
M. Yaqoob Wani
Immunology Section,Indian Veterinary Research Institute, Izatnagar,Bareilly, 243122, Uttar Pradesh, India
Tapas Kumar Goswami
Immunology Section,Indian Veterinary Research Institute, Izatnagar,Bareilly, 243122, Uttar Pradesh, India
Divya Chaudhary
Immunology Section,Indian Veterinary Research Institute, Izatnagar,Bareilly, 243122, Uttar Pradesh, India
Rajendra Singh
Division of Pathology, Indian Veterinary Research Institute, Izatnagar,Bareilly, 243122, Uttar Pradesh, India
Kuldeep Dhama
Division of Pathology, Indian Veterinary Research Institute, Izatnagar,Bareilly, 243122, Uttar Pradesh, India
International Journal of Pharmacology
Year: 2014 | Volume: 10 | Issue: 7 | Page No.: 389-397







ABSTRACT
Lipopolysaccharide is the most important component of Gram negative bacteria that activates immune system through TLR-4 pathway. In the present study, the adjuvant effects of Pasturella multocida 52 strain (Pm52) on Hen Egg-white Lysozyme (HEL) and the possible attenuation of septic signs by pravastatin were investigated. In vitro results showed that Pm52 LPS stimulates nitric oxide (NO) production from splenocytes in time and dose dependent manner where as pravastatin (10 μM) inhibited the LPS induced NO production to basal levels (p<0.05). Furthermore, presence of pravastatin (10 μM) does not inhibit the in vitro proliferative responses of splenocytes by graded doses of Pm52 extracted LPS. The co-administration of Pm52 LPS with HEL by subcutaneous route in mice significantly increased TNF-α and NO levels at 2 and 6 h, respectively, as well as the HEL specific IgG development at 14th day Post Inoculation (PI). Administration of pravastatin by intraperitoneal route significantly reduced serum TNF-α and NO levels, the pro-inflammatory mediators responsible for induction of septic shock as well as stabilized TNF-α: IL-10 ratio without affecting HEL specific adaptive immune responses. However, further studies are required in this direction to determine the long term immune modulating effects of LPS and pravastatin on the host animals so as to be used in a beneficial way.
PDF Abstract XML References Citation
Received: May 26, 2014;
Accepted: September 11, 2014;
Published: October 31, 2014
How to cite this article
M. Yaqoob Wani, Tapas Kumar Goswami, Divya Chaudhary, Rajendra Singh and Kuldeep Dhama, 2014. Mitigation of Septic Signs by Pravastatin during LPS Co-Administered Hen-Egg White Lysozyme Immunization in Mice. International Journal of Pharmacology, 10: 389-397.
DOI: 10.3923/ijp.2014.389.397
URL: https://scialert.net/abstract/?doi=ijp.2014.389.397
DOI: 10.3923/ijp.2014.389.397
URL: https://scialert.net/abstract/?doi=ijp.2014.389.397
INTRODUCTION
The Pathogen Associated Molecular Patterns (PAMPs) are the conserved microbial motifs that are recognized by innate Pathogen Recognition Receptors (PRRs) (Mogensen, 2009). The PRRs are regarded as essential and integral part of the normal early embryonic development and have important roles in wide variety of interactions (Strunk et al., 2011). The PRRs help not only in early, specific and balanced immune recognition but also in the development of effective adaptive immune response to different antigenic responses (Akira et al., 2001; Barton and Medzhitov, 2002). The various PAMPs recognized by PRRs include bacterial Lipopolysaccharide (LPS), flagellin protein, peptidoglycans, double stranded RNA molecules and ummethylated DNA (CpG) motifs (Kawai and Akira, 2011). Among them LPS is one of the early recognized immune stimulants and is the major structural constituent of Gram negative bacteria, constitutes 1-10% of bacterial dry weight (Hodgson, 2006).
The biological mechanisms of LPS recognition are dynamic involving stimulation and/or inhibition of complex mechanisms and thus indicate it to be an exogenous hormone than a toxin (Marshall, 2005). Due to mimicking inflammatory effects, LPS is often used as model of inflammation than innate immunity (Mitchell et al., 2007). However, its role as vaccine adjuvant cannot be ruled out due tissue damage or toxicity as induced by inflammatory cytokines which is a necessary prerequisite for an effective immune response as per Matzinger danger hypothesis (Matzinger, 2002). Individual cytokines or their combinations have been tried as adjuvants in several vaccine formulations. However, LPS has several advantages over cytokines: it is inexpensive and not restricted to species differences; it generally produces a more robust response than individual cytokines (Mitchell et al., 2007). Ideally a vaccine adjuvant should strongly stimulate B-and T-cell immunity while avoiding the excess innate immune system activation and inflammatory cytokine production that mediates adjuvant reactogenicity and toxicity (Petrovsky, 2008). The LPS acts not only as a potent B-cell mitogen but also activates T-cells to produce IFN-γ and TNF-α, thereby enhancing cellular immunity (Petrovsky and Aguilar, 2004). The main difficulty of LPS hindering its FDA approval is the acute nature of sepsis and septic shock induction.
