
Rosa Martha Perez Gutierrez
Laboratorio de Investigaci�n de Productos Naturales, Escuela Superior de Ingenieria Quimica e Industrias Extractivas IPN. Av. Instituto Politecnico Nacional S/N, Unidad Profesional Adolfo Lopez Mateos CP 07708, Mexico DF
Vicente Aguiniga Juarez
Laboratorio de Investigaci�n de Productos Naturales, Escuela Superior de Ingenieria Quimica e Industrias Extractivas IPN. Av. Instituto Politecnico Nacional S/N, Unidad Profesional Adolfo Lopez Mateos CP 07708, Mexico DF
Jahel Valdes Sauceda
Laboratorio de Investigaci�n de Productos Naturales, Escuela Superior de Ingenieria Quimica e Industrias Extractivas IPN. Av. Instituto Politecnico Nacional S/N, Unidad Profesional Adolfo Lopez Mateos CP 07708, Mexico DF
Irasema Anaya Sosa
Departamento de Graduados de Alimentos, Escuela Nacional de Ciencias Biologicas IPN. Carpio/Plan de Ayala S/N, Santo Tomas Mexico DF
International Journal of Pharmacology
Year: 2014 | Volume: 10 | Issue: 7 | Page No.: 368-379







ABSTRACT
In this study the antidiabetic and antiglycation effects of Apium graveolens was evaluated. Streptozotozin-induced diabetic rats type 1 and 2 were orally treated with chloroform extract (AG-C) for 30 days. The extract (400 mg kg-1) showed good oral glucose tolerance test, effect in both normoglycaemic and hyperglycaemic rats, an antioxidant effect decreasing in the serum level of TBARS and gave its optimum antioxidant enzymes, in liver, kidney and pancreas, decrease levels of glucose and glycosylated hemoglobin, reduced total cholesterol, low-density cholesterol and triglycerides, while increased high-density cholesterol, aspartate, alanine aminotransferase, alkaline phosphatase, total bilirubin, total protein and insulin in serum and pancreas. Also protected RIN-5F cells from AGEs-induced oxidative stress. Gluconeogenic enzymes was significantly increased while hexokinase was decreased in the liver along with glycogen. The AG-C have a significant anti-glycative activity in vitro and it can also effectively protect the BSA during glycation. Their capacity to scavenge methylglyoxal suggested carbonyl scavenging as a major mechanism of antiglycation and showed potent Amadorin activity when compared to aminoguanidine. The AG-C after 30 days of treatment had beneficial effects on renal metabolic abnormalities, including oxidative stress and AGE formation. Results, support that AG-C improves glucose metabolism by reducing insulin resistance and stimulating insulin production by protecting pancreatic β-cells from oxidative stress, inhibits lipid abnormalities and have a hepatoprotective and renalprotective role. The close relationship between lipid peroxidation and nonenzymatic protein glycation (AGEs/ALEs) suggests that the antidiabetic activity of the celery could be due to the synergistic effect of antioxidant activity and antiglycation activity.
PDF Abstract XML References Citation
Received: July 06, 2014;
Accepted: October 20, 2014;
Published: October 31, 2014
How to cite this article
Rosa Martha Perez Gutierrez, Vicente Aguiniga Juarez, Jahel Valdes Sauceda and Irasema Anaya Sosa, 2014. In vitro and in vivo Antidiabetic and Antiglycation Properties of Apium graveolens in Type 1 and 2 Diabetic Rat. International Journal of Pharmacology, 10: 368-379.
DOI: 10.3923/ijp.2014.368.379
URL: https://scialert.net/abstract/?doi=ijp.2014.368.379
DOI: 10.3923/ijp.2014.368.379
URL: https://scialert.net/abstract/?doi=ijp.2014.368.379
INTRODUCTION
Diabetes mellitus, one of the most common endocrine metabolic disorders has caused significant morbidity and mortality due to complications (neuropathy, nephropathy, retinopathy, heart attack, stroke and peripheral vascular disease) (Patel et al., 2011). Reactive Oxygen Species (ROS), generated in cells, are fundamental in modulating various physiological functions and represent an essential part of aerobic life and metabolism (Nikki, 2001). Sometimes, excessive generation of these radicals disrupts the antioxidant defense system of the body which may lead to oxidative stress. This situation brings about a variety of disorders including coronary heart disease, diabetes, arthritis, inflammation and cancer.
Advanced Glycation End-products (AGEs) are the final products of the nonenzymatic reaction between reducing sugars and amino groups in proteins, lipoproteins and nucleic acids. Recently, AGEs accumulation in vivo has been considered to play a major role in the pathogenic process of diabetes and its complications (Brownlee, 1995). Thus, the discovery and investigation of compounds with an AGEs inhibitor activity, would certainly offer a potential therapeutic approach for the prevention of diabetes or other pathogenic complications. Management of diabetes is not only related to the increased release of insulin and its action for glycemic but also important in controlling the glycemic index and postprandial hyperglycemia.
Apium graveolens L. (Celery) is a commercially important spice belonging to the family Umbelliferrae. Celery is used in various forms such as fresh herb, stalk, seeds, oil and oleoresin for flavoring of foods and for medicinal purposes (Sowbhagya, 2014). Celery extracts are reported to possess many nutraceutical properties, viz., antioxidant, vasorelaxant, antihypertensive, hepatoprotective, anti-oxidative stress, larvicidal, hypolipidemic, anti-inflammatory, repellent, hypoglycemic, antiulcerogenic, hyperlipemia, hypertension and memory enhancement and antibacterial (Whitehouse et al., 2001). In other study triterpenoids and flavonoids, were isolated (Zhou et al., 2009).
