
Claudia P. Sanchez-Olguin
Department of Physiology, National School of Biological Sciences,National Polytechnic Institute, Mexico
Sonia Guzman-Velazquez
Department of Physiology, National School of Biological Sciences,National Polytechnic Institute, Mexico
Angel I. Melo
Research Center in Animal Production, CINVESTAV-Laboratory Tlaxcala,Autonomous University of Tlaxcala, Tlaxcala City, Mexico
Gonzalo Flores
Neuropsychiatry Laboratory, Physiology Institute, Autonomous University of Puebla, Puebla City, Mexico
Fidel De-La-Cruz
Department of Physiology, National School of Biological Sciences,National Polytechnic Institute, Mexico
Sergio R. Zamudio
Department of Physiology, National School of Biological Sciences,National Polytechnic Institute, Mexico
International Journal of Pharmacology
Year: 2014 | Volume: 10 | Issue: 8 | Page No.: 406-417







ABSTRACT
Maternal separation disrupts mother-infant interactions during early life and produce long-lasting effects on physiological and behavioral processes in offspring. Maternal separation in rats can produce schizophrenia-like behaviors, such as stress hyper-reactivity, deficits in sensorimotor gating and disruption of cognitive processes and these behavioral dysfunctions can persist throughout adulthood. A neurotrophic peptide mixture, cerebrolysin, reduces behavioral and neuropathological alterations in a neurodevelopmental model of schizophrenia. This study examined the beneficial effects of sub-chronic cerebrolysin administration on long-lasting behavioral alterations caused by pre-weaning repeated maternal separation in rats. Cerebrolysin was administered daily for 4 weeks to 1 month old rats. The results showed that cerebrolysin treatment during post-pubertal age (2-3 months) reduced the hyper-reactivity to novel environment caused by maternal separation. Furthermore, this study also demonstrated that cerebrolysin administration partially reverted maternal separation-induced alterations in the acoustic startle response and its habituation process. These results suggest that cerebrolysin may exert beneficial effects in the management of some neuropsychiatric disorders.
PDF Abstract XML References Citation
Received: July 15, 2014;
Accepted: October 06, 2014;
Published: November 28, 2014
How to cite this article
Claudia P. Sanchez-Olguin, Sonia Guzman-Velazquez, Angel I. Melo, Gonzalo Flores, Fidel De-La-Cruz and Sergio R. Zamudio, 2014. Sub-Chronic Cerebrolysin Treatment Attenuates the Long-lasting Behavioral Alterations Caused by Maternal Separation in Rats. International Journal of Pharmacology, 10: 406-417.
DOI: 10.3923/ijp.2014.406.417
URL: https://scialert.net/abstract/?doi=ijp.2014.406.417
DOI: 10.3923/ijp.2014.406.417
URL: https://scialert.net/abstract/?doi=ijp.2014.406.417
INTRODUCTION
The offspring of altricial mammals are born in an immature state and maternal care affords a wealthy environment that shapes early physiological and cognitive development and later behaviors (Levine, 1957; Levy et al., 2003; Rees et al., 2005). The effects of deficient maternal care early in life on adult behavior were studied previously in rats, monkeys and humans (Fleming and Li, 2002; Maestripieri et al., 1997; O’Connor and Rutter, 2000). Maternal behaviors in rodents consist of a series of motor patterns performed by the mother, such as nest-building, retrieving, licking of young, adoption of a nursing posture and physiological functions, such as the production and ejection of milk (Gonzalez-Mariscal and Melo, 2013). Maternal Separation (MS) abates mother-pup interactions during early life and it produces long-lasting effects on physiological and behavioral processes in offspring (Fleming et al., 1999; Fleming and Li, 2002).
Pups are separated daily from the nest and mother during first two weeks of life in MS paradigms (Rees et al., 2005). Pups in some models are individually separated and only auditory and olfactory communications among siblings are possible (Monroy et al., 2010; Pascual and Zamora-Leon, 2007). The MS in rats produces schizophrenia like behaviors, such as stress hyper-reactivity, anxiety-like behaviors, anhedonia (Caldji et al., 1998; Daniels et al., 2004; Der-Avakian and Markou, 2010; Kikusui and Mori, 2009; Wigger and Neumann, 1999), disrupted habituation (Finamore and Port, 2000) and prepulse inhibition (PPI) of the acoustic startle response (Ellenbroek and Cools, 2000; Li et al., 2013). These behavioral dysfunctions persist throughout adulthood (Daniels et al., 2004; Macri and Wurbel, 2006). MS in rats also produces long-lasting morphological changes in pyramidal neurons of the hippocampus and prefrontal cortex (PFC) (Monroy et al., 2010; Pascual and Zamora-Leon, 2007) limbic structures related with the pathophysiology of schizophrenia (Glantz and Lewis, 1997; Kolomeets et al., 2005). This data suggest that MS can be used as an animal model of neuropsychiatric disease (Van den Buuse et al., 2003; Li et al., 2013).