Statins are widely and extensively used as the most effective drugs for lowering cardiovascular risk associated with elevated Low Density Lipid (LDL) cholesterol (Brown and Goldstein, 1986; Gao et al., 2008). There are several lines of evidence that suggest the beneficial effects associated with statin treatment and are mediated by LDL independent (pleiotropic) mechanisms. The various pleiotropic effects include anti-inflammatory actions, improvement of endothelial and microvascular functions, modulation of endothelial nitric oxide synthase (eNOS) (Lefer, 2002; Greenwood et al., 2006). Moreover, studies have confirmed that statins decrease the acute inflammatory substances during septic condition (Ando et al., 2000; McGown et al., 2010; Wani et al., 2013; Yeo et al., 2013). However, there are chances that the extensive and long term use of statins may have many adverse effects on the host itself. In this regard the present study was undertaken to study the effects of pravastatin on the LPS co-administered weak immunogen to monitor the immune stimulation and its concurrent effects in mice. The results indicated that hydrophilic pravastatin was effective in reducing the LPS induced septic condition without effecting its long term beneficial of LPS immune adjuvant effects.
MATERIALS AND METHODS
Bacteria: Freeze-dried ampoule of the P. multocida strain 52 (Pm52) of serotype B:2 used in this study was procured from the Division of Bacteriology and Mycology of Indian Veterinary Research Institute (IVRI), Izatnagar. Culture was revived in Brain Heart Infusion (BHI) broth, followed by identification on the basis of morphology and biochemical characters as described previously (Mir et al., 2012).
Experimental animals: Swiss Albino mice aged 6-8 weeks and weighing about 25-30 g were obtained from the Laboratory Animal Resource (LAR) section of the IVRI, Izatnagar. Animals were used after an acclimatization period of 4-5 days. All experimental procedures on animals were carried out according to the recommendations and approval of the Institute Animal Ethics Committee (IAEC) under the guidelines set forth by the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), Ministry of Environment and Forestry, Government of India.
LPS isolation characterization and quantification: The LPS from the P. multocida Pm52 cell wall was isolated by hot water/phenol (Westphal and Jann, 1965), characterized by silver staining method (Tsai and Frasch, 1982) and quantified indirectly by measuring the carbohydrate content of LPS using the phenol-sulphuric acid method (Masuko et al., 2005) with some modifications.
Splenocytes culture: Mice were humanely killed with chloroform anaesthesia; spleens were collected aseptically and single cell suspensions were obtained by repeated perfusion of spleen using Phosphate Buffer Saline (PBS) as described previously (Wani et al., 2011). Splenocytes were finally resuspended in complete RPMI 1640 complete growth medium (containing 10% FCS with sodium bicarbonate, vitamins and streptomycin and penicillin), enumerated and plated in 96 well culture plates.
Effect of pravastatin on Pm52 LPS stimulated splenocytes: To determine the effect of LPS treatments on Nitric Oxide (NO) production and the lymphocyte proliferation, splenocytes were seeded in the 96 well culture plate at concentration of 2x105 cells/well and stimulated with different doses of Pm52 derived LPS (2 and 4 μg mL-1) either in presence or absence of pravastatin (10 μM). Nitric oxide production was measured indirectly by determining the concentrations of nitrite levels in the culture supernatant at 48 and 72 h post stimulation using Greiss reagent as described earlier (Wani et al., 2011). For determining the effect of LPS and pravastatin treatments on the proliferation of splenocytes were stimulated with graded doses of Pm52 derived LPS (1, 2 and 4 μg mL-1), incubated for 96 h and treated with MMT dye as per the standard method (Mosmann, 1983).
Immunization trial: A total of 24 healthy female mice were randomly distributed into four groups, caged separately and immunized as describe in Table 1. Serum from the blood collected from the retro orbital plexus of the eye at various Post Inoculation (PI) intervals from various animals was collected and analyzed for the serum cytokines, total nitric oxide (nitrite and nitrate levels: NOx) and anti-HEL specific IgG levels.
Quantification of serum cytokines and total nitric oxide: Serum was obtained from the blood collected at 0, 2, 6, 12 and 24 h PI and stored at -20°C till used. The serum levels of TNF-α and IL-10 cytokines were determined using commercially available ELISA kits (Genetix Biotech). Total serum nitrite (NOx) concentrations were determined using modified Greiss reaction (Sastry et al., 2002).
| Table 1: | Immunization protocol for various groups |
 | |
| †Hen egg white lysozyme (HEL) was inoculated through subcutaneous route and the inoculums were prepared in 100 μL of sterile phosphate buffer saline, ‡Dose for HEL and LPS was 10 μg each when ever inoculated, δPravastatin was administered through intraperitoneal route at the dose rate of 0.02 μg kg-1 b.wt. | |
The method is based on the reduction of nitrate to nitrite by means of copper cadmium alloy, followed by its quantification using the Greiss reagent.