In previous studies, acute treatment with A. graveolens ethanolic extract caused significantly inhibitory effects on total cholesterol and triglyceride level (Iyer and Patil, 2011). In other study, the activities of antioxidant enzymes and the content of malondialdehyde (MDA) in livers of rats were significant decrease (Iyer and Patil, 2011). Hypoglycemic effect of celery leaf was studied on diabetes rats induced alloxan, have shown potential hypoglycemic effect (Jelodar et al., 2007). In the present study, antidiabetic, antiglycation activities, protective effects of celery on diabetes and its complications on the functions of the liver, kidney and pancreas are discussed.
MATERIALS AND METHODS
Plant material: Fresh celery was collected in Amecameca, Mexico State. Voucher specimen (6543) is kept in the Herbarium of Escuela Nacional de Ciencias Biologicas-IPN. The fresh materials for the celery, (stalk and leaves) were cut into small pieces and dried at 40°C and all of the plant materials were finely powdered and passed through a 20 mesh sieve prior to extraction.
Plant extraction: In a separate conical flask, 2 L of hexane, chloroform and methanol, respectively was added into each 200 g of the sample’s powder. The flasks were then placed in an incubated shaker at 200 rpm for three days at temperature of 35±2°C. The mixture was then filtered using Whatman filter paper into a round bottom flask and concentrated using a rotary evaporator at 40-50°C. The crude extracts were weighed and kept in tubes wrapped with aluminium foil to reduce the risk of oxidation.
Animals: The study was conducted in male Wistar albino rats, weighing about 180±30 g. Before and during the experiment, animals were fed a standard laboratory diet (Mouse Chow 5015, Purina) with free access to water. Mice were procured from the bioterium of ENCB and were controlled environment (temperature 25±2°C). Animals were acclimatized for a period of three days in their new environment before the initiation of experiment. Litter in cages was renewed three times a week to ensure hygiene and maximum comfort for animals.
The experiments reported in this study were carried following the guidelines stated in Principles of Laboratory Animal Care (National Institute of Health publication (NIH) 85-23, revised 1985 and the Mexican Official Normativity (NOM-062-Z00-1999). All experiments and protocols described in present study were approved by the Institutional Animal Ethical Committee of Escuela Nacional de Ciencias Biologicas-IPN (Regd. No.639).
Induction of Severe Diabetes (SD): Severe diabetes was induced in overnight fasted male mice by a single intraperitoneal injection of 60 mg kg-1 of streptozotocin (STZ) in a volume 1 mL kg-1 body weight dissolved in cold citrate buffer (pH 4.5) (Zhou et al., 2012). Hyperglucemia was confirmed by measuring glucose 72 h after the streptozotocin shot and 7 days after injection, confirming a high glucose level. Rat with permanent high fasting blood glucose level >300 mg dL-1 were included for the experiments.
Induction of Mild Diabetes (MD): Mild diabetes was induced in overnight fasted mice by administering a single intraperitoneal injection of 45 mg kg-1 b.wt. The STZ in 0.1 mol L-1 cold citrate buffer (pH 4.5), 15 min after the intraperitoneal administration of 120 mg kg-1 nicotinamide. The STZ treated animals were allowed to drink 5% glucose solution over night to overcome drug induced hypoglycemia. After 10 days of development of diabetes, rats with moderate diabetes having persistent glycosuria and hyperglycaemia (blood glucose >250 mg dL-1) were used for further experimentation (Tahara et al., 2008).
Effect of single oral administration of extracts of A. graveolens in glucose level in severe and mild diabetic rat: After the rats had been denied access to food/water overnight, they were randomly divided into thirty three groups (six mice per group) matched for body weight. Normal rats administered distilled water. The other diabetic groups were orally administered 100, 200 and 400 mg kg-1 body weight b.wt. of extracts of hexane (AG-H), chloroform (AG-C) and methanol (AG-M) suspended in Tween 80, 1% via gavage). Diabetic treated rat received glibenclamide (GB) a dose of 5 mg kg-1 b.wt. as standard drug. Blood samples were collected from the tail vein at 0, 2, 4, 6, 8 and 12 h after the administration. The plasma glucose concentration was determined by an enzymatic colorimetric method using a commercial kit (Sigma Aldrich, USA).
Antidiabetic test in chronic severe and mild streptozotocine-induced diabetic rat: In a parallel study eleven groups (n = 10) of diabetic rats were used to determine the chronic effect of AG-C extract. Each group was submitted to a specific treatment, as follows: Normal control and severe and mild diabetic rats, groups, were fed with normal diet and drinking water ad libitum and were given saline by gastric gavage. Severe and mild diabetic rats that received AG-C extract by gastric gavage (400 mg per kg of body weight) every day were designated as SD+AG-C and MD+AG-C groups. Two groups with severe (SD +GB) and mild diabetes (MD +GB) mice were administered with glibenclamide (GB) 4 mg kg-1 as positive control.
Determination of body weight and food intake: The body weight of each rat was measured once each week and the total amount of food and water consumed was recorded 3 times per week.
Oral glucose tolerance test and insulin tolerance test: Rats of each group were orally administered AG-C at doses of 400 mg kg-1 b.wt. on a daily basis for 30 days. At the end of the experiment, an Oral Glucose Tolerance Test (OGTT) was performed to assess the animals’ sensitivity to a high glucose load. Overnight fasted mice were fed orally 2 g glucose kg-1 b.wt. Blood samples were collected from the caudal vein from a small incision at the end of the tail at 0 min (immediately after glucose load), 30, 60, 90 and 120 min after glucose administration. At day 18, Insulin Tolerance Test (ITT) was performed in overnight fasted rats from all groups. Half an hour after administrating extract the rats were intraperitoneally injected with 0.15 U kg-1 of insulin (Recombinant Human Insulin Injection, Lilly). Blood samples were collected in the same way as indicated in OGTT and blood glucose levels were determined at 0, 30, 60 and 120 min after insulin injection.
Antioxidant parameters levels in serum, liver, pancreas and kidney: Activity of serum superoxide dismutase (SOD) was measured by the xanthine oxidase method using commercial kits with the absorbance measured using spectrophotometer at 550 nm. Serum catalase (CAT) and glutathione peroxidase (GSH-Px) activities were measured by the colorimetric method measuring absorbances at 405 and 412 nm, respectively. Glutathione reductase (GSH) by measuring the rate of NADPH oxidation at 340 nm. All the assay kits were purchased from Cayman Chemical (Michigan, USA) and the procedures were according to the kits instructions. In the pancreas the protein concentration was determined by the Bradford method as described in the Bio-Rad protein assay kit. Lipid peroxidation (LPO), that is, thiobarbituric acid reactive substances (TBARS) was estimated by the method of Fraga et al. (1988) and expressed as μM g-1 of liver and kidney tissue. Serum Glutamate Oxaloacetate Transaminase (SGOT), Glutamate Pyruvate Transaminase (SGPT), Serum Alkaline Phosphatase (SALP), Total Bilirubin (TB) and Total Protein, using a commercial Diagnostic Kit Biocompare, BioVision, Biocompare and Thermo scientific, respectively. Malondialdehyde (MDA) as thiobarbituric acid reactive substances was measured at 532 nm spectrophotometrically (Uchiyama and Mihara, 1978).
Serum lipid profile and glucose levels: At the end of the experiment the effect of each treatment was investigated by serum determination of Total Cholesterol (TC), Triglycerides (TG) and HDL-cholesterol, using a commercial Diagnostic Kit (Genzyme Diagnostics) and LDL-cholesterol was calculated as the remaining difference of total cholesterol and HDL. Blood glucose levels were measured employing the glucose oxidase peroxidase (GOD-POD) method (Park et al., 2007).
Assay of glycogen content in liver and skeletal muscle. G6Pase and GK activity in liver: The glycogen content, GK and G6Pase activity were measured according to Assay Kit hich were purchased from Bio-Rad Laboratories. CA, USA.
Determination of serum insulin level, pancreatic insulin content: Serum insulin and pancreatic insulin content were measured by Enzyme Linked Inmunosorbent Assay (ELISA) using the kit (Boehringer Mannheim Diagnostic, Mannheim, Germany). The level of insulin was expressed in μIU mL-1.
Protective effect in pancreatic β-cells against oxidative stress RIN-5F cells: These cells derived from rat pancreatic β-cells were obtained from American Type Culture (Collection; number: CRL-2058) were maintained in RPMI 1640 medium supplemented with 10% (v/v) FBS, streptomycin (100 μg mL-1) and penicillin G (100 U mL-1) (10% FBS/RPMI 1640) under an atmosphere of 5% CO2, 95% humidified air at 37°C. Cells (5x105 cells/well) were cultured in Nunc 12-place multiwell plates. Thereafter, RIN-5F cells received 1 mL of fresh medium (1% FBS/RPMI 1640) without or with Byrsoninas A and B and AGEs for another 3 h. The effect Byrsoninas A and B and AGEs on oxidative stress was examined using measurement of intracellular ROS levels based on ROS-mediated conversion of non-fluorescent 2’,7’-dichloro-dihydrofluorescein diacetate (DCFH-DA) (Hosseinzadeh et al., 2002). The intensity of fluorescence reflects enhanced oxidative stress, after 3 h of incubation, RIN-5F cells were incubated with DCFH-DA (25 μM) in 1% FBS/RPMI 1640 at 37°C for 20 min, at the end of incubation, DCFH fluorescence of the cells from each well was measured at an emission wavelength of 530 nm and an excitation wavelength of 488 nm using a flow cytometer (Becton Dickinson, San Jose, CA, USA).
Bovine Serum Albumin (BSA) glucose assay: The methodology was based on that of Brownlee (1995) BSA (l0 mg mL-1) was incubated with glucose (500 mM) in Phosphate Buffered-Saline (PBS) (5 mL total volume, pH 7.4) and extract containing 0.02% sodium azide at 37°C with a final concentrations of BSA (2 mg mL-1), glucose (40 mM), sample (0.1-0.5 mg mL-1). All the reagent and samples were sterilized by filtration through 0.2 μm membrane filters. The protein, the sugar and the prospective inhibitor were included in the mixture simultaneously. Aminoguanidine was used as an inhibitor positive control. Reactions without any inhibitor were also set up. Each solution was kept in the dark in a capped tube. After 15 days of incubation, fluorescence intensity (excitation wavelength of 370 nm and emission wavelength of 440 nm) was measured for the test solutions.
BSA-Methylglyoxal assay: This assay was modified based on a published method (Rahbar and Figarola, 2003). The assay evaluates the middle stage of protein glycation. The BSA and methylglyoxal were dissolved in phosphate buffer (100 mM, pH 7.4) to a concentration of 20 mg mL-1 and 60 mM, respectively. Isolated were dissolved in the same phosphate buffer. One milliliter of the BSA solution was mixed with 1 mL of methylglyoxal solution and 1 of mL compounds 1 and 2. The mixture was incubated at 37°C. Sodium azide (0.2 g L-1) was used as an aseptic agent. Phosphate buffer was used as a blank. Aminoguanidine and phloroglucinol were used as positive controls. After seven days of incubation, fluorescence of the samples was measured using an excitation of 340 nm and an emission of 420 nm, respectively.
Amadorin activity: Amadorin activity was determined using a post-Amadori screening assay (Khalifah et al., 1999). Lysozyme (10 mg mL-1) was incubated with 0.5 M ribose in 0.1 M sodium phosphate buffer containing 3 mM sodium azide, pH 7.4 at 37°C for 24 h. Unbound ribose was removed by dialysis against 4 L of 0.1 M sodium phosphate buffer, pH 7.4 at 4°C for 48 h with 5-6 changes. Following dialysis, the protein concentration was determined using the Bio-Rad standard protein assay kit based on the Bradford dye-binding procedure (Bradford, 1976). Dialysed ribated lysozyme (10 mg mL-1) was reincubated with 10 mg mL-1 of 1-8 and aminoguanidine in 0.1 M sodium phosphate buffer containing 3 mM sodium azide, pH 7.4 at 37°C for 15 days.
Glycosylated haemoglobin: HbA1c was estimated using a commercial diagnostic kit from Sigma-Aldrich (Human haemolysate [glycated haemoglobin (HbA1c)] Kit).
Glucose and AGE level in kidney: The renal glucose level was determined by the method of Momose et al. (1963). In brief, frozen kidney tissue was homogenized with ice-cold physiological saline and after being deproteinized, the glucose content was determined using the Wako kit described above. The renal AGE level was determined by the method of Nakayama et al. (1993). In brief, minced kidney tissue was dilapidated with chloroform and methanol (2: 1, v/v) overnight. After washing, the tissue was homogenized in 0.1 N NaOH, followed by centrifugation at 8000xg for 15 min at 4°C. The amounts of AGEs in these alkali-soluble samples were determined by measuring the fluorescence at an emission wavelength of 440 nm and an excitation wave length of 370 nm. A native BSA preparation (1 mg mL-1 of 0.1 N NaOH) was used as a standard and its fluorescence intensity was defined as one unit of fluorescence. The fluorescence values of the samples were measured at a protein concentration of 1 mg mL-1 and expressed in arbitrary units (AU).
Mitochondrial TBA-reactive substance level in kidney: Mitochondria were prepared from kidney homogenate by differential centrifugation (800xg and 12000xg, respectively) at 4°C according to the methods of Jung and Pergande (1985) with minor modifications. Each pellet was resuspended in preparation medium and the concentration of TBA-reactive substances was determined by the method of was estimated by the method of Fraga et al. (1988).
Statistical analysis: Data is expressed as Mean±S.E.M of multiple experiments. Paired Student’s t-tests were used to compare two groups or ANOVA with Tukey for multiple comparisons using PRISM software (Graph Pad, San Diego, eA, USA). Values of p<0.05 were considered statistically significant.
RESULTS
Effects of AG-C on blood glucose level of single administration: In streptozotocin-induced untreated diabetic mice, fasting blood glucose level was significantly higher than that of untreated normal ones both before and after AG-C administration. Table 1 shows blood glucose levels in STZ-induced diabetic rat (SD and MD) and normoglycemic rat at different intervals of time with an administration of AG-C extract at doses of 100, 200 and 400 mg kg-1 to SD and MD.
| Table 1: | Effect of single oral administration of chloroform extract of A. graveolens in glucose level in normal, severe and mild diabetic mice |
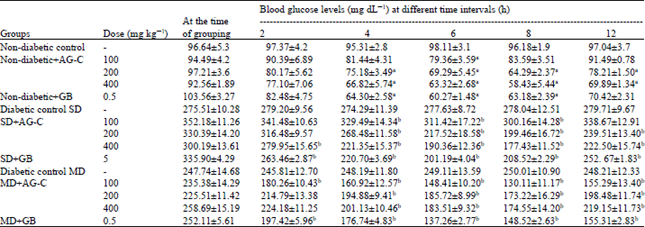 | |
| Each values represent Mean±SD (n = 6). ap<0.05 compared to normal group (ANOVA) followed by Dunnett’s test. bp<0.01 compared to diabetic group (ANOVA) followed by Dunnett’s test. GB: Gibenclamide | |
| Table 2: | Effects of AG-C on blood glucose level of repeated administration in severe and mild diabetic rats. Effect on food intake, body weight and water intake |
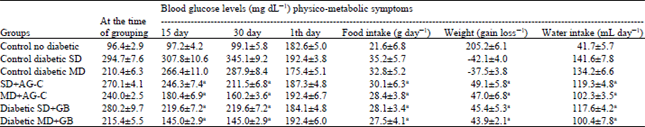 | |
| Values are Mean±SD of 6 rats, values bearing different superscripts in the same column are significantly different (p<0.05) | |
The percentage of glucose reduction with 400 mg kg-1 doses at 8 h was 26 and 32.0%, respectively. Glibenclamide produced a glucose reduction of 38 and 41%, respectively at 8 h. Maximum percentage of blood glucose reduction in normoglycemic mice at 8 h with 400 mg kg-1 doses of AG-C was 37%. However, hexane extract and methanol extract at the same doses produce only a small effect so the results are not presented here. In the study, we found that the chloroform extract of A. graveolens significantly lowered fasting plasma glucose levels in the diabetic mice in a dose-dependent manner.
Effects of AG-C on blood glucose level of repeated administration: Changes of blood glucose level after repeated AG-C and glibenclamide administration in SD and MD diabetic rats were shown in Table 2. After 15 and 30 days of administration of 400 mg kg-1 of AG-C and glibenclamide to diabetic ones, progressive reduction was observed (p<0.01) in blood glucose level of treated diabetic rats during the experimental period.
STZ treatment rapidly produced the characteristic signs of diabetes such as increased intake of both water and food, failure to gain weight (Table 2) and increased blood glucose concentrations. The final body weight showed significant increase from the initial body weight in all the groups except in the diabetic group, in which there was significant decrease in body weight compared to the initial body weight. Although, intake of both water and food were increased compared with the control group. The Table 2 exhibited an improvement of these physico-metabolic symptoms after treatment with the extract when compared to that of the STZ control group. Insulin was not administered. All control of male Wistar rats survived 4 weeks after STZ injection.
Effects of AG-C on glucose loaded STZ diabetic rats and insulin tolerance test: Glucose administration of 2 mg kg-1 orally to STZ diabetic rats produced a significant increase in blood glucose levels (Table 3). AG-C treatment reduced the blood glucose levels significantly. Glucose reduction by AG-C (400 mg kg-1) was less than that of glibenclamide. Diabetic group showed glucose intolerance since blood glucose level increased sharply the intraperitoneal injection of glucose and remained at a very high level after 120 min. In contrast, the rise in glucose level was greatly suppressed in SD+AG-C and MD+AG-C groups, suggesting that this extract prove the impaired glucose tolerance in diabetic rats.
| Table 3: | Effects of AG-C on glucose loaded (OGTT) in SD and MD diabetic rats |
 | |
| Values are expressed as Mean±SD (n = 6), aSignificantly (p<0.05) different from normal rats. bSignificantly (p<0.05) different from diabetic rats. (p<0.05). cSignificantly (p<0.05) different from normal and diabetic rats, where the significance was performed by One way ANOVA followed by post hoc Dunnett’s test | |
| Table 4: | Effects of AG-C on blood glucose levels in insulin tolerance test in SD and MD diabetic rats |
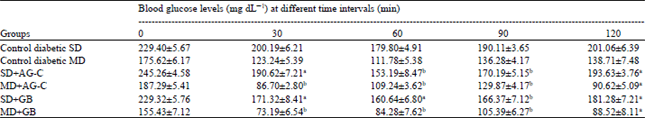 | |
| Blood samples were taken for glucose determinations 0, 30, 60 and 120 min later. Each value represents the Mean±SEM from 6 mice. Significantly different from control, ap<0.05, bp<0.01 (by ANOVA) | |
| Table 5: | Antioxidative status of rats and biochemical parameters at the end of the experimental period |
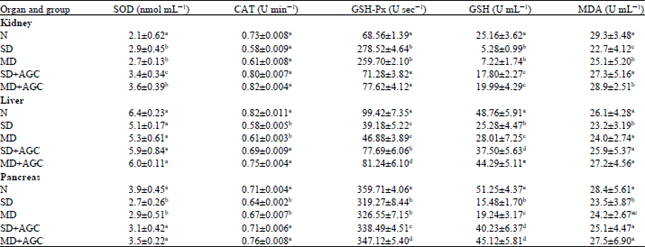 | |
| Each value represents the Mean±SEM from 6 rats. Values within columns bearing the same lower-case letters (a, b, c) do not different at p<0.05 and are not in any particular order. Normal control (N), Diabetic control (SD), Diabetic control (MD) | |
Insulin tolerance test results for AG-C are shown in Table 4. AG-C 10 mg kg-1 treated diabetic rats showed a significant decrease in blood glucose levels 30, 60 and 120 min after insulin administration compared with the controls (30 and 60 min, p<0.01; 120 min, p<0.05). AG-C treated rats also had lower blood glucose levels in the insulin tolerance test and hyperinsulinemia improved. Insulin 0.5 U kg-1 treated rats did not have lower blood glucose levels because of insulin resistance in the peripheral tissues, suggesting that AG-C lessens insulin resistance.
Effects of AG-C on oxidative stress markers and aminotransferases activities: In case of biochemical estimation, improvement of serum enzyme levels were observed in the treated groups with respect to the diabetic control group as shown in Table 5. There was a significant elevation of lipid peroxide, SOD, CAT, GSH-Px and GSH in the pancreas, liver and kidney with diabetes when compared to the normal control group. It was seen that the administration of AG-C helped to decrease these levels which is an indication of the inhibition of oxidative damage of the said tissues.
| Table 6: | Effects of AG-C on serum enzyme levels in normal and hyperglycaemic rats |
 | |
| Each value represents the Mean±SEM (n = 6 rats). ap<0.001 compared with normal control group and bp<0.001 compared with SD and MD control groups. ALP: Alkaline phosphatase, SGOT: Serum glutamate oxaloacetate transaminase, SGPT: Serum glutamate and TB: Total bilirubin | |
| Table 7: | Effect of chloroform extract of L. graveolens on lipid profile, malondialdehyde concentration and on glucose, triglyceride, total cholesterol contents in the kidney of normal and diabetic mice |
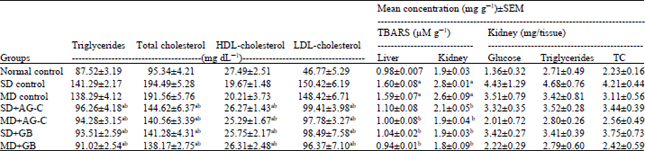 | |
| All values are expressed as Mean±SEM, n = 10. ap<0.05 when compared to normal control group, bp<0.01 when compared to diabetic control group, where the significance was performed by Oneway ANOVA followed by post hoc Dunnett’s test. Plasma insulin values at 0 h before drug administration are significantly different compared to respective days 30 after drug treatment. Significant difference of diabetic control from normal control ap<0.001. Significant difference of treated groups from diabetic control bp<0.01, cp<0.05. dp<0.01 when compared with glibenclamide 5 mg kg-1 treated group | |
The above biochemical parameters were significantly (p<0.01) altered in STZ induced diabetic rats compared to normal control rats. In diabetic rats, administration of AG-C reduced these biochemical parameters (Table 5). Elevation of serum biomarker enzymes such as SGOT, SGPT, TB, ALP and total protein were observed in diabetic rats indicating impaired liver function which is obviously due to hepatocellular necrosis (Table 6). In STZ induced diabetic rats, an elevated level were observed it may be due to STZ mediated liver damages which may cause leakage of above enzymes into the blood (Holst, 2007).
Effects of AG-C on serum lipid profile: The serum lipid profile in control and experimental animals are shown in Table 7. The rise in blood glucose was accompanied with increase in serum total cholesterol, triglyceride, low density lipoprotein, very low density lipoprotein and decreased in high density lipoprotein cholesterol in diabetic rats than those in normal control rats. Treatment with AG-C or glibenclamide to diabetic rats resulted in significant decrease in these parameters compared to those in untreated diabetic rats. However, serum HDL levels were significantly increased in diabetic treated rats. Treatment of AG-C led to significant reductions of renal glucose, triglyceride and total cholesterol contents in diabetic mice (Table 7) which suggested that extract prevented the excessive glucose supply and abnormal lipid accumulation in the kidney. AG-C possessed a definite hypotriglyceridemic property in STZ-induced diabetic rats. This result suggested that AG-C would be helpful in the prevention of diabetic complications through improving dyslipidemia. Supplementation with this extract resulted in a significant (p<0.05) diminution in TBARS levels, compared with controls and levels moved toward values in control group mice (Table 7).
Glucose and AGE level in kidney: Several lines of studies have provided substantial evidence that multiple factors caused by hyperglycemia contribute to the development of diabetic kidney disease. Among them, the impacts of AGEs have been recognized over a wide range, resulting in the express ion and activation of pathogenic mediators implicated in the development of diabetic nephropathy, such as extracellular matrix, oxidative stress, cytokines and growth factors, via., receptor-dependent and/or independent pathways. Therefore, renal AGE accumulation and the mitochondrial lipid peroxidation level were demonstrated first . As a result, diabetic control rats showed increased kidney weight and AGE accumulation significantly, indicating renal hypertrophy and also showed an increased level of TBA-reactive substance. Oral administration of AG-C ameliorated these changes (Table 7). Particularly, AG-C successfully reduced AGE and TBA-reactive substance level at the dose of 400 mg kg-1 suggesting that AG-C suppressed the state of oxidative stress and decreased the levels of serum protein and hemoglobin glycosylated significantly suggesting that it would inhibit oxidative damage and irreversible renal damage caused by the protein glycation reaction under diabetes.
| Table 8: | Effects of AG-C on G6Pase, hexokinase, glycogen content in the liver, plasma, pancreatic insulin, Hb and HbA1c in diabetic rats |
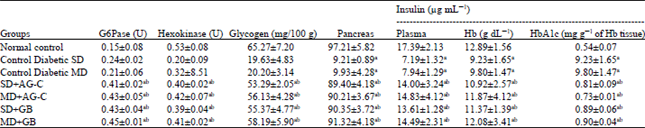 | |
| Each value represents the Mean±S.E.M. (n = 6 rats). ap<0.001 compared with normal control group and bp<0.001, compared with SD and MD control groups | |
Effects of AG-C on liver function, plasma, pancreatic insulin, Hb and and HbA1c endiabetic rats: Table 8 depicts the activities of gluconeagenic enzymes in the liver of the control and experimental rats. The activity of Hepatic Hexokinase (HK) was decreased significantly (p<0.001) and the activities of glucose-6-phosphatase (G-6Pase) was increased significantly (p<0.001) in diabetic rats compared to normal rats. Treatment with AG-C to diabetic rats significantly (p<0.001) increased the activity of hepatic HK and significantly (p<0.001) decreased the activities of G-6Pase compared to diabetic rats which was similar to that of glibenclamide treated rats. Hepatic glycogen content in the liver of diabetic rats were significantly (p<0.001) decreased. Oral administration of AG-C or glibenclamide reversed these glycogen content levels to near normalcy, when compared with diabetic rats.
Furthermore, the attenuating effect of this extract on experimental physiological symptoms of streptozotocin induced diabetes has been confirmed here by the study of glucose-6-phosphatase activity in liver, as well as the quantification of glycogen in liver which are the important indicators of diabetes mellitus.
Table 8 shows the levels of HB, HbAlc of normal and diabetic rats. Diabetic rats showed a significant (p<0.001) elevation in the level of HbA1c and a significant (p<0.001) decrease in the level of total HB compared to normal rats. Treatment with AG-C and glibenclamide to diabetic rats significantly (p<0.001) decreased the level of HbA1c and significantly (p<0.001) increased the level of total HB when compared to diabetic rats. Furthermore, the results also showed that AG-C lowered the elevated serum glucose levels in diabetic rats with significantly increasing serum and pancreatic insulin levels (Table 8), suggesting that AG-C did have any insulin-stimulating effect and the hypoglycemic effect of AG-C observed in this study was associated with a stimulation of insulin secretion. Oral treatment with AG-C significantly reversed the levels of insulin, glucose HB and HbAlc could be due to the insulinotropic effect of AG-C.
| Table 9: | Effect of AG-C on AGEs-induced oxidative stress in RIN-5F |
 | |
| Each value represents the Mean±SEM for 3 assays, *p<0.05. BSA alone (CNT) was used as control. Effects of AGE1 and AGE2 on intracellular ROS levels in RIN-5F cells after 3 h treatment with AGEs | |
Effects of AG-C on AGEs-induced oxidative stress in RIN-5F cells: RIN-5F cells that had been adhered and pre-cultured with medium alone for 72 h were treated with Byrsoninas A and B or medium alone for another 3 h. At the end of the culture, the intracellular peroxide level was measured with DCFH-DA fluorescent probe. The fluorescence intensity for AG-C treated group mice was significantly (p<0.05) lower than the intensity for control (no treatment) group mice. This result suggested that AG-C might reduce oxidative stress in RIN-5F cells. To verify this hypothesis, RIN-5F cells were given oxidative stress by adding AGE1 and AGE2, BSA conjugated with glucose and glyceraldehyde, respectively, to experimental media. After 3 h treatment with AGEs, fluorescence intensity of AGE1 and AGE2 treated cells was significantly higher than that of the CNT-treated cells (p<0.05). In the same experiments with AGEs, treatment of RIN-5F cells with AG-C for 3 h resulted in dramatic reductions in oxidative stress with significant differences (p<0.05). These results (Table 9) clearly indicated that AG-C protected pancreatic β-cells from AGEs-induced oxidative stress.
Bovine Serum Albumin (BSA) glucose, BSA-methylglyoxal and amadorin activity: In order to determine the inhibitory effect of AG-C on the formation of AGEs, several assay methods have been proposed, Including assays based on the inhibition of specific fluorescence generated during the course of glycation and AGE formation and assays based on the inhibition of AGE-protein cross-linking. Table 10, displays the inhibitory effects of AG-C on AGE formation in BSA-glucose, Amadori products and BSA-methylglyoxal models.
| Table 10: | Inhibitory effects of chloroform extract from (AG-C) and aminoguanidine on the formation of advanced glycation end products (AGEs), in vitro induced by glucose, methylglioxal and ribose |
 | |
| Data are Mean±standard deviation of triplicate tests | |
| Table 11: | Effect of AG-C on renal mitochondrial TBA-reactive substance, renal weight and AGE levels |
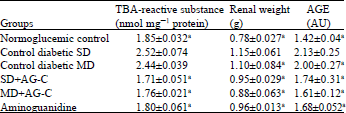 | |
| Data as expressed as±SD; ap<0.05 vs. diabetic control values | |
Glycation is accompanied by metal-catalysed oxidation of glucose and Amadori products to form free radicals capable of protein fragmentation. Lysozyme was glycated by glucose or methylglyoxal in the presence of AG-C. This ribated lysozyme was reincubated and the effects of AG-C and aminoguasnidine on glycation induced crosslinking was monitored. AG-C inhibited formation of glucose and methylglyoxal derived advanced glycation endproducts and showed potent Amadorin activity when compared to aminoguanidine.
Effect of AG-C on renal mitochondrial TBA-reactive substance, renal weight and AGE levels: Prolonged exposure to hyperglycemia in diabetes participates in the formation and accumulation of AGEs which correlates with the severity of renal complications in diabetes. Table 11 shows the results of biochemical analysis. The kidney weight of diabetic control rats was heavier than that of normal rats but it was reduced by administrations of AG-C (p<0.01).
TBA-reactive substance and AGE levels significantly reduce with the administration of AG-C. Since the administration of AG-C had a marked effect on glucose and glucose-associated oxidative stress rather than on lipids, as shown in the serum and renal constituents, we subsequently focused on intracellular glycation and AGE formation in kidney tissue. The AGE formation, one of the metabolic disorders caused by hyperglycemia, has been focused on as a marker of long-term glycemic control in body tissues. These products have also been strongly implicated in the pathogenesis of diabetic micro- and macrovascular diseases, because AGE-modified proteins can stimulate a variety of cellular responses.
DISCUSSION
In this study, induction by STZ of SD produced insulin deficient rats, high doses of STZ has been selectively destroys insulin-produced β-cells of the pancreas (Aybar et al., 2001). In contrast, STZ-induced diabetic rats pre-treated with nicotinamide (MD) exhibited moderate hyperglycemia associated with loss of postprandial early-phase insulin secretion (Sharma et al., 2011). Because of this we decided to study the celery in both models (SD and MD) partial destruction of β-cells of the pancreas. The experimental diabetic model used low dose of STZ (45 mg kg-1 b.wt.) destroyed partial some population of pancreatic β-cells. There were residual β-cells which secreted insufficient insulin causing type 2 diabetic models which lead to impaired glucose uptake by peripheral cells and impaired insulin secretion and action and contributes a number of features similar with human diabetes. The STZ-diabetic mice showed significantly decreased plasma glucose level on treatment with AG-C which might bring about glucose lowering action by stimulating the surviving β-cells of islets of Langerhans to release more insulin.
The reduction in body weight may be attributed to insulin depletion provoking a loss of adipose tissues or might also be the result of degradation of structural proteins due to unavailability of carbohydrate as energy source or due to the antihyperglycemic effect of AG-C thereby enhancing glucose metabolism (Pepato et al., 1996).
Oxidative stress in diabetes mellitus has been shown to co-exist with a reduction in the endogenous antioxidant status. Oxidative stress is caused by a relative overproduction of Reactive Oxygen Species (ROS). The ROS results in lipid peroxidation and subsequently increased in TBARS levels leading to degradation of cellular macromolecules.
A marked increase in the concentration of TBARS in STZ-induced diabetic rats indicates enhanced lipid peroxidation leading to tissue injury and failure of the antioxidant defense mechanisms to prevent overproduction of ROS. Treatment with AG-C inhibited hepatic, kidney and pancreas lipid peroxidation in diabetic rats as revealed by reduction TBARS levels toward normal levels. This indicated the inhibition in free radicals (ROS) generation in STZ-induced diabetic rats (Matkovics et al., 1998).
Glutathione plays an important role in the endogenous nonenzymatic antioxidant system. Primarily, it acts as reducing agent and detoxifies hydrogen peroxide in presence of the enzyme, glutathione peroxidase. The depleted GSH may be due to reduction in GSH synthesis or degradation of GSH by oxidative stress in STZ-induced hyperglycemic animals (Mandal et al., 2011). AG-C treatment significantly elevated the reduced hepatic, kidney and pancreas glutathione levels toward normal in diabetic rats. The results showed that the antihyperglycemic activity of AG-C was accompanied with the enhancement in nonenzymatic antioxidant protection. These findings suggest that the AG-C may exert its antihyperglycemic effect through the enhancement of cellular antioxidant system.
CAT is a heme-containing enzyme catalyzing the detoxification of H2O2 to water and oxygen The inhibition of CAT activity as a result of STZ-induced hyperglycemia were observed in our present study. AG-C treatment significantly recovered the CAT activity toward normal in liver, kidney and pancreas. SOD, protects oxygen metabolizing cells against harmful effects of superoxide free-radicals. AG-C treatment resulted in significant modulation of lipid peroxidation, endogenous nonenzymatic (GSH) and enzymatic (SOD, CAT) antioxidant and detoxification systems (Matsuura et al., 2002).
TBARS levels are an index of endogenous lipid peroxidation and oxidative stress with intensified free radical production. Therefore, measurement of TBA-reactive substance levels is frequently used to determine the level of oxidative stress in diabetic patients. In addition, increased lipid peroxidation in the pancreas implies a level of susceptibility to diabetic oxidative stress. TBARS levels in both the liver and kidneys of diabetic control group mice were high and were clearly reduced (p<0.001) upon administration of AG-C. These elevated TBARS levels in diabetic mice might be due to stimulation of hepatic triglyceride synthesis as a result of a free fatty acid influx (Pociot et al., 1993). Repeated separate administration of AG-C have a beneficial effect for reducing the degree of hyperlipidemia that is associated with hyperglycemia.
In the diabetic state, the glycation reaction is observed in different tissues and organs and for different kinds of glycated proteins (Latha and Daisy, 2011), therefore, the results of this study demonstrated that AG-C protected RIN-5F cells from AGEs-induced oxidative stress followed by decreasing insulin gene expression and secretion. The serum insulin levels of mice in the AG-C groups tended to increase, compared with the level in diabetic control mice (p<0.001), suggesting that AG-C partially rescued exhausted pancreatic β-cells from further AGEs-induced oxidative stress. AGEs might cause deterioration in the function of pancreatic β-cells in patients with long-term hyperglycemia. Therefore, AG-C can act as suppressors of pancreas cell damage and can inhibit progression of the pancreatic dysfunction induced by chronic hyperglycemia.
As discussed earlier in the introduction section, stalks and stem celery containing many important phytoconstituents like essential oils, phenolic, compounds, triterpenes and f1avonoids kaempferol, quercetin, apigenin and luteolin which are proven to be potent hypoglycaemic agents and their hypoglycaemic activity and could prevent oxidative stress. Kaempferol, may be a naturally occurring anti-diabetic compound by protecting pancreatic β-cell survival and function in a hostile environment that would otherwise lead to type 2 diabetes (Kasetti et al., 2010). Quercetin and naringenin might possibly be able to protect β-cells from cytokines toxicity by enhancing cell survival through PI3-kinase pathway, independent of p-p38 MAPK or iNOS (Pari and Satheesh, 2004). Apigenin and luteolin are potential SGLT2 inhibitor for diabetic neuropathy and luteolin ameliorated inflammation related endothelial insulin resistance in an IKKβ/TRS-1/Akt/eNOS-dependent pathway (Rahbar and Figarola, 2003). Treatment with apigenin, significantly lowered the blood glucose levels of diabetic animals and protected the liver and kidneys against STZ-induced damage in rats (Voziyan et al., 2002). Therefore, it can be postulated that the presence of flavonoids in the extract could be involved in the antihyperglycemic action shown by AG-C.
CONCLUSION
The present results indicate that chloroform extract of Apium graveolens has an ability to reduce hyperglycemia, hiperlipidemic, oxidative stress, insulin resistance, formation of AGE. These activities were comparable with conