 g turmeric/kg diet, respectively, during the experimental period. At the end of the experiment, blood and spleen samples were collected and evaluated for genotoxicity, cytotoxicity and immunotoxicity biomarkers induced by endosulfan. Also, histopathological investigation of the spleen tissue was conducted. A significant increase in genotoxicity and cytotoxicity biomarkers (micronucleusfrequency, DNA fragmentation, 8-hydroxy-2-deoxyguanosine level (8 OHdG) and lactate dehydrogenase activity (LDH)) was recorded in endosulfan treated chicks. In addition, the immunotoxicity biomarkers showed a significant decrease in phagocytic percentage, stimulated Nitric oxide (NO) production and decrease in the serum lysozyme activity. Histopathological findings supported the cytotoxicity results. Dietary supplementation of turmeric at different levels ameliorated deleterious effects of endosulfan and restored them to be near control level. The results of the study suggested that regular consumption of turmeric in the diet is a promising approach in attenuating the disturbances induced by endosulfan in the most studied biomarkers.
PDF Abstract XML References Citation
Received: August 11, 2014;
Accepted: October 15, 2014;
Published: November 28, 2014
How to cite this article
Mayada R. Farag, Mahmoud M. Alagawany and Kuldeep Dhama, 2014. Antidotal Effect of Turmeric (Curcuma longa) against Endosulfan-Induced Cytogenotoxicity and Immunotoxicity in Broiler Chicks. International Journal of Pharmacology, 10: 429-439.
DOI: 10.3923/ijp.2014.429.439
URL: https://scialert.net/abstract/?doi=ijp.2014.429.439
DOI: 10.3923/ijp.2014.429.439
URL: https://scialert.net/abstract/?doi=ijp.2014.429.439
INTRODUCTION
Pesticide-related adverse effects in humans, animals and birds have become a serious public health concern. The exposure of non-target species to these toxic chemicals is virtually unavoidable due to their presence in air, water, soil and food worldwide. Among all forms of chemical pesticides, organochlorines are considered to be the most hazardous with respect to environmental pollution, since they are very persistent, non-biodegradable, add to the residue buildup in the food chain, cause eco-system imbalances and result in various biological disorders. Their residues are found in vegetables, grains, fruits, meat, eggs and even in human and animal blood and tissues (Garg et al., 2004). In Egypt, all types of organochlorines (OCPs) have been banned for agriculture and public health reasons. However, due to their cheap price, easy to use and effective in eradicating pests, some kinds of them are still widely employed, coupled with a lack of law enforcement (Nasr et al., 2009). Endosulfan (C9H6Cl6O3S), a compound of the cyclodiene subgroup of OCPs, is used extensively worldwide in agriculture and horticulture as a contact and stomach insecticide and as an acaricide on field cereals, oil seeds, coffee, cotton, vegetables, potato, tea and fruit crops (Vorkamp et al., 2004). One of the major routes of exposure in non-target species including birds is by ingestion of food contaminated with the endosulfan residues as a result of application or bioaccumulation and also via pulmonary and dermal routes. Its residues have been detected in a variety of different wild and domestic mammalian species, fish and in imported and locally raised chicken and bovine meat in Egypt (Aboul-Enein et al., 2010). The presence of residues in avian species suggests the plausibility of endosulfan to induce toxicopathological effects in these species. These suggestions are supported by some studies which have shown that endosulfan exposure resulted in some toxicological effects and immune dysfunction in avian and mammalian species (Aggarwal et al., 2009). The genotoxicity of endosulfan was also demonstrated (Lu et al., 2000).
Recently, turmeric (Curcuma longa) which is an extensively used spice, food preservative and coloring material having multiple biological actions and medicinal applications has received attention as a component of designer foods for its antioxidant properties (Akbarian et al., 2012). Curcumin is the main component responsible for its biological activity, but this herb also contains other components such as curcuminoids and polypeptides with biological action (Mesa and Ramirez-Tortosa, 2000). A number of studies have been conducted to evaluate its effect on the performance of broiler chickens (Hosseini-Vashan et al., 2012). However, information about the antagonistic effect of Curcuma longa on the immune dysfunction and cytogenotoxic endpoints of endosulfan in broiler chicks is scarce. Therefore, the present experiment was conducted to investigate the attenuating role of turmeric on the possible endosulfan-induced adverse effects on the cytogenotoxicity and immunotoxicity biomarkers. Besides, the ameliorating effect of turmeric on the pathological abnormalities in spleen tissues of endosulfan treated broiler chicks was also examined.
MATERIALS AND METHODS
The present study was carried out at Poultry Research Farm, Faculty of Veterinary Medicine, Zagazig University, Egypt. All the procedures were carried out according to the Local Experimental Animal Care Committee and approved by the Ethics of Institutional Committee of Department of Forensic Medicine and Toxicology, Faculty of Veterinary Medicine, Zagazig University, Egypt.
Chemicals: Endosulfan (Technicalgrade, 98%) was obtained from M/s Mitsu Crop Science Pvt. Ltd., India. All the other chemicals used were obtained from Merck, India and Sigma (St. Louis, MO, USA).
Birds and experimental diets: A total of one hundred eighty day-old mixed sexes Hubbard broiler chicks, purchased from a local hatchery, were weighed on arrival and randomly assigned to one of 6 treatments with three replicates of 10 chicks each based on a completely randomized design. The chicks were housed in conventional type cages with feed and water provided for ad libitum consumption and fed a diet formulated to meet nutrient requirements recommended commercially (Table 1).
| Table 1: | Ingredient and nutrient content of experimental diets |
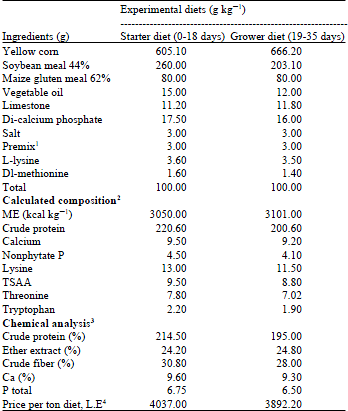 | |
1Provides per kg of diet: Vitamin A: 12,000 I.U, Vitamin D3: 5000 I.U, Vitamin E: 130.0 mg, Vitamin K3: 3.605 mg, Vitamin B1 (thiamin): 3.0 mg, Vitamin B2 (riboflavin), 8.0 mg, Vitamin B6, 4.950 mg, Vitamin B12: 17.0 mg, Niacin: 60.0 mg, D-Biotin: 200.0 mg, Calcium D-pantothenate: 18.333 mg, Folic acid: 2.083 mg, Manganese: 100.0 mg, Iron: 80.0 mg, Zinc: 80.0 mg, Copper: 8.0 mg, Iodine: 2.0 mg, Cobalt: 500.0 mg and Selenium: 150.0 mg. 2Calculated according to NRC (1994), 3Chemical analysis according to AOAC (2006), 4Caculated according to the price of feed ingredients when the experiment was started | |
Lighting condition was 23 h light and 1 h darkness. Vaccination and medical program were done according to the different stages of age under supervision of a veterinarian.
Six starter and grower diets were formulated to provide a similar nutrient profile with the exception of using additives or a combined addition of these additives. First group of chicks was fed basal diet, while the other five groups were fed basal diet supplemented with 5 g turmeric, 10 g turmeric, 30 mg endosulfan, 30 mg endosulfan plus 5 g turmeric and 30 mg endosulfan plus 10 g turmeric/kg diet, respectively, during the experimental period of five weeks. All the chicks were fed starter diets from 0-18 days of age and experimental grower diets from 19-35 days of age. The basal diet was analyzed for crude protein, ether extract, crude fiber, ash, calcium and phosphorus according to the procedures of the AOAC (2006).
Blood sample collection and tissue preparation: By the end of experimental period, 3 birds from each replicate were weighed, numbered and then slaughtered humanely. The blood samples were collected from the control and treated chicks where it was classified into 2 parts. First part allowed to clot overnight at room temperature, then centrifuged at 3,000 rpm for 10 min for separation of serum which was stored at -20°C for biochemical analysis, while the other part was collected in heparinized tubes and immediately used for evaluation of phagocytosis assay and micronucleus assessment.
Genotoxicity and cytotoxicity biomarkers
Micronucleus assay: Drops of whole blood from the experimental chicks of all the groups were directly smeared on slides. The slides were air-dried for 24 h, fixed in methanol for 10 min, followed by 10% Giemsa staining. To detect micronuclei in erythrocytes, the slides were analyzed using a 1000 oil-immersionlens. The mean frequency of micronuclei was evaluated per 1000 cells per group of chicks (Holden et al., 1997).
Estimation of serum 8-hydroxy-2-deoxyguanosine: The serum 8 OHdG level was detected using The Oxi Select_Oxidative DNA Damage ELISA Kit (Cell Biolabs, Inc., USA) according to Breton et al. (2003) method. The quantity of 8 OHdG in unknown sample is determined by comparing its absorbance with that of a known 8 OHdG standard curve. The kit has a detection sensitivity range of 100 pg mL-1 to 20 ng mL-1.
Lactate dehydrogenase (LDH) activity: The serum LDH activity was evaluated using a readymade reagent kit (Bio Med Diagnostic) according to the method described by Kachmar and Moss (1976).
DNA fragmentation assay: The DNA fragmentation assay was carried out on splenocytes of the chicks of each group using cellular DNA fragmentation Elisa kit (Cat. N0. 11 585 041 001) provided by Roche Diagnostics, Switzerland according to the manufacturer’s instructions.
Immunotoxicity biomarkers
Phagocytosis assay: The phagocytic activity was assessed according to Wilkinson (1977). The white blood cells were separated from peripheral blood of rat in the different groups. Heat-inactivated Candida albicans (C. albicans) was used to determine the phagocytic capacity. Macrophages containing C. albicans (phagocytosis%) as well as the number of C. albicans per one hundred phagocytes (phagocytic index) were counted.
Lysozyme activity assay: Serum lysozyme activity was measured using the turbidometric method as described by Ellis (1990). A 25 μL of each serum sample was added to the plate wells containing agarose gel diluted in 1% in phosphate buffer saline in which Micrococcus lysodeikticus cell had been dispersed. The diameter of clear zone formed around the wells after 24 h was measured. The lysozyme concentration levels were obtained from logarithmic curve using standard lysozyme.
Nitric oxide (NO) assay: The NO level in serum samples of the experimental chicks of all the groups were measured using the method described by Rajaraman et al. (1998) using the Griess reagent. Fifty microliters of each serum sample was incubated with an equal volume of Griess reagent in a 96 flat-bottomed microtiter plate well and incubated for 10 min at 27°C. After incubation, the optical density was recorded using an ELISA reader at wave length 570 nm. The nitrite concentration was calculated by using Na-nitrite standard curve.
Histopathological investigation: The spleen specimens were collected from chicks of different groups, weighed and fixed in 10% buffered neutral formalin solution. Five-micron thick paraffin sections were prepared, stained by hematoxylin and eosin for histopathological examination (Bancroft and Stevens, 1996).
Statistical analysis: The data generated in the present study was statistically analyzed using general linear models procedure adapted by SPSS for user’s guide with one-way ANOVA. Duncan test within program SPSS was done to determine the degree of significance between means.
RESULTS
In the present study, viability of birds during the whole experimental period as well as relative spleen weight (organ weight, g/100 g of body weight) of broilers at 35 days were statistically influenced by the dietary treatments. Where, birds fed on diet supplemented with 30 mg endosulfan/kg showed the lowest values for both viability and spleen weight among all the treatment groups. Conversely, the highest values were achieved by birds fed 5 g turmeric (Fig. 1a and b).
Genotoxicity and cytotoxicity biomarkers: The endosulfan-treated group of experimental chicks showed a significant increase in the frequency of micronucleus-containing erythrocytes (40.24), while this frequency significantly decreased (31 and 25) in endosulfan plus turmeric treatment on both the dose levels (5 and 10 g kg-1), respectively, to be near. The frequency reported in both turmeric (5 or 10 g) supplemented diet and control groups was 12.45, 11.82 and 8.56, respectively (Fig. 2a).
The 8 OHdG levels in the serum samples of the chicks were found to be significantly elevated in endosulfan fed group (76.97) compared with control group (59.02). While dietary supplementation of turmeric (5 and 10 g kg-1) protected DNA from oxidative damage either alone or in the presence of endosulfan in the diet, where addition of 5 g kg-1 of Curcuma longa to the diet containing endosulfan could decrease the level of 8 OHdG (68.46) but still higher than control, meanwhile 10 g kg-1 turmeric restored the level of 8 OHdG to the control value (Table 2).
Feeding chicks on the diet containing endosulfan markedly elevated the activity of LDH in serum (1056.95) compared with control and turmeric supplemented diets. Meanwhile, supplementation of diet with endosulfan in combination with turmeric at the two different added levels (5 and 10 g kg-1) significantly attenuated the activity of this enzyme when compared with endosulfan alone group (Table 2).
The percentage of DNA fragmentation was evaluated for all the treatment groups, where supplementation of basal diet with 5 or 10 g turmeric/kg decreased the DNA damage than control values to be 11 and 10%, respectively, instead of 15% for control group. Meanwhile, DNA fragmentation percentage was predominantly higher in splenocytes ofchicks fed on diet containing endosulfan (48%), supplementation of endosulfan diet with turmeric tended to decrease the DNA damage caused by endosulfan to be 28% in co-treatment by 5 g turmeric and 25% for endosulfan plus 10 g turmeric group (Fig. 2b).
Immunotoxicity biomarkers: The effect of endosulfan and turmeric exposure on immune status of the broiler chicks is illustrated in Fig. 3. Phagocytic percentag statistically decreased in chicks macrophages treated with endosulfan (60.70) when compared with control (85.17) and C. longa groups on both levels (86.21 and 86.94), respectively. Supplementation of turmeric to diet containing endosulfan showed significant increase in the phagocytic percent to be comparable with control group. The phagocytic index was significantly decreased in endosulfan fed group (0.71) with respect to control (1.20) and turmeric added groups (5 and 10 g kg-1) (1.18 and 1.22), respectively. Addition of turmeric at the two different levels to the diet containing endosulfan restored the phagocytic index to control level (Fig. 3a, b).
There was marked decrease in the serum lysozyme activity in the endosulfan fed chicks (87.69) compared to control and turmeric added groups at both levels. Co-treatment of endosulfan containing diet with turmeric (5 or 10 g kg-1) significantly increased the lysozyme activity to be 119.36 and 126.2, respectively but still lower than the frequency reported in control and both turmeric added groups.
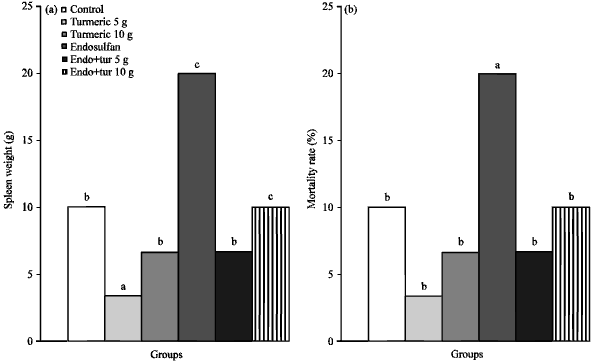 | |
| Fig. 1(a-b): | Effects of endosulfan (Endo) and feed added turmeric (Tur) on (a) Spleen weight and (b) Mortality rate. Each bars carrying different letters were significantly different (p<0.05) |
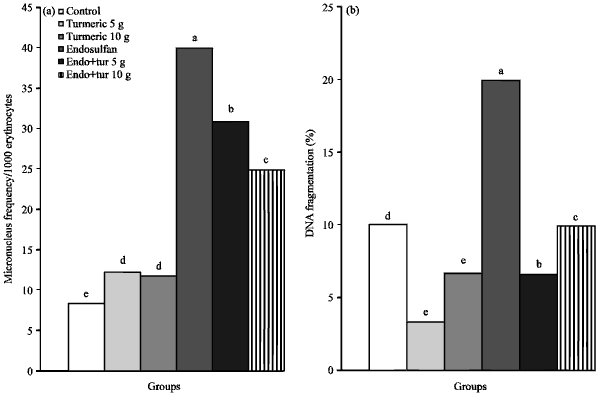 | |
| Fig. 2(a-b): | Effects of endosulfan (Endo) and feed added turmeric (Tur) on (a) Micronucleus frequency and (b) DNA fragmentation percentage. Each bars carrying different letters were significantly different (p<0.05) |
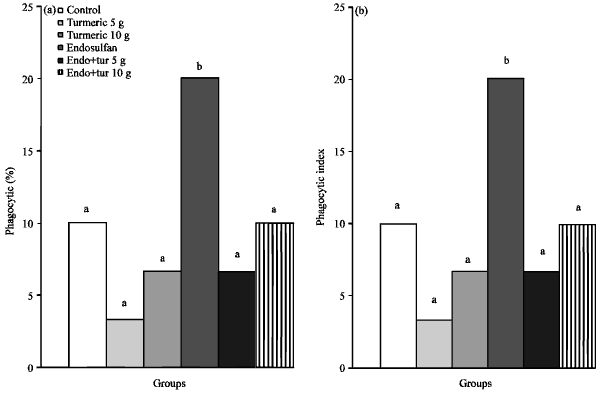 | |
| Fig. 3(a-b): | Effects of endosulfan (endo) and feed added turmeric (tur) on (a) Phagocytic (%) and (b) Phagocytic index. Each bars carrying different letters were significantly different (p<0.05) |
Also, endosulfan significantly stimulated NO production (2-fold increase with respect to control and turmeric added groups (p<0.01), whereas, supplementation of diet with endosulfan plus curcuma (5 or 10 g kg-1), significantly decreased NO level (76.15 and 63.95), respectively, but the both percents of turmeric in the presence of endosulfan could not restore the NO level to control value (Table 2).
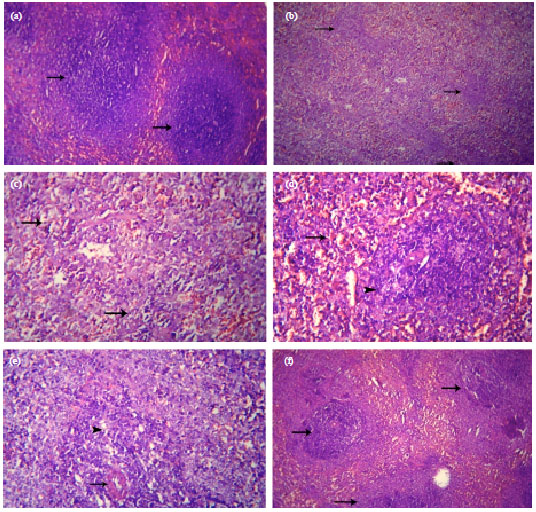 | |
| Fig. 4(a-f): | Spleen of birds from (a) Control and turmeric groups shows normal pattern and moderate hyperplasia in the lymphocytes of white pulp (arrow) (19x100), (b) Endosulfan received group shows severe depletion and necrosis in the lymphocytes of white pulp (arrows) (Bx 100 and Cx 400), (c) Endosulfan and turmeric (5 mg) group shows congestion (arrow) and moderate depletion in the lymphocytes of white pulp (arrowhead), (d) Hyaline degeneration in the wall of central arteriole (arrow) (e) Turmeric (10 mg) and endosulfan group shows normal white and red pulp (arrows) and (f) HEx400 |
| Table 2: | Effect of experimental diets on LDH and lysozyme, NO and 8 OHdG levels in serum of broilers at day 35 |
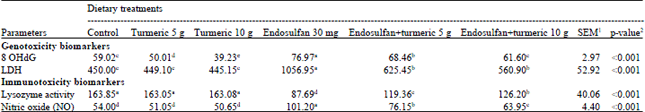 | |
| Values in the same row not sharing a common superscript differ significantly (p<0.05), 1SEM: Standard error mean, 2Overall treatment p-value | |
Histopathological investigation: The spleen of control or Curcuma longa treated birds showed normal pattern and architectures. Moderate hyperplasia in the lymphocytes of white pulp was detected in C. longa group (Fig. 4a). Endosulfan received group revealed severe depletion and necrosis in the lymphocytes of white pulp with some proliferation in the reticuloendothelial cells (Fig. 4b and c). Endosulfan and C. longa (5 g) group showed moderate depletion in the lymphocytes of white pulp (Fig. 4d) with severe congestion in the red pulp and hyaline degeneration in the wall of central arteriole (Fig. 4e). Endosulfan and C. longa (10 g) showed nearly normal white and red pulps (Fig. 4f).