Determination of humoral immune responses: To evaluate the humoral immune response, serum samples were collected at 0, 7th, 14th, 21st and 28th day PI of mice. Indirect-ELISA for anti-HEL specific total IgG was standardized using standard positive and negative sera of the mice. Optimum concentrations/dilutions for antigen, antibody and conjugate were determined using the checker board titration method to achieve acceptable positive/negative ratio. Briefly, 96 well ELISA plate (Nunc-Immunoplate with Maxisorp Surface) were coated with 100 μL of HEL (16 μg) in bicarbonate buffer. Test sera samples (diluted 1:100) were added (100 μL/well) in triplicate wells and after necessary washing steps goat anti-mouse IgG horseradish peroxidase (HRPO) conjugate (100 μL/well at 1:10,000 dilutions; Sigma Aldrich, USA) was added. After usual washing steps, the plates were finally developed by adding 100 μL/well of freshly prepared substrate solution (10 mg of O-Phenylenediaminedihydrochloride, 2 μL of Hydrogen peroxide in citrate buffer; pH 4.6) and incubated for 15 min in the dark. The reaction was stopped by addition of 50 μL of 1N H2SO4 and optical density was measured at 490 nm using a microplate reader (BioTek, USA).
Confirmation of antigen specific antibodies by Western blot: The HEL specific antibodies in sera of the immunized animals were determined by Western blot analysis. The HEL was electrophoretically resolved over SDS PAGE (12.5%) and transferred to polyvinyl difluoride (PVDF) membrane. Semi dry blotting steps were followed as described by Towbin et al. (1979). The PVDF membrane was then kept in blocking solution (PBST-5% skim milk powder) for 1 h at room temperature with shaking to block unbound sites. After washing thrice with PBST, the membrane was first incubated with anti-HEL dilution serum (1:400) for 3 h at 37°C and then after proper washing anti-IgG HRPO-conjugate (1:2000) was added for 1 h. The membrane was finally washed thrice and placed in 10 mL of substrate buffer (6 μL of H2O2 and 0.06% Diaminobenzidine tetra-hydrochloride: DAB). The reaction was stopped with distilled water and the blots were dried on a filter paper.
 | |
| Fig. 1: | Characterization of extracted Pasturella multocida LPS in the gel stained by silver staining method. Lane 1: Standard E. coli LPS (Sigma Aldrich, USA) showing ladder like pattern, Lane 2: Pm52 derived LPS showing characteristic band pattern of one behind the other |
RESULTS
Isolation and characterization of Pm52 LPS: During hot/phenol extraction method, Pm52 LPS remained in aqueous phase and on silver staining showed band pattern of one behind the other. The standard Escherichia coli LPS gave the characteristic ladder like pattern (Fig. 1). The obtained amount of Pm52 LPS extracted was about 600 μg g-1 of wet bacterial pellet.
Pravastatin inhibited the in vitro LPS induced NO production from splenocytes: Nitric oxide was determined indirectly by measuring nitrite concentrations in supernatant at 48 and 72 h post LPS stimulation. The Pm52 LPS was found to stimulate NO production in a time and dose dependent manner (p<0.01).
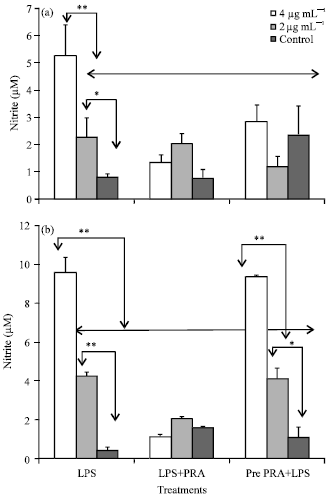 | |
| Fig. 2(a-b): | Effect of Pm52 derived LPS on nitric oxide production from splenocytes. The cells were stimulated either in presence of pravastatin (LPS+PRA) or were pre-incubated with pravastatin for 24 h after which supernatant was discarded and stimulated with LPS (pre PRA+LPS). NO release after (a) 48 h post stimulation and (b) 72 h post stimulation. Values are represented as Mean±SEM of three individual experiments. **p<0.01, *p<0.05 |
Addition of pravastatin (10 μM) to the splenocyte cultures was found to significantly reduce levels of LPS induced NO release both at 48 and 72 h post stimulation (p<0.05) (Fig. 2a-b). Notably, the reduction in nitrite levels due to pravastatin were significantly more in 4 μg mL-1 LPS stimulated cells compared to 2 μg mL-1 LPS stimulated cells at 72 h post stimulation. Pretreatment of cells with Pravastatin was found inferior in reducing the levels of NO in LPS stimulated cultures than their simultaneous use. However, significantly lower levels of NO were produced by splenocytes pre-pretreated with pravastatin at 48 h.
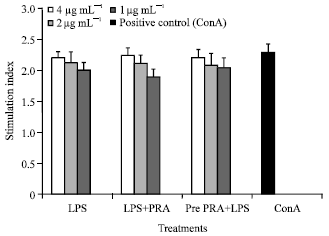 | |
| Fig. 3: | Effect of Pm52 derived LPS on lymphocyte proliferation. Different doses of LPS were used in presence (LPS+PRA) or absence (LPS) of pravastatin to stimulate lymphocytes (2x105) or the lymphocytes were pre-treated with pravastatin for 24 h and then stimulated with LPS (pre PRA+LPS) |
Pravastatin has minimal effect on LPS induced lymphocyte proliferation: Splenocytes were seeded in the 96 well culture plate at concentration of 2x105cells/well and stimulated with graded doses of Pm52 derived LPS (1, 2 and 4 μg mL-1) in presence or absence of pravastatin (10 μM). The LPS was found to stimulate cells almost to the same extent with SI values ranging from 1.92±0.01 (1 μg mL-1) to 2.21±0.02 (4 μg mL-1) compared to standard positive control (Con A, SI value = 2.30±0.01). Cells stimulated in presence of pravastatin or pretreated for 24 h with pravastatin also gave same stimulation indices as those of LPS alone treated cells. Although, a trend of dose dependent LPS stimulation was observed no significant changes were found among different LPS concentrations used (Fig. 3).
Pravastatin mitigated LPS induced excessive serum TNF levels: In order to determine the effects of pravastatin on post LPS induced serum cytokine (TNF-α and IL-10) levels at various time intervals serum samples were collected at 0, 2 and 6 h post immunization from all the groups. For the basal level of TNF-α and IL-10 levels for the present experiment, a total of six mice were randomly selected and average levels were measured. These levels were found to be 324.02±132.28 pg mL-1 for TNF-α and 658.52±109.29 pg mL-1 for IL-10 and were taken as basal references for all the groups. Inoculation of Pm52 derived LPS was found to increase serum TNF-α levels significantly (p<0.001) at 2 h PI and returned almost basal values at 6 h PI. Administration of pravastatin in both the ways as 3 h pretreatment or simultaneous with the LPS+HEL inoculums was found to inhibit LPS induced TNF-α release significantly (Fig. 4).
| Table 2: | Ratio of TNF-α: IL-10 cytokines in the sera of different groups at various post inoculation time intervals |
 | |
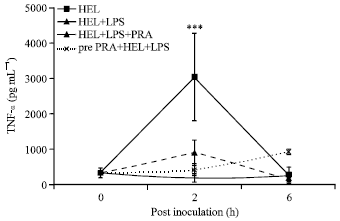 | |
| Fig. 4: | Effect of pravastatin on Pm52 derived LPS on induction of serum TNF-α levels. Lysozyme was inoculated to mice either alone (HEL), with LPS (HEL+LPS) and concurrently treated with pravastatin at antigen inoculation (HEL+LPS+ PRA) or 3 h before the actual immunization as pretreatment (Pre-PRA+HEL+LPS). Values represents Mean±SEM of five mice/group; ***p<0.001 |
Furthermore, pravastatin was found to maintain and balance the serum TNF-α: IL-10 ratio at various post inoculation time intervals (Table 2).
Pravastatin prevented LPS induced elevated serum NOx levels: The immediate effects of Pm52 derived LPS on serum NO levels were determined indirectly by measuring total serum nitrite (NOx). Basal serum NOx level determined for 8 mice was found to be 11.13 ±8.96 μM and was taken as the basal reference value for this study. Inoculation of LPS+HEL was found to significantly increase NOx levels as early as 2 h PI and the values remained high up to 6 h and finally attained basal levels at 24 h PI. Pravastatin was found to inhibit LPS induced NO production significantly (p<0.05). Notably, pretreatment with pravastatin was found to be so effective that no significant changes in serum NO levels were observed in this and control group (Fig. 5).
Pravastatin has minimal effect on LPS associated adjuvant effects: Anti-HEL specific IgG isotype was measured by indirect ELISA. Total IgG response against HEL was found to vary significant within groups (p<0.01). The IgG levels started to increase on 7th day PI in all the groups.
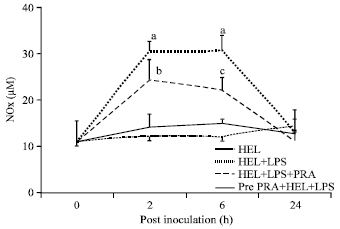 | |
| Fig. 5: | Effect of pravastatin on LPS induced total serum nitric oxide levels (NOx) during early hours after immunization. Mice were inoculated either lysozyme alone (HEL); lysozyme and lipopolysaccharide (HEL+LPS), lysozyme, lipopolysaccharide and pravastatin (HEL+LPS+ PRA) and pretreated with pravastatin (24 h) before actual lysozyme and lipopolysaccharide (pre PRA+lysozyme+lipopolysaccharide). Values represents Mean±SEM of five mice/group, Mean values (a, b, c) differ significantly from other values, p<0.05 |
The total IgG response showed increasing trends till day 14 PI and then onwards showed decreasing trend in all the groups except Group 2 which showed increasing trend up to 21 day PI. The immune response in Group 2 (1.03±0.118) and Group 3 (1.2±0.14) were significantly higher than the other groups on day 14th PI. The highest immune response was recorded on day 14th for Group 3 (1.20±0.14) and on 21st for Group 2 (1.27±0.0.18). Kinetics of antibody response revealed that mice immunized with LPS as adjuvant along with pravastatin showed significantly higher immune response as early as on 7th day (0.74±0.10) than all other group (p<0.05) (Table 3).
Western blotting confirmed the anti-HEL specific antibodies in the serum: The development of anti-HEL IgG specific antibodies were confirmed from the immunized mice at 14th day PI and the relative intensity indicated higher antibody response in LPS adjuvanted animals as compared to HEL alone inoculated mice (Fig. 6).
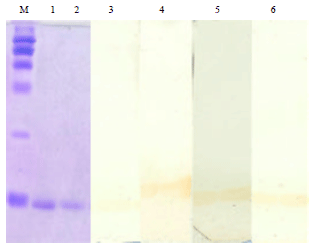 | |
| Fig. 6: | Confirmation of anti-HEL specific antibodies by western blotting in the sera of different groups on 14th day PI. Lane M: Protein weight marker, Lane 1 and 2: HEL in gel stained with commusive brilliant blue, Lane 3: HEL immunized group, Lane 4: HEL+LPS immunized group, Lane 5: HEL+LPS+PRA group and Lane 6: Pravastatin pretreated group immunized with HEL+LPS |
| Table 3: | HEL specific IgG antibody in levels in the sera of mice immunized with Pm52 derived LPS as adjuvant and pravastatin as immunomodulator |
 | |
| Mean values (row wise) bearing different superscript (A, B) vary significantly (p<0.01), Mean values (column wise) bearing different superscript (a, b, c, d ) vary significantly (p<0.01) | |
DISCUSSION
The discovery of PAMPs as powerful activators of the immune system through Toll-Like Receptor (TLR) interaction prompted a variety of TLR agonists to be used as vaccine adjuvants. These agonists activate the innate immune cells, in particular Dendritic Cells (DC), with production of pro-inflammatory cytokines and chemokines resulting in the better induction of antigen specific adaptive immune responses. Among them, bacterial LPS, a TLR4 agonist, is seen as one of the most potential TLR adjuvant for vaccines (Hawkins et al., 2003). However, the induction of systemic toxicity limits its use as vaccine adjuvant. A possible way to overcome this limitation is either to modify the chemical structure of lipid-A part or to counter the systemic toxicity of LPS by immune modifiers/modulators. A detoxified form of lipid A, Monophosphoryl lipid-A (MPLA), has been developed and already successfully used as an adjuvant in vaccine formulations (Petrovsky and Aguilar, 2004; Freytag and Clements, 2005). In this regard, the adjuvant potential of P. multocida derived LPS on a weak immunogen, HEL was explored in the present study. The pravastatin was concurrently administered to determine its short term and long term effects on vaccine induced immune responses.
During extraction, Pm52 LPS remained in an aqueous phase along with nucleic acids due to its amphipathic nature and the finding was in conformity with the Pm70 and PmX73 strains extraction protocols (Michael et al., 2005a, b). The extracted Pm52 LPS showed characteristic band pattern of one behind the other, atypical for P. multocida serotype B strain (Rimler, 1990). In order to verify the biologic activity of extracted Pm52 LPS, the isolated splenocytes were in vitro stimulated with Pm52 LPS for NO production in the present study. It was found that splenocytes stimulated with LPS (2 and 4 μg mL-1) produced NO production in time and concentration dependent manner (p<0.05). Although, higher concentration of Pm52 LPS were required to study its effect on NO production by splenocytes in the present study, the possible explanation for this lower potency may be the aggregative behavior of Pm52 derived LPS through its lipid A domain preventing its accessibility to cells in the culture medium. Low NO production by cells during in vitro trials does not necessarily reflect the true biological activity of the extracted LPS as during in vivo conditions an acute phase protein, LPS binding protein, favour its disintegration whereby it exhibits its biological activity in realistic manner (Hodgson, 2006).
During vaccination or infection immune response is greater against various components of the antigen, out of which several antigen moiety may act either as a synergistic or suppressive to the net outcome. To overcome the possibility of such effects, in the present study purified antigen, HEL was selected to detect the immunomodulatory effect of LPS. A large number of references are available regarding use of lysozyme as an antigen with variable result depending upon the dose used and genetic makeup of the species resulting either high or low response (Furman and Sercarz, 1981; Keller et al., 1989). For practical purpose, vaccine has to be used across species and beyond genetic makeup. In this regard, out bred Swiss Albino mice with heterogeneous genetic makeup were immunized with HEL to evaluate the LPS activity. It was found that inoculation of HEL in conjunction with LPS induced higher immune stimulation and development of higher antibody levels in mice at all days PI. The immune response kinetics showed that IgG response was significantly higher in all HEL+LPS inoculated group as compared to HEL alone administered mice (p<0.05). While group immunized with HEL+LPS showed highest immune response on 21 dpi, significantly higher IgG levels were found in group immunized with HEL+LPS+Pravastatin as early as 7th dpi (p<0.01). The LPS acts primarily by activating signal ‘0’ through TLR and indirectly through signal ‘2’ by activating antigen presenting cells and triggering the secretion of cytokines such as IL-12 in a similar way as by its detoxified product MPLA (Brunner et al., 2010). However, the immediate effects of LPS inoculation also led to induction of septic signs and the inflammatory status.
Administration of LPS induces septic shock by regulating multiple pathways. The TNF-α and NO are the two central molecules released during early stages of LPS induced sepsis. TNF-α is a primary mediator of inflammation and has pleiotrophic effects on target cells (Blackwell and Christman, 1996) while NO induces septic shock by its toxic effects on various cells leading to disseminated intravascular coagulation. The plasma TNF-α and NO levels increase within hours after LPS inoculation and NO levels increases due to increased activity of inducible nitric oxide synthase (iNOS) (Nathan, 1992). However, NO being highly unstable undergoes rapid reduction in serum and changed to nitrite and nitrate, the stable biological products. In the present study, serum samples were collected at 0, 2 and 6 h post immunization. It was observed that TNF-α concentration followed an increasing trend in all the groups which received LPS in the inoculums at 2 h PI. Moreover, highest increase was observed in the group which received HEL+LPS while it was lowest in antigen alone administered group.
Current evidences suggest that statins have pleoptripic effects with both immunomodulatory and suppressive effects (Jamroz-Wisniewska and Bettowski, 2005; Wani et al., 2013). There is also possibility that an indiscriminate use of statins can lead to change in gene expression profile in the human populations. In this regard, the present study demonstrates the effect of pravastatin on LPS induced inflammatory mediators during in vitro and in vivo conditions. It has been hypothesized that inhibition of NO during earlier phases would be novel strategy for septic shock treatment (Iskit and Guc, 2004). It was found that addition of pravastatin decreased the LPS induced NO production significantly from splenocytes to basal levels. Furthermore, pravastatin administration was found to decrease the LPS induced serum nitrite levels both concurrently as well as pre LPS inoculation. This indicates that pravastatin can maintain the basal levels by inhibiting iNOs as verified by other studies (Madonna et al., 2005). Similarly, pravastatin was found to have significantly reduced, LPS induced TNF-α production at 2 h PI in both pretreatment as well as in simultaneous treatment groups. Possibly pretreatment with pravastatin 3 h before may have delayed the cytokine secretion at the beginning and once its bioavailability was reduced the negative feedback back mechanism was disrupted resulting in higher production of cytokine by LPS moieties. The results were in agreement with previous reports using cervastatin to prevent the mortality in LPS induced septic shock and effects on TNF-α and nitrite and nitrate levels (Ando et al., 2000). While the suppression of inflammatory mediator causing septic signs is necessary, for eliciting an effective cell mediated immune responses and immunoglobulin development, proper and balanced proliferation of lymphocytes is essential. It was found that pravastatin has minimal effect on lymphocyte proliferation using LPS stimulated splenocytes cultures. The different concentration of Pm52 LPS stimulated splenocytes to the same extent either alone or in presence of pravastatin.
During normal physiological processes and controlled immunopathological pathways, the immune system follows multiple strategies for immune regulation in particular by IL-10 secretion to counter act hyper immune stimulation (Wani et al., 2013). Scientific literature suggested that IL-10 is a peliotrophic cytokine produced by a variety of activated cell types and affects immune responses ranging from regulatory T-cell function to inhibition of macrophage activation (Siewe et al., 2006). Several findings suggest the elevated IL-10 provides a protective mechanism that prevents TNF-α production (De Waal Malefyt et al., 1991; Gerard et al., 1993; Howard et al., 1993). In the present finding higher IL-10 levels could not be detect in any of the immunized group receiving antigen alone or in combination with LPS. This may be due to dose of LPS received by each mice was quite low and far below the LD50 dose and thus inadequate to elicit IL-10 levels. In comparison most of the septic models induced by lethal doses of LPS, the IL-10 level increased within 1-3 h and subsequently attains a basal level at 12 h post inoculation (Berg et al., 1995; Durez et al., 1999). The IL-10 levels were within the basal levels which may be due to endogenous IL-10 secretion of the host and there the ratio of TNF-α: IL-10 ratios were stabilized by pravastatin during LPS administration.
CONCLUSION
In conclusion, the present study demonstrates that Pm52 LPS stimulates Nitric Oxide (NO) production from splenocytes in time and dose dependent manner while pravastatin (10 μM) inhibited the LPS induced NO production. Moreover, pravastatin (10 μM) does not inhibited the proliferative responses of splenocytes by Pm52 extracted LPS. The co-administration of Pm52 LPS with HEL significantly increased TNF-α and NO levels at 2 and 6 h, respectively. Furthermore, LPS significantly increased the HEL specific IgG development. Administration of pravastatin reduces serum TNF-α and NO levels, the pro-inflammatory mediators responsible for induction septic shock and stabilized TNF-α: IL-10 ratio without affecting HEL specific adaptive immune responses. The use of LPS and pravastatin combination can modulate immune system of the host animal in a beneficial way. However, further studies are required in this direction to determine the long term, immune modulating effects of LPS and pravastatin on the host animals.
REFERENCES
- Akira, S., K. Takeda and T. Kaisho, 2001. Toll-like receptors: Critical proteins linking innate and acquired immunity. Nat. Immunol., 2: 675-680.
CrossRef - Ando, H., T. Takamura, T. Ota, Y. Nagai and K.I. Kobayashi, 2000. Cerivastatin improves survival of mice with lipopolysaccharide-induced sepsis. J. Pharmacol. Exp. Ther., 294: 1043-1046.
Direct Link - Barton, G.M. and R. Medzhitov, 2002. Control of adaptive immune responses by toll-like receptors. Current Opin. Immunol., 14: 380-383.
CrossRefPubMedDirect Link - Berg, D.J., R. Kuhn, K. Rajewsky, W. Muller and S. Menon et al., 1995. Interleukin-10 is a central regulator of the response to LPS in murine models of endotoxic shock and the Shwartzman reaction but not endotoxin tolerance. J. Clin. Invest., 96: 2339-2347.
CrossRefDirect Link - Blackwell, T.S. and J.W. Christman, 1996. Sepsis and cytokines: Current status. Br. J. Anaesthesia, 77: 110-117.
CrossRefPubMedDirect Link - Brown, M.S. and J.L. Goldstein, 1986. A receptor-mediated pathway for cholesterol homeostasis. Science, 232: 34-47.
CrossRefDirect Link - Brunner, R., E. Jensen-Jarolim and I. Pali-Scholl, 2010. The ABC of clinical and experimental adjuvants: A brief overview. Immunol. Lett., 128: 29-35.
CrossRefPubMedDirect Link - De Waal Malefyt, R., J. Abrams, B. Bennett, C.G. Figdor and J.E. de Vries, 1991. Interleukin 10(IL-10) inhibits cytokine synthesis by human monocytes: An autoregulatory role of IL-10 produced by monocytes. J. Exp. Med., 174: 1209-1220.
CrossRefDirect Link - Durez, P., T. Appelboom, C. Pira, P. Stordeur, B. Vray and M. Goldman, 1999. Antiinflammatory properties of mycophenolate mofetil in murine endotoxemia: Inhibition of TNF-α and upregulation of IL-10 release. Int. J. Immunopharmacol., 21: 581-587.
CrossRefDirect Link - Freytag, L.C. and J.D. Clements, 2005. Mucosal adjuvants. Vaccine, 23: 1804-1813.
CrossRefDirect Link - Furman, A. and E.E. Sercarz, 1981. The failure of nonresponder mice to develop IgG memory assessed by in vitro culture with an antigen-LPS conjugate. J. Immunol., 126: 2430-2435.
PubMedDirect Link - Gao, F., L. Linhartova, A.M. Johnston and D.R. Thickett, 2008. Statins and sepsis. Br. J. Anaesthesia, 100: 288-298.
CrossRefDirect Link - Greenwood, J., L. Steinman and S.S. Zamvil, 2006. Statin therapy and autoimmune disease: From protein prenylation to immunomodulation. Nat. Rev. Immunol., 6: 358-370.
CrossRefDirect Link - Hawkins, W.G., J. Trcka, N. Segal, N.E. Blachere and J.S. Gold et al., 2003. The role of lipopolysaccharide in T-cell responses following DNA vaccination. Vaccine, 21: 1548-1553.
CrossRefPubMedDirect Link - Hodgson, J.C., 2006. Endotoxin and mammalian host responses during experimental disease. J. Comp. Pathol., 135: 157-175.
CrossRefDirect Link - Howard, M., T. Muchamuel, S. Andrade and S. Menon, 1993. Interleukin 10 protects mice from lethal endotoxemia. J. Exp. Med., 177: 1205-1208.
CrossRefPubMedDirect Link - Iskit, A.B. and M.O. Guc, 2004. A new therapeutic approach for the treatment of sepsis. Med. Hypotheses, 62: 342-345.
CrossRefPubMedDirect Link - Jamroz-Wisniewska, A. and J. Bettowski, 2005. Adverse effects of statins. Int. J. Pharmacol., 1: 210-225.
CrossRefDirect Link - Kawai, T. and S. Akira, 2011. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity, 34: 637-650.
CrossRefPubMedDirect Link - Keller, M.A., M.A. Kaplan, N.S. Kenyon, A. Miller and E.E. Sercarz, 1989. Primary response to lysozyme (HEL) and HEL-LPS in neonatal A/J mice: Presence of characteristic adult pattern of regulatory idiotype and fine specificity restriction. Cell Immunol., 119: 143-152.
CrossRefDirect Link - Madonna, R., P. di Napoli, M. Massaro, A. Grilli and M. Felaco et al., 2005. Simvastatin attenuates expression of cytokine-inducible nitric-oxide synthase in embryonic cardiac myoblasts. J. Biol. Chem., 280: 13503-13511.
CrossRefPubMedDirect Link - Marshall, J.C., 2005. Lipopolysaccharide: An endotoxin or an exogenous hormone? Clin. Infect. Dis., 41: S470-S480.
CrossRefDirect Link - Masuko, T., A. Minami, N. Iwasaki, T. Majima, S.I. Nishimura and Y.C. Lee, 2005. Carbohydrate analysis by a phenol-sulfuric acid method in microplate format. Anal. Biochem., 339: 69-72.
CrossRefPubMedDirect Link - McGown, C.C., N.J. Brown, P.G. Hellewell, C.S. Reilly and Z.L.S. Brookes, 2010. Beneficial microvascular and anti-inflammatory effects of pravastatin during sepsis involve nitric oxide synthase III. Br. J. Anaesthesia, 104: 183-190.
CrossRefDirect Link - Michael, F.S., E. Vinogradov, J. Li and A.D. Cox, 2005. Structural analysis of the lipopolysaccharide from Pasteurella multocida genome strain Pm70 and identification of the putative lipopolysaccharide glycosyltransferases. Glycobiology, 15: 323-333.
CrossRefPubMedDirect Link - Michael, S.F., J. Li and A.D. Cox, 2005. Structural analysis of the core oligosaccharide from Pasteurella multocida strain X73. Carbohydr. Res., 340: 1253-1257.
CrossRefPubMedDirect Link - Mogensen, T.H., 2009. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev., 22: 240-273.
CrossRefPubMedDirect Link - Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods, 65: 55-63.
CrossRefPubMedDirect Link - Nathan, C., 1992. Nitric oxide as a secretory product of mammalian cells. FASEB J., 6: 3051-3064.
Direct Link - Petrovsky, N. and J.C. Aguilar, 2004. Vaccine adjuvants: Current state and future trends. Immunol. Cell Biol., 82: 488-496.
CrossRefPubMedDirect Link - Rimler, R.B., 1990. Comparisons of Pasteurella multocida lipopolysaccharides by sodium dodecyl sulfate-polyacrylamide gel electrophoresis to determine relationship between group B and E hemorrhagic septicemia strains and serologically related group A strains. J. Clin. Microbiol., 28: 654-659.
Direct Link - Sastry, K.V.H., R.P. Moudgal, J. Mohan, J.S. Tyagi and G.S. Rao, 2002. Spectrophotometric determination of serum nitrite and nitrate by copper-cadmium alloy. Anal. Biochem., 306: 79-82.
CrossRefPubMedDirect Link - Siewe, L., M., Bollati-Fogolin, C. Wickenhauser, T. Krieg, W. Muller and A. Roers, 2006. Interleukin-10 derived from macrophages and/or neutrophils regulates the inflammatory response to LPS but not the response to CpG DNA. Eur. J. Immunol., 36: 3248-3255.
CrossRefPubMedDirect Link - Strunk, T., A. Currie, P. Richmond, K. Simmer and D. Burgner, 2011. Innate immunity in human newborn infants: Prematurity means more than immaturity. J. Maternal-Fetal Neonatal Med., 24: 25-31.
CrossRefPubMedDirect Link - Towbin, H., T. Staehelin and J. Gordon, 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA., 76: 4350-4354.
PubMedDirect Link - Tsai, C.M. and C.E. Frasch, 1982. A sensitive silver stain for detecting lipopolysaccharides in polyacrylamide gels. Ann. Biochem., 119: 115-119.
CrossRefDirect Link - Wani, M.Y., T.K. Goswami, R.A. Mir, D. Chaudhary and G.C. Ram, 2011. Isolation, quantification and characterization of Pasteurella multocida Pm52 derived lipopolysaccharaide and its effect on nitric oxide production in mice. J. Immunol. Immunopathol., 13: 75-82.
Direct Link - Wani, M.Y., T.K. Goswami, R.A. Mir, P. Chaudhuri and K. Dhama, 2013. Moderation of immunopathological parameters by pravastatin in Pasteurella multocida (Pm52) induced septicaemic mice. Int. J. Pharmacol., 9: 513-523.
CrossRefDirect Link - Yeo, C.D., C.K. Rhee, I.K. Kim, H.H. Kang and S.H. Lee et al., 2013. Protective effect of pravastatin on lipopolysaccharide-induced acute lung injury during neutropenia recovery in mice. Exp. Lung Res., 39: 99-106.
CrossRef