Cerebrolysin (CBL) is a biologically active preparation of low molecular weight peptides that exert neuroprotective and neurotrophic properties (Veinbergs et al., 2000; Zhang et al., 2010). The beneficial effects of CBL administration are partially related to its ability to mimic the action of endogenous neurotrophic factors, such as Brain-Derived Neurotrophic Factor (BDNF) and Nerve Growth Factor (NGF) (Masliah and Diez-Tejedor, 2012). The CBL treatment may improve dendritic length and dendritic spine density in limbic regions of aged mice (Alcantara-Gonzalez et al., 2012; Juarez et al., 2011). The CBL also ameliorates synaptic and dendritic pathology in animal models of schizophrenia (Vazquez-Roque et al., 2012, 2014). These effects may underlie CBL use in the treatment of neurodegenerative diseases, such as multiple sclerosis, Parkinson’s disease, Alzheimer’s disease, dementia and acute or chronic stroke (Alvarez and Fuentes, 2011; Lukhanina et al., 2004; Masliah and Diez-Tejedor, 2012). However, CBL is also effective in the treatment of neurodevelopmental disorders, such as autism and schizophrenia (Flores and Atzori, 2014).
The present study examined the possible favorable effects of sub-chronic CBL administration on long-lasting behavioral alterations caused by a brief and repeated period of maternal separation in rats. Cerebrolysin was administered daily for 4 weeks to 1 month old rats. Behavioral analyses in post-pubertal animals included locomotor activity induced by novel environment, measures of sensorimotor function, such as acoustic startle response (ASR) and prepulse inhibition (PPI) and the relatively simple cognitive process of ASR habituation.
MATERIALS AND METHODS
Animals: Pregnant Sprague-Dawley rats were obtained from our facilities (Harlan Mexico was the original source) at gestational days 14-17. Rats were housed in Plexiglas cages with a stainless steel cover in a temperature (20-22°C) and humidity (50-60%) controlled room maintained on a 12 h light/dark cycle (lights on: 07:00-19:00 h) with free access to food and water. All animal procedures were performed in accordance with the Guide for Care and Use of Laboratory Animals’ of the Mexican Council for Animal Care (Norma Oficial Mexicana NOM-062-ZOO-1999) and the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Every effort was made to alleviate any pain or distress that might be experienced by the animals during this experiment.
Maternal separation: The day of birth was designated as Postnatal Day (PD) 0. Litters were culled to 8 male pups after birth and housed together with their mother in rat cages. MS protocol modified from Van den Buuse et al. (2003) was performed by removing pups from their home cage and individually isolating them for 1 h from PD 1-5 and 2 h from PD 6-14. Temperature was controlled (33±1°C from PD 1-9 and 22±1°C from PD 10-14) during the isolation period and only auditory and olfactory communications were accessible among siblings. Pups were returned to their home cages after isolation. Control pups were left undisturbed with their mothers in home cages. Maternal separation procedures were always performed in the same room at the same time of the day by the same person. Rats were weaned on PD 21 and housed (four animals per cage) with siblings from the same maternal care condition. Pup weights were recorded every two days from PD 4-12 and every four days throughout CBL treatment.
Cerebrolysin treatment: Four behavioral experiments were performed to assess possible beneficial effects of CBL on the behavior of post-pubertal MS rats: (1) Motor activity, (2) Acoustic startle response, (3) Prepulse inhibition of the acoustic startle response and (4) Habituation of the acoustic startle response. At PD 30, control (n =16) and MS (n =16) male rats were injected intraperitoneally daily for 28 days with 5 mL kg-1 of CBL (215.2 mg mL-1; Renacenz, Ever Neuro Pharma GmbH, Unterach, Austria) or vehicle (NaCl 0.9% solution). The CBL dose and treatment duration were chosen on the basis of previous findings (Rockenstein et al., 2006; Vazquez-Roque et al., 2012). Four groups of 8 animals each were formed: (1) Vehicle-treated control rats (CT-VEH), (2) CBL-treated control rats (CT-CBL), (3) Vehicle-treated MS rats (MS-VEH) and (4) CBL-treated MS rats (MS-CBL). Each group was formed with pups from at least three different litters of the same maternal care condition. Behavioral studies started five days after the end of CBL or VEH administration. Animals were submitted to open field motor activity on PD 63, ASR and PPI on PD70 and ASR habituation on PD78.
Behavioral tests
Open field motor activity: Animals were placed individually in the middle of an open field (black-painted wooden box 60x60x30 cm; wxlxh) on PD 63. Lighting conditions were similar to the light intensity in the housing room. Two 32-W overhead fluorescent bulbs were suspended 208 cm above the center of the field and provided 195 lux at the box floor. Spontaneous motor activity in an unfamiliar environment was measured using a real-time video image analyzer, Videomex-V (Columbus Instruments, OH, USA). This system keeps record of the distance the animal travels, the amount of time spent travelling, the amount of time spent in a non-ambulatory activity (which includes postural adjustments and stereotyped behaviors, such as grooming, rearing, sniffing, etc.) and the amount of time spent resting. A total of 9 min of activity for each animal was measured (Zamudio et al., 2005). The floor of the open field was wiped with a detergent solution between rats.
Acoustic startle response (ASR) and prepulse inhibition (PPI): Experiments were performed using a single automated Responder-X system (Columbus Instruments, Columbus, OH, USA). The startle chamber consisted of a polycarbonate plastic cage (15.4x28.1x17.5 cm; wxlxh) with a load-cell aluminum platform on the floor. This system uses unrestrained subjects and the load-cell platform records the force (g) of the startle reflex. Background noise (65 dB) and acoustic stimuli were provided via a loudspeaker placed 10 cm next to the test chamber. This system was kept within a sound-attenuating and ventilated cabinet. Stimuli were delivered and responses were measured using Responder-X software (Columbus Instruments, Columbus, OH, USA) on a PC. The startle response was taken as the first positive peak in force from calibrated zero. This measure is the sum of the subject’s mass and the startle response. We subtracted the subject’s mass (g) from the peak amplitude for all measurements of the startle response. Rats were placed in the startle chamber for a 5 min acclimation period with a 65-dB background noise. ASR and PPI were measured at PDs 70-73 in the same session, as reported previously (Ralph-Williams et al., 2003; Vazquez-Roque et al., 2012; Vinkers et al., 2007). Each experimental session consisted of baseline (BL) pulses, Non-Stimulus (NS) trials, startle trials (pulse-alone) and prepulse trials (prepulse+pulse). The background noise was present throughout the session. Six BL pulse trials of broadband noise (120 dB, 40 msec) were presented after the acclimation period to achieve a relatively stable level of startle response amplitude. Data of these BL trials was not considered in ASR and PPI analyses. Subsequently, a total of 80 trials of 8 different types were presented in pseudorandom order with a 30 sec inter-stimulus interval. Ten NS trials consisted of background noise only. Pulse-alone trials included 10 each of three different trials that consisted of 40 msec acoustic broadband noise pulses of 85, 105 and 120 dB. Four different prepulse+pulse trials consisted of a 40 msec noise prepulse (75 or 85 dB), a 100 msec delay and a 40 msec startle pulse (105 or 120 dB broadband noise). Therefore, the four prepulse+pulse trials were 10 each of 75-105, 85-105, 75-120 and 85-120 dB. The startle apparatus was wiped with a detergent solution between animals. Individual startle responses were calculated as the average of 10 same-type pulses or prepulse+pulse trials. PPI percentage was calculated as the difference between pulse-alone trials and the respective prepulse+pulse (same pulse intensity) trial divided by the pulse-alone trial multiplied by 100.
Habituation of the acoustic startle response: Experiments were performed using the single automated Responder-X system. This test was modified from Sasaki et al. (2001). Briefly, animals at PDs 78-81 were placed in the startle chamber for a 5 min acclimation period with a 65 dB background noise. The test session consisted of 150 consecutive startle pulses of 120 dB and 40 msec in duration with an inter-stimulus interval of 15 sec. The startle apparatus was cleaned with detergent solution between animals. ASR magnitudes were measured as described above.
Statistical analyses: Data of body weight during lactation was analyzed using a two-way analysis of variance (ANOVA) for repeated measures with maternal care condition as the between-subjects factor and time as the within-subjects factor. Mean body weight data during CBL treatment was analyzed using a three-way ANOVA for repeated measures with maternal care condition and CBL treatment as independent factors. Motor activity data was analyzed using a two-way ANOVA with maternal care condition and cerebrolysin treatment as independent factors. ASR and PPI data (within each pulse intensity) was analyzed using a two-way ANOVA with maternal care condition and CBL treatment as independent factors. Student-Newman-Keuls test was used when appropriate to make multiple, pair-wise group comparisons. Finally, linear regression analyses were performed for ASR habituation data. The level of significance was set at p<0.05. SigmaStat software version 3.5 (Systat Software Inc., San Jose, CA, USA) was used for all analyses.
RESULTS
Figure 1 shows the effect of the maternal care condition on body weight gain. Control and MS pups had the same body weight at PD 5 but a significant reduction in body weight gain in pups from the MS condition was observed from PD 10 until weaning (Fig. 1a). Notably, pups from the MS condition did not show differences in body weight compared to control pups when CBL treatment began (PD 30). Additionally, body weight gain values were not significantly different among groups during the drug treatment (Fig. 1b).
Figure 2 shows the spontaneous motor activity in an unfamiliar environment. ANOVA analyses revealed significant main effects of maternal care condition and CBL treatment on locomotion. Student-Newman-Keuls tests showed that post-pubertal vehicle-treated MS rats presented an increase in the distance traveled (Fig. 2a) and the amount of time spent travelling (Fig. 2b) compared to control groups. Notably, CBL treatment prevented the MS-induced increase in locomotion (Fig. 2a, b). In contrast, two-way ANOVA did not reveal significant differences caused by the maternal condition or CBL treatment on non-ambulatory and resting behaviors. Vehicle-treated MS rats showed similar mean values as control rats in the amount of time spent in non-ambulatory activity (Fig. 2c) and time in resting (Fig. 2d).
Two-way ANOVA revealed significant main effects of the maternal care condition and CBL treatment on acoustic startle magnitude (ASR). Student-Newman-Keuls tests demonstrated that post-pubertal vehicle-treated MS rats exhibited significant reductions in ASR at the pulse intensities of 105 and 120 dB (Fig. 3) compared to control groups. The CBL treatment attenuated these reductions in ASR but this effect was only significant at the 105-dB intensity. Small ASR amplitudes at a pulse intensity of 85 dB were found but two-way ANOVA did not reveal significant difference among groups (Fig. 3).
The effects of the maternal care condition and CBL treatment on the mean percentage of prepulse inhibition (PPI) in post-pubertal rats are shown in Fig. 4. Mean PPI values of approximately 40 and 60% were observed with prepulses of 75 and 85 dB. However, two-way ANOVA analyses did not reveal significant main effects caused by maternal separation or CBL treatment in any of the prepulse+pulse intensities tested (75-105, 85-105, 75-120 and 85-120 dB).
Figure 5 shows the effects of maternal care condition and CBL treatment on ASR habituation. The ASR magnitudes in control groups decreased throughout the trials. Linear regression analyses revealed significant correlations with negative slopes in these groups (CT-VEH: b = -1.000, r = 0.594, p<0.001 and CT-CBL: b = -0.995, r = 0.513, p<0.001).
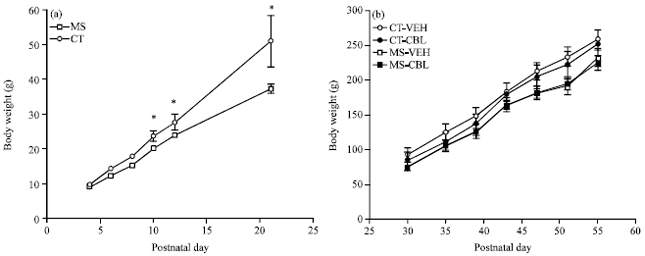 | |
| Fig. 1(a-b): | Effect of CBL treatment on body weight of rats with maternal separation, (a) Body weight gain in offspring from the Maternal Separation (MS) condition was lower than pups from the control group (CT). On PD 30, four groups were formed: vehicle-treated control rats (CT-VEH), cerebrolysin-treated control rats (CT-CBL), vehicle-treated MS rats (MS-VEH) and CBL-treated MS rats (MS-CBL). Throughout CBL treatment, (b) Body weight gain was similar in all groups. Values are means with standard errors represented by vertical bars. *Mean values were significantly different from the MS group (p<0.05; two-way repeated-measures ANOVA) |
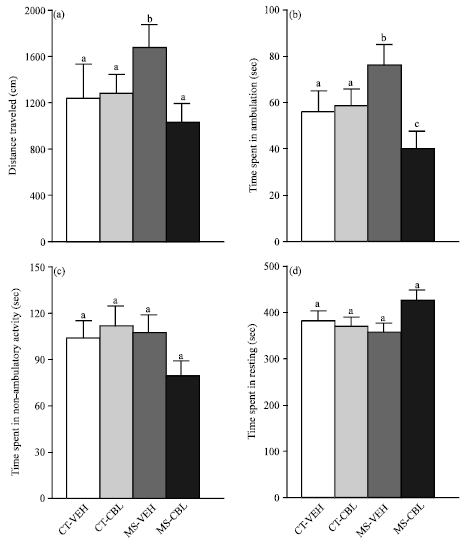 | |
| Fig. 2(a-d): | Effect of CBL treatment on motor activity in an unfamiliar open-field of rats with maternal separation. Vehicle-treated control rats (CT-VEH), cerebrolysin-treated control rats (CT-CBL), vehicle-treated maternal separation rats (MS-VEH) and CBL-treated MS rats (MS-CBL). Means±SEM of (a) Distance traveled, (b) Amount of time spent moving, (c) Amount of time spent in non-ambulatory activities and (d) Amount of time spent resting. Locomotor activity (a and b) was higher in MS-VEH rats than control groups, which was reduced by CBL treatment (MS-CBL group). Groups labeled with different letters are significantly different (p<0.05, two-way ANOVA) |
In contrast, no trend in ASR was observed in MS-VEH rats and there was no significant correlation (b = -0.100, r = 0.089, p = 0.279). CBL treatment partially restored that deficit in the habituation process; the MS-CBL group showed a significant correlation (r = 0.286; p<0.001) with a less negative slope (b = -0.401) than controls.
DISCUSSION
In present study, it was found that repeated maternal separation during the pre-weaning period (PD 1-14) induced long-lasting behavioral changes. Spontaneous ambulation in a novel environment was significantly higher in MS rats than control rats, MS also caused a reduction in the ASR and disruption of the habituation process but PPI was not altered. Notably, post-weaning sub-chronic CBL treatment attenuated the MS-induced behavioral alterations.
Body weight results must be considered before a discussion of the main findings of the present study. Early repeated maternal separation alters the quality of maternal care, which elicits several effects in the offspring (Pryce et al., 2001a). The first observed consequence is a change in body weight. It was found that body weight gain of MS pups was lower than control animals during the separation period and until weaning, which is consistent with previous studies using early repeated MS paradigms (Daoura et al., 2013; Zhang et al., 2013).
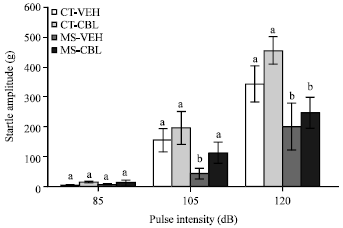 | |
| Fig. 3: | Effect of CBL treatment on the Acoustic Startle Response (ASR) in rats with maternal separation. Vehicle-treated control rats (CT-VEH), cerebrolysin-treated control rats (CT-CBL), vehicle-treated maternal separation rats (MS-VEH) and CBL-treated MS rats (MS-CBL). Data represent Means±SEM of the startle amplitude as measured in grams. ASR at 105 dB was lower in MS-VEH rats than control groups, which was reversed by CBL treatment (MS-CBL group). Groups labeled with different letters for each pulse intensity are significantly different (p<0.05, two-way ANOVA) |
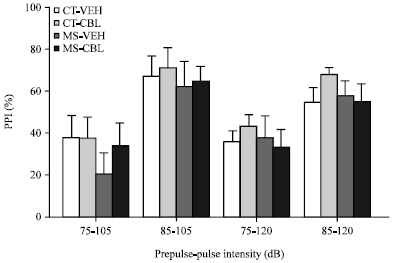 | |
| Fig. 4: | Effect of CBL treatment on prepulse inhibition (PPI) of the Acoustic Startle Response (ASR) in rats with maternal separation. Vehicle-treated control rats (CT-VEH), cerebrolysin-treated control rats (CT-CBL), vehicle-treated maternal separation rats (MS-VEH) and CBL-treated MS rats (MS-CBL). Data represent Means±SEM of the percentage of PPI. Two-way ANOVA analyses revealed no significant main effects of maternal care condition or CBL treatment |
All groups of rats had similar body weights after treatments (PD 58). This observation is consistent with results from Zhang et al. (2013) in which no differences in weights of male rats at PD 60 were observed between MS and control groups. Therefore, sub-chronic CBL (a mixture of low molecular weight peptides and free amino acids) administration does not appear to directly affect energy metabolism.
In general, adult offspring that were exposed to maternal separation in infancy display enhanced stress responsiveness and vulnerability for stress-related illness as adults (Faturi et al., 2010). Consistent with present data, previous studies (Pryce et al., 2001b; Slotten et al., 2006) showed that spontaneous ambulation in a novel environment was higher in MS rats than non-handled control rats. This behavioral abnormality is often related to increased mesolimbic-dopaminergic activity (Flores et al., 1996; Wan et al., 1996).
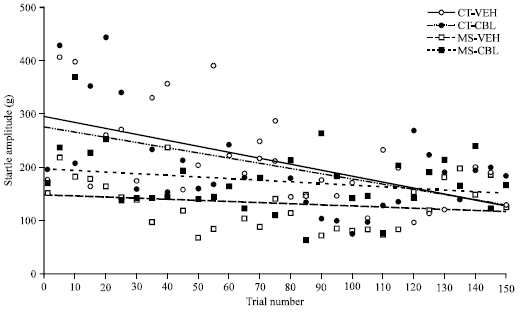 | |
| Fig. 5: | Effect of CBL treatment on the habituation of the Acoustic Startle Response (ASR) in rats with maternal separation. Vehicle-treated control rats (CT-VEH), cerebrolysin-treated control rats (CT-CBL), vehicle-treated maternal separation rats (MS-VEH) and CBL-treated MS rats (MS-CBL). Data represent the means of the startle amplitude as measured in grams (only data from each five trials are plotted). Linear regression analyses revealed a significant correlation with a negative slope in control groups (CT-VEH: b = -1.000, CT-CBL: b = -0.995). In contrast, no significant correlation was observed in MS-VEH rats. CBL treatment in MS rats produced a partial recovery of the habituation process (MS-CBL group showed a significant correlation with b = -0.401) |
Rats with a bilateral bilateral neonatal Ventral Hippocampus (nVH) lesion are a widely used neurodevelopmental animal model for the study of schizophrenia because these animals exhibit many schizophrenia-like behaviors (Tseng et al., 2009). Increased locomotor activity in a stressful novel environment is observed in animals with nVH lesions (Lipska et al., 1993). Sub-chronic treatment of animals with haloperidol or clozapine (dopaminergic antagonists) suppressed hyperlocomotion in this model (Lipska and Weinberger, 1994). A recent study (Vazquez-Roque et al., 2012) demonstrated that sub-chronic CBL treatment in rats with an nVH lesion ameliorated locomotor hyperresponsiveness and increased tyrosine hydroxylase expression in the nucleus accumbens (NAcc) shell. Notably, sub-chronic CBL treatment also attenuated the increase in locomotor activity elicited by MS in the present study. Taken together, this data suggest that MS or nVH lesion results in a post-pubertal development of mesolimbic-dopaminergic hyperactivity, which can be counteracted by sub-chronic CBL treatment.
Prepulse inhibition (PPI) of the Acoustic Startle Response (ASR) is a model of sensorimotor gating mechanisms in the brain. Deficits in PPI are observed in patients suffering from mental illnesses, such as schizophrenia (Braff et al., 2001). Rats with nVH lesions (a neurodevelopmental model of schizophrenia) also show disruption in PPI (Le Pen and Moreau, 2002; Vazquez-Roque et al., 2012). However, findings on the effects of MS on the development of adult PPI are controversial. Ellenbroek and Cools (2000), Ellenbroek et al. (2004) and Li et al. (2013) reported that MS diminishes PPI but other investigators (Finamore and Port, 2000; Lehmann et al., 2000; Pryce et al., 2001a; Van den Buuse et al., 2003) reported no effects of disruptions in maternal care on PPI. The effect of MS on PPI also varies based on rat strain (Varty and Geyer, 1998). The present study demonstrated that the postnatal experience of MS did not impair PPI in adulthood compared with normal maternal care (control group). These apparent differences may be due to methodological differences in the application of MS, including differences in the neonatal day of separation, duration of the separation, rat strain and PPI protocols. It was also demonstrated that CBL treatment did not alter PPI neither in MS or control rats.
The effects of MS on ASR received less attention. ASR is an unconditioned reflex of an organism to a sudden, relatively intense auditory stimulus. The response includes a contraction of the muscles of the body and limbs and often blinking. It was demonstrated that rats with MS had a decrement in the ASR compared to controls. This result is consistent with a previous report (Pryce et al., 2001b) in which the ASR to startle pulses was relatively high in non-handled subjects and relatively low in MS subjects. In contrast, Finamore and Port (2000) reported no differences in ASR caused by maternal care conditions. The mechanism of MS-induced changes in ASR in post-pubertal ages is undetermined. However, a relatively simple brainstem circuit mediates ASR. Acoustic stimuli are transmitted to the cochlear nuclei and signals are relayed from the cochlear nuclei to the caudal pontine reticular nucleus (PnC) then to cranial and spinal motor neurons (Koch, 1999). Projections from other brain areas modulate activity in the PnC, notably the central nucleus of the amygdala. The amygdala is the command center for enhancement of the ASR (fear-induced potentiation of the ASR) (Koch and Schnitzler, 1997), in addition the ventral hippocampus positively modulates amygdala activity via the bed nucleus of the stria terminalis (Lee and Davis, 1997). ASR magnitude is reduced when it is elicited in the presence of a conditioned stimulus that was previously paired with a primary reward (pleasure-attenuation of the ASR) (Schmid et al., 1995). Moreover, excitotoxic lesions of the nucleus accumbens (NAcc) prevent the attenuation of the ASR in presence of a rewarding stimulus (Koch et al., 1996). Therefore, MS may modify the ASR via neurodevelopmental alterations in these limbic structures. Support for this hypothesis comes from Monroy et al. (2010), who reported that MS decreased total dendritic length and dendritic spine density in ventral hippocampal pyramidal neurons and NAcc medium spiny neurons at a post-pubertal age. Also, anatomical-functional changes were demonstrated in the PFC of MS rats (Baudin et al., 2012; Monroy et al., 2010; Pascual and Zamora-Leon, 2007). The PFC regulates diverse neural processes through connections with several structures, such as the hippocampus, amygdala and NAcc (Arnsten et al., 2012). The alterations in neuromorphology and interconnectivity of the PFC, NAcc and hippocampus can account for the effects of MS on ASR but alterations in other brain regions (e.g., PnC, cochlear nuclei and lateral central gray) cannot be excluded. Notably, sub-chronic CBL administration to MS rats partially counteracted the reduction in ASR at post-pubertal ages, which suggests that CBL promotes a recovery of dendritic damage to NAcc, PFC and hippocampal neurons. This assumption is supported by nHV lesion experiments in rats (neurodevelopmental model of schizophrenia). Neuropathological analyses of these limbic regions showed that post-weaning sub-chronic CBL treatment of rats with an nVH lesion ameliorated behavioral alterations, dendritic pathology and neuronal loss in the PFC, basolateral amygdala and NAcc (Vazquez-Roque et al., 2012, 2014).
Several studies showed impairments in cognitive tasks, such as the Morris water maze and novel object recognition, in adult MS rats (Aisa et al., 2007; Benetti et al., 2009; Hui et al., 2011; Huot et al., 2002). In fact, impairments in a simple cognitive process, such as habituation, were also described in MS rats. Finamore and Port (2000) and Pryce et al. (2001b) showed an altered habituation of the ASR caused by MS, which is consistent with our results. It was found that MS interrupted the process of habituation. Controls rats showed a decrement in the amplitude of ASR during trials but MS rats failed to present reductions in startle due to repeated stimulation. However, post-weaning sub-chronic CBL administration in this experiment only partially affected the recovery of habituation in MS rats. It is difficult to explain this partial improvement because the neural mechanism underlying ASR habituation is not clear. This work and the above mentioned studies used paradigms of short-term habituation in which the decline of the startle response amplitude after repeated stimulation occurred within a single test session (Koch and Schnitzler, 1997). This type of habituation may occur within the basic ASR circuitry and not through the participation of more rostral brain structures (Leaton et al., 1985) but likely at the connection between cochlear nuclei neurons with PnC neurons (Pilz and Schnitzler, 1996). The recovery of dendritic neuronal damage mediated by CBL treatment was described primarily in forebrain areas (Alcantara-Gonzalez et al., 2012; Vazquez-Roque et al., 2012, 2014) and there are no reports on the effects of CBL on more caudal brain regions, such as PnC. Therefore, a reduced sensitivity of PnC neurons to CBL could explain the partial recovery of the habituation process in CBL-treated MS rats.
The effects of CBL on the amelioration of behavioral alterations in adult MS rats may be related to its capacity to promote dendritic regeneration and/or neuroprotection. Neurotrophic effects of CBL on embryonic chicken neurons were demonstrated (Satou et al., 2000) and CBL may also indirectly increase neurotrophins, such as BDNF, NGF, ciliary neurotrophic factor (CNTF) and Glial-derived Neurotrophic Factor (GDNF) (Chen et al., 2007; Menon et al., 2012; Ubhi et al., 2013). The neurotrophic action mediated by CBL interferes with excitotoxicity, free radical formation and inflammatory responses (Gonzalez et al., 1998; Hutter-Paier et al., 1996; Veinbergs et al., 2000). However, further studies should be conducted to determine the neurobiological mechanisms of CBL actions in MS rats.
CONCLUSION
In conclusion, a neurotrophic agent, CBL, reduces behavioral alterations after puberty in rats that suffered maternal separation. These results support the beneficial effects of CBL in the management of some neuropsychiatric disorders.
ACKNOWLEDGMENT
SRZ, FDC, AIM and GF, acknowledge the National Research System of Mexico for membership, CPS acknowledges the CONACYT for scholarship. Thanks to American Journal Experts (CVK: EB92-FDB0-C4DE-E8AC-5C9) for editing the English language text. Funding for this study was provided in part by grant from SIP-IPN 20130757 to S. Zamudio.
REFERENCES
- Alvarez, X.A. and P. Fuentes, 2011. Cerebrolysin in Alzheimer's disease. Drugs Today, 47: 487-513.
CrossRef - Arnsten, A.F.T., M.J. Wang and C.D. Paspalas, 2012. Neuromodulation of thought: Flexibilities and vulnerabilities in prefrontal cortical network synapses. Neuron, 76: 223-239.
CrossRef - Baudin, A., K. Blot, C. Verney, L. Estevez and J. Santamaria et al., 2012. Maternal deprivation induces deficits in temporal memory and cognitive flexibility and exaggerates synaptic plasticity in the rat medial prefrontal cortex. Neurobiol. Learn. Memory, 98: 207-214.
CrossRef - Benetti, F., P.B. Mello, J.S. Bonini, S. Monteiro, M. Cammarota and I. Izquierdo, 2009. Early postnatal maternal deprivation in rats induces memory deficits in adult life that can be reversed by donepezil and galantamine. Int. J. Dev. Neurosci., 27: 59-64.
CrossRefDirect Link - Braff, D.L., M.A. Geyer and N.R. Swerdlow, 2001. Human studies of prepulse inhibition of startle: Normal subjects, patient groups and pharmacological studies. Psychopharmacology, 156: 234-258.
CrossRefDirect Link - Caldji, C., B. Tannenbaum, S. Sharma, D. Francis, P.M. Plotsky and M.J. Meaney, 1998. Maternal care during infancy regulates the development of neural systems mediating the expression of fearfulness in the rat. Proc. Natl. Acad. Sci. USA., 95: 5335-5340.
PubMed - Chen, H., Y.C. Tung, B. Li, K. Iqbal and I. Grundke-Iqbal, 2007. Trophic factors counteract elevated FGF-2-induced inhibition of adult neurogenesis. Neurobiol. Aging, 28: 1148-1162.
CrossRef - Daniels, W.M.U., C.Y. Pietersen, M.E. Carstens and D.J. Stein, 2004. Maternal separation in rats leads to anxiety-like behavior and a blunted ACTH response and altered neurotransmitter levels in response to a subsequent stressor. Metab. Brain Dis., 19: 3-4.
CrossRefDirect Link - Daoura, L., I. Nylander and E. Roman, 2013. Qualitative differences in pup-retrieval strategies in a maternal separation paradigm. J. Behav. Brain Sci., 3: 603-616.
CrossRefDirect Link - Der-Avakian, A. and A. Markou, 2010. Neonatal maternal separation exacerbates the reward-enhancing effect of acute amphetamine administration and the anhedonic effect of repeated social defeat in adult rats. Neuroscience, 170: 1189-1198.
CrossRefDirect Link - Ellenbroek, B.A., N.M.W.J. de Bruin, P.T.J.M. van Den Kroonenburg, E.L.J.M. van Luijtelaar and A.R. Cools, 2004. The effects of early maternal deprivation on auditory information processing in adult Wistar rats. Biol. Psychiatry, 55: 701-707.
Direct Link - Ellenbroek, B.A. and A.R. Cools, 2000. The long-term effects of maternal deprivation depend on the genetic background. Neuropsychopharmacology, 23: 99-106.
CrossRefDirect Link - Faturi, C.B., P.A Tiba, S.E. Kawakami, B. Catallani, M. Kerstens and D. Suchecki, 2010. Disruptions of the mother-infant relationship and stress-related behaviours: Altered corticosterone secretion does not explain everything. Neurosci. Biobehav., Rev. 34: 821-834.
CrossRefDirect Link - Finamore, T.L. and R.L. Port, 2000. Developmental stress disrupts habituation but spares prepulse inhibition in young rats. Physiol. Behav., 69: 527-530.
CrossRef - Flores, G. and M. Atzori, 2014. The potential of cerebrolysin in the treatment of Schizophrenia. Pharmacol. Pharm., 5: 691-704.
CrossRefDirect Link - Flores, G., G.K. Wood, J.J. Liang, R. Quirion and L.K. Srivastava, 1996. Enhanced amphetamine sensitivity and increased expression of dopamine D2 receptors in postpubertal rats after neonatal excitotoxic lesions of the medial prefrontal cortex. J. Neurosci., 16: 7366-7375.
PubMed - Glantz, L.A. and D.A. Lewis, 1997. Reduction of synaptophysin immunoreactivity in the prefrontal cortex of subjects with schizophrenia. Arch. Gen. Psychiatry, 54: 660-669.
CrossRefPubMedDirect Link - Gonzalez, M.E., L. Francis and O. Castellano, 1998. Antioxidant systemic effect of short-term Cerebrolysin® administration. J. Neural Transm. Suppl., 53: 333-341.
CrossRefPubMedDirect Link - Hui, J.J., Z.J. Zhang, S.S. Liu, G.J. Xi and X.R. Zhang et al., 2011. Hippocampal neurochemistry is involved in the behavioural effects of neonatal maternal separation and their reversal by post-weaning environmental enrichment: A magnetic resonance study. Behav. Brain Res., 217: 122-127.
CrossRefDirect Link - Huot, R.L., P.M. R.H. Lenox and R.K. McNamara, 2002. Neonatal maternal separation reduces hippocampal mossy fiber density in adult long evans rats. Brain Res., 950: 52-63.
CrossRef - Hutter-Paier, B., E. Grygar and M. Windisch, 1996. Death of cultured telencephalon neurons induced by glutamate is reduced by the peptide derivative Cerebrolysin. J. Neural Transm. Suppl., 47: 267-273.
Direct Link - Juarez, I., D.J. Gonzalez, R. Mena and G. Flores, 2011. The chronic administration of cerebrolysin induces plastic changes in the prefrontal cortex and dentate gyrus in aged mice. Synapse, 65: 1128-1135.
CrossRefDirect Link - Koch, M., A. Schmid and H.U. Schnitzler, 1996. Pleasure attenuation of startle is disrupted by lesions of the nucleus accumbens. Neuroreport, 7: 1442-1446.
PubMed - Le Pen, G. and J.L. Moreau, 2002. Disruption of prepulse inhibition of startle reflex in a neurodevelopmental model of schizophrenia: Reversal by clozapine, olanzapine and risperidone but not by haloperidol. Neuropsychopharmacology, 27: 1-11.
CrossRefDirect Link - Lee, Y. and M. Davis, 1997. Role of the hippocampus, the bed nucleus of the stria terminalis and the amygdala in the excitatory effect of corticotropin-releasing hormone on the acoustic startle reflex. J. Neurosci., 17: 6434-6446.
PubMed - Levine, S., 1957. Infantile experience and resistance to physiological stress. Science, 126: 405-405.
CrossRefPubMedDirect Link - Lukhanina, E.P., I.N. Karaban, I.A. Burenok, N.A. Mel'nik and N.M. Berezetskaia, 2004. [Effect of cerebrolysin on the electroencephalographic indices of brain activity in Parkinson's disease]. Zh. Nevrol. Psikhiatr. Im. S.S. Korsakova, 104: 54-60, (In Russian).
PubMed - Masliah, E. and E. Diez-Tejedor, 2012. The pharmacology of neurotrophic treatment with cerebrolysin: Brain protection and repair to counteract pathologies of acute and chronic neurological disorders. Drugs Today, 48: 3-24.
PubMed - Pascual, R. and S.P. Zamora-Leon, 2007. Effects of neonatal maternal deprivation and postweaning environmental complexity on dendritic morphology of prefrontal pyramidal neurons in the rat. Acta Neurobiol. Exp., 67: 471-479.
PubMed - Sasaki, A., W.C. Wetsel, R.M. Rodriguiz and W.H. Meck, 2001. Timing of the acoustic startle response in mice: Habituation and dishabituation as a function of the interstimulus interval. Int. J. Comput. Psychol., 14: 258-268.
Direct Link - Vazquez-Roque, R.A., B. Ramos, C. Tecuatl, I. Juarez and A. Adame et al., 2012. Chronic administration of the neurotrophic agent cerebrolysin ameliorates the behavioral and morphological changes induced by neonatal ventral hippocampus lesion in a rat model of schizophrenia. J. Neurosci. Res., 90: 288-306.
CrossRefPubMed - Veinbergs, I., M. Mante, M. Mallory and E. Masliah, 2000. Neurotrophic effects of cerebrolysin in animal models of excitotoxicity. J. Neural. Transm. Suppl., 59: 273-280.
PubMed