DISCUSSION
In the present study, the ameliorative effect of feed added turmeric on broiler chicks exposed to endosulfan in diets was studied. Endosulfan predominantly increases the mortality rate than control and both turmeric groups in agreement with (Ganeshwade et al., 2012). This may be due to the severe physiological stress of endosulfan at cellular level.
The occurrence of micronucleus assay in erythrocytes (MN) represents an integrated response to chromosome-instability phenotypes and altered cellular viabilities caused by genetic defects and/or exogenous exposures to genotoxic agents (Iarmarcovai et al., 2008). The MN was used in this study as a marker of genotoxicity of the environmental pollutant, endosulfan. In the present experiment, broiler chicks exposed to 30 mg of endosulfan/kg diet showed a significant elevation of micronucleus frequency which comes on parallel with aquatic creatures results where elevated frequency of micronuclei was reported by Lajmanovich et al. (2005) and in Australoheros facetus (Crupkin et al., 2013) after exposure to endosulfan.
In the present study, DNA damage was assessed by evaluation of 8 OHdG level in serum and DNA fragmentation in spleen of birds of the different experimental groups. The significantly higher serum level of 8 OHdG and DNA fragmentation percentage in spleen observed in the chicks fed on diet supplemented with endosulfan revealed that the endosulfan is genotoxic.
The incidence of DNA damage was totally in agreement with the result of micronucleus assay of the present study, which indicates a possible relation between micronucleus formation and DNA damage which results in formation of smaller fragments of DNA, the cellular mechanism could form smaller nuclei with these fragments.
The DNA damage could be attributed to the induction of ROS under stress caused by exposure to endosulfan which attack the DNA and damage its integrity as endosulfan has been reported to induce oxidative tissue damage resulting from the release of Oxygen Free Radicals (OFRs), due to high reactivity of ORFs, most components of cellular structure and function may be become potential targets of oxidative damage. Moreover, these radicals can react with all biological macromolecules i.e., lipids, protein, nucleic acid and carbohydrates. The initial reaction generates a second radical, which reacts with a second macromolecule and oxidative damage in membrane phospholipids; pose a constant threat to cellular integrity and function. The lipid radicals are generated during the early encounter with an oxidant and oxygen to produce lipid dioxyl radical (Bartsch and Nair, 2000). The results of the present study came in harmony with previous data from in vitro and in vivo studies and showed that endosulfan is mutagenic and clastogenic and induce effects on cell cycle kinetics (Pandey et al., 1990). It has caused sex linked recessive mutations in Drosophila (Sharma et al., 2012), chromosomal aberrations in hamsters and dominant lethal mutations in mice (Naqvi and Vaishnavi, 1993). The genotoxicity of endosulfan was also demonstrated in HepG2 cells using sister chromatid exchanges, micronuclei and DNA strand breaks as detected by single-cell gel electrophoresis (Lu et al., 2000). The present results are also corroborated with the previous report who suggested that 8 OHdG has shown to be formed up on reaction of endosulfan with DNA (Karakaya et al., 1997).
The extent of cellular injury due to endosulfan toxicity in the present study was assessed by monitoring the value of LDH. Endosulfan significantly elevated LDH activity in serum which is considered as a presumptive marker of necrotic lesions. Cell necrosis leads to rise in LDH enzyme concentration in serum and tissue, provides an index of cell death and membrane permeability and an increase in its activity in the serum occurs as a result of cell membrane disintegration and enzyme leakage (Amin and Hamza, 2005). Thus, it is obvious that the increase of degenerative effects of endosulfan become more prominent with the increase in the LDH activity in serum.
Lipid peroxidative damage that occurs mainly in the cell membrane, caused by endosulfan may be the reason for increased release of LDH into the systemic circulation, where the earlier studies observed that endosulfan induces an elevation of lipid peroxides and ROS level in different tissues (Rastogi et al., 2014).
In the present study, turmeric was found to succeed in minimizing the cytotoxicity and genotoxicity of endosulfan to be comparable with control level. This may be due to that turmeric is a rich source of beneficial phenolic compounds: The curcuminoids, the three main curcuminoids having strong antioxidant activity being curcumin, demethoxycurcumin and bisdemethoxycurcumin (Balasubramanyam et al., 2003). Many studies have shown the capacity of curcumin, the most active component of turmeric, to prevent lipid peroxidation, a key process in the onset and progression of many diseases (Miquel et al., 2006). Moreover, Curcumin (CMN) reduces the oxidative stress in animals by its high ROS scavenging capacity and by protecting the antioxidant enzymes from being denaturated.
The presence of π-conjugation in CMN makes it more hydrophobic, which promotes its localization in the lipid bilayer membrane and lipid solubility. This allows the reaction of CMN with the lipid peroxyl radical and it acts as a chain terminating antioxidant (Sankar et al., 2010).
CMN minimize the risk of DNA fragmentation caused by lambda cyhalothrin in liver of rats (Madkour, 2012) and rat lymphocytes (Sharma et al., 2010). These results coincided with Siddique et al. (2010) who stated that CMN inhibits the generation of ROS that are responsible for DNA damage.
The results obtained in this study indicate that endosulfan can adversely affect immune parameters in broiler chicks. Significant decreases in lysozyme activity and phagocytic activity and increases in NO production were prominently observed. These alterations of immune system induced by endosulfan confirmed the previous finding of Aggarwal et al. (2009) on studying the DNA damaging effects in both blood lymphocytes and splenocytes in chicks fed endosulfan at 30 ppm in feed for a period of 60 days. The results indicated that endosulfan induces apoptosis in chicken peripheral blood lymphocytes and splenocytes as detected by DNA gel electrophoresis, which may be responsible for immunotoxicity induced by this chemical. Similar to the present observations on the effect of endosulfan on the activity of lymphocytes, significant depression in the functional activity of macrophages, assessed by nitrobluetetrazolium reduction test has been reported in chickens fed diet containing 30 ppm endosulfan (Khurana et al., 1998). The immunosuppressive effect of sublethal and subchronic concentrations of endosulfan was also detected in Swiss albino (Bhatia et al., 1998). However, Tellez-Banuelos et al. (2009) found that the effect of exposure to 7 ppb of endosulfan on phagocytic activity on tilapia spleen cells exhibited a significant increase in the percentage of actively phagocytosing cells and phagocytic index compared to the non-exposed control group.
Nitric oxide (iNOS), a gaseous free radical, is closely related to defence mechanisms against infectious organisms, it also regulates the functional activity, growth and death of many immune and inflammatory cell types including macrophages (Coleman, 2001). Therefore, iNOS generation was used in this study to investigate the effects of endosulfan on the functional integrity of macrophages. Earlier reports showed that treatment of mouse macrophages with endosulfan induces the expression of iNOS, Peroxynitrite formed from O2 and NO decreased lysozyme catalytic activity and enhanced its susceptibility to proteasomal degradation (Han et al., 2007; Curry-McCoy et al., 2009). Endosulfan also induces proinflammatory cytokines in mammals, suggesting the inflammatory potential of this insecticide. Moreover, a continuous production of these cytokines leads to an immune stimulation which could in turn trigger immune system deregulation including an autoimmune process (Descotes et al., 2000).
Concurrent treatment with turmeric significantly attenuated the alteration of immune system induced by endosulfan which may be attributed to the antioxidative and proinflamatory effect of the main component CMN. Numerous studies have confirmed the antioxidative and anti-inflammatory activities of CMN. The CMN mediates its protective effect by inducing the expression of the enzyme heme oxygenase-1 (HO-1) in vascular endothelial cells, this enzyme is very known well for its antioxidant potential (Kou et al., 2013). In vitro studies showed that CMN inhibits the translocation of proinflammatory and prooxidative transcription factors, namely, the nuclear factor kappa B (NF-κB) from cytoplasm to the nucleus (Aggarwal et al., 2006). The potential proinflammatory molecules, cyclooxygenase-2 (COX-2) and inducible nitric oxide synthesis (iNOS) have also been reported as potential targets of CMN in gastrointestinal epithelial cells (Zhang et al., 1999). Treatment of bacterial lipopolysaccharide (LPS) treated mice with CMN (92 μg kg-1 administered intragastrically) showed reduced hepatic iNOS mRNA expression to an extent of 50-70% (Chan et al., 1998).
It is postulated that endosulfan cause damage to lymphoid organs involved in immune responses and ultimately leads to immunosuppression, which can be explained by the reduction in splenic weight and the depletion of splenocytes observed in the present study in birds fed diet supplemented with 30 mg of endosulfan/kg. The reduction of splenic weight was confirmed by histopathological alterations in the spleen architecture, as manifested by severe depletion and necrosis in the lymphocytes of white pulp with some proliferation in the reticuloendothelial cells which supported the previous studies of Garg et al. (2004) in which, the relative splenic weight was significantly reduced in broiler chicks due to treatment of endosulfan and Varshneya et al. (1988) who reported necrotic and hyperplastic changes in lymphoid follicles of spleen during chronic endosulfan toxicity in white leghorn cockerels. And completely agreed with the opinion of Aggarwal et al. (2009) who stated that reduction in weights of the lymphoid organs can be indicative of the relative immune status, where a direct positive correlation was evident among reduction in the organ weights and a decrease in humoral and cell-mediated immune response in birds after 8 weeks of exposure to endosulfan.
Supplementation of diet containing endosulfan with turmeric ameliorate the immune suppressive effects of endosulfan through the protection against the lymphoid depletion, endosulfan and Curcuma longa (5 g) group showed decreased depletion in the lymphocytes of white pulp and hyperplasia of some white pulp. Endosulfan and C. longa (10 g) showed nearly normal white and red pulps.
CONCLUSION
The exposure of broiler chicks to endosulfan in their diets affects the viability of birds and induce genotoxic, cytotoxic, immunotoxic and histopathological effects. Supplementation of diet with turmeric produced appreciable ameliorative effects on these alterations offering a new approach in preventing oxidative stress deterioration induced by insecticides, however not restored fully to the control level. Thus, authors recommend that regular consumption of turmeric in the diet provides a constant supply of potential antioxidants that could counteract these sort of disorders. Further explorative studies are needed using several dose levels and durations of turmeric and incorporating other protective parameters which may give a full protection against endosulfan toxicity.
ACKNOWLEDGMENT
Authors thank grateful to Dr. Mohamed Hamed Mohamed, Professor of Pathology, Faculty of Veterinary Medicine, Zagazig University for his guidance and support in histopathological examination.
REFERENCES
- Aboul-Enein, A.M., I.N. Nasr, F.M.A. Elella and E.S. Abdullah, 2010. Monitoring of some organochlorines and organophosphorus residues in imported and locally raised chicken and bovine muscles in Egypt. J. Applied Sci. Res., 6: 600-608.
Direct Link - Aggarwal, S., H. Ichikawa, Y. Takada, S.K. Sandur, S. Shishodia and B.B. Aggarwal, 2006. Curcumin (diferuloylmethane) down-regulates expression of cell proliferation and antiapoptotic and metastatic gene products through suppression of IκBα kinase and Akt activation. Mol. Pharmacol., 69: 195-206.
CrossRefDirect Link - Aggarwal, M., S.B. Naraharisetti, S.N. Sarkar, G.S. Rao, G.H. Degen and J.K. Malik, 2009. Effects of subchronic coexposure to arsenic and endosulfan on the erythrocytes of broiler chickens: A biochemical study. Arch. Environ. Contam. Toxicol., 56: 139-148.
CrossRefDirect Link - Akbarian, A., A. Golian, H. Kermanshahi, A. Gilani and S. Moradi, 2012. Influence of turmeric rhizome and black pepper on blood constituents and performance of broiler chickens. Afr. J. Biotechnol., 11: 8606-8611.
Direct Link - Amin, A. and A.A. Hamza, 2005. Oxidative stress mediates drug-induced hepatotoxicity in rats: A possible role of DNA fragmentation. Toxicology, 208: 367-375.
CrossRefPubMedDirect Link - Balasubramanyam, M., A.A. Koteswari, R.S. Kumar, S.F. Monickaraj, J.U. Maheswari and V. Mohan, 2003. Curcumin-induced inhibition of cellular reactive oxygen species generation: Novel therapeutic implications. J. Biosci., 28: 715-721.
PubMed - Bancroft, J., A. Stevens and D. Turner, 1996. Theory and Practice of Histological Techniques. 4th Edn., Churchill, Livingston, New York, London, San Francisco, Tokyo Pages: 740.
CrossRefDirect Link - Breton, J., F. Sichel, F. Bianchini and V. Prevost, 2003. Measurement of 8-hydroxy-2'-deoxyguanosine by a commercially available ELISA test: Comparison with HPLC/electrochemical detection in calf thymus DNA and determination in human serum. Anal. Lett., 36: 123-134.
CrossRef - Chan, M.M.Y., H.I. Huang, M.R. Fenton and D. Fong, 1998. In vivo inhibition of nitric oxide synthase gene expression by curcumin, a cancer preventive natural product with anti-inflammatory properties. Biochem. Pharmacol., 55: 1955-1962.
CrossRefDirect Link - Coleman, J.W., 2001. Nitric oxide in immunity and inflammation. Int. Immunopharmacol., 1: 1397-1406.
CrossRef - Crupkin, A.C., P. Carriquiriborde, J. Mendieta, A.M. Panzeri, M.L. Ballesteros, K.S.B. Miglioranza and M.L. Menone, 2013. Oxidative stress and genotoxicity in the South American cichlid, Australoheros facetus, after short-term sublethal exposure to endosulfan. Pestic. Biochem. Physiol., 105: 102-110.
CrossRefDirect Link - Curry-McCoy, T.V., N.A. Osna and T.M. Donohue Jr., 2009. Modulation of lysozyme function and degradation after nitration with peroxynitrite. Biochimica Biophysica Acta (BBA)-Gen. Subj., 1790: 778-786.
CrossRefDirect Link - Descotes, J., G. Choquet-Kastylevsky, E. Van Ganse and T. Vial, 2000. Responses of the immune system to injury. Toxicol. Pathol., 28: 479-481.
CrossRef - Holden, H.E., J.B. Majeska and D. Studwell, 1997. A direct comparison of mouse and rat bone marrow and blood as target tissues in the micronucleus assay. Mutat. Res./Gen. Toxicol. Environ. Mutagen., 391: 87-89.
CrossRef - Hosseini-Vashan, S.J., A. Golian, A. Yaghobfar, A. Zarban, N. Afzali and P. Esmaeilinasab, 2012. Antioxidant status, immune system, blood metabolites and carcass characteristic of broiler chickens fed turmeric rhizome powder under heat stress. Afr. J. Biotechnol., 11: 16118-16125.
CrossRefDirect Link - Iarmarcovai, G., S. Bonassi, A. Botta, R.A. Baan and T. Orsiere, 2008. Genetic polymorphisms and micronucleus formation: A review of the literature. Mutat. Res./Rev. Mutat. Res., 658: 215-233.
CrossRefDirect Link - Khurana, S.K., R.S. Chauhan and S.K. Mahipal, 1998. Immunotoxic effects of cypermethrin and endosulfan on macrophage functions of broiler chicks. Indian J. Anim. Sci., 68: 105-106.
Direct Link - Kou, M.C., S.Y. Chiou, C.Y. Weng, L. Wang, C.T. Ho and M.J. Wu, 2013. Curcuminoids distinctly exhibit antioxidant activities and regulate expression of scavenger receptors and heme oxygenase-1. Mol. Nutr. Food Res., 57: 1598-1610.
CrossRef - Karakaya, A., P. Jaruga, V.A. Bohr, A.P. Grollman and M. Dizdaroglu, 1997. Kinetics of excision of purine lesions from DNA by Escherichia coli Fpg protein. Nucl. Acids Res., 25: 474-479.
CrossRefDirect Link - Lajmanovich, R.C., M. Cabagna, P.M. Peltzer, G.A. Stringhini and A.M. Attademo, 2005. Micronucleus induction in erythrocytes of the Hyla pulchella tadpoles (Amphibia: Hylidae) exposed to insecticide endosulfan. Mutat. Res./Gen. Toxicol. Environ. Mutagen., 587: 67-72.
CrossRef - Madkour, N.K., 2012. Protective effect of curcumin on oxidative stress and DNA fragmentation against lambda cyhalothrin-induced liver damage in rats. J. Applied Pharmaceut. Sci., 2: 76-81.
Direct Link - Tellez-Banuelos, M.C., A. Santerre, J. Casas-Solis, A. Bravo-Cuellar and G. Zaitseva, 2009. Oxidative stress in macrophages from spleen of Nile tilapia (Oreochromis niloticus) exposed to sublethal concentration of endosulfan. Fish Shellfish Immunol., 27: 105-111.
CrossRef - Mesa, M.D. and M.C. Ramirez-Tortosa, 2000. Nutritional and pharmacological effects of Curcuma longa L. extracts. Recent Res. Dev. Nutr., 3: 157-171.
Direct Link - Miquel, J., A. Ramirez-Bosca, J.V. Ramirez-Bosca and J.D. Alperi, 2006. Menopause: A review on the role of oxygen stress and favorable effects of dietary antioxidants. Arch. Gerontol. Geriatr., 42: 289-306.
CrossRefDirect Link - Naqvi, S.M. and C. Vaishnavi, 1993. Bioaccumulative potential and toxicity of endosulfan insecticide to non-target animals. Comp. Biochem. Physiol. Part C: Comp. Pharmacol., 105: 347-361.
CrossRefPubMedDirect Link - Nasr, I.N., M.H. Arief, A.H. Abdel-Aleem and F.M. Malhat, 2009. Persistent organic pollutants (pops) in Egyptian aquatic environment. J. Applied Sci. Res., 5: 1929-1940.
Direct Link - NRC., 1994. Nutrient Requirements of Poultry. 9th Edn., National Academy Press, Washington, DC., USA., ISBN-13: 9780309048927, Pages: 176.
Direct Link - Pandey, N., F. Gundevia, A.S. Prem and P.K. Ray, 1990. Studies on the genotoxicity of endosulfan, an organochlorine insecticide, in mammalian germ cells. Mutat. Res./Genet. Toxicol., 242: 1-7.
CrossRefDirect Link - Rajaraman, V., B.J. Nonnecke, S.T. Franklin, D.C. Hammell and R.L. Horst, 1998. Effect of vitamins A and E on nitric oxide production by blood mononuclear leukocytes from neonatal calves fed milk replacer. J. Dairy Sci., 81: 3278-3285.
CrossRefDirect Link - Rastogi, D., R. Narayan, D.K. Saxena and D.K. Chowdhuri, 2014. Endosulfan induced cell death in Sertoli-germ cells of male Wistar rat follows intrinsic mode of cell death. Chemosphere, 94: 104-115.
CrossRef - Sankar, P., A.G. Telang and A. Manimaran, 2010. Curcumin protects against cypermethrin-induced genotoxicity in rats. Environ. Toxicol. Pharmacol., 30: 289-291.
CrossRefPubMedDirect Link - Sharma, D.C., P.N. Saxena, V.K. Singh and R. Sharma, 2010. Assessment of DNA degradation in lymphocytes of albino rat (Rattus norvegicus) under lambda cyhalothrin stress. World Applied Sci. J., 11: 24-28.
Direct Link - Siddique, Y.H., G. Ara, T. Beg and M. Afzal, 2010. Protective effect of curcumin against chlormadinine acetate induced genotoxic damage in cultured human peripheral blood lymphocytes. Pharmacologyonline, 3: 644-650.
Direct Link - Vorkamp, K., F. Riget, M. Glasius, M. Pecseli, M. Lebeuf and D. Muir, 2004. Chlorobenzenes, chlorinated pesticides, coplanar chlorobiphenyls and other organochlorine compounds in Greenland biota. Sci. Total Environ., 331: 157-175.
CrossRef - Lu, Y., K. Morimoto, T. Takeshita, T. Takeuchi and T. Saito, 2000. Genotoxic effects of alpha-endosulfan and beta-endosulfan on human HepG2 cells. Environ. Health Perspect., 108: 559-561.
PubMedDirect Link - Zhang, F., N.K. Altorki, J.R. Mestre, K. Subbaramaiah and A.J. Dannenberg, 1999. Curcumin inhibits cyclooxygenase-2 transcription in bile acid- and phorbol ester-treated human gastrointestinal epithelial cells. Carcinogenesis, 20: 445-451.
CrossRefDirect Link - Sharma, M., J. Burre, P. Bronk, Y. Zhang, W. Xu and T.C. Sudhof, 2012. CSPα knockout causes neurodegeneration by impairing SNAP-25 function. EMBO J., 31: 829-841.
CrossRefPubMedDirect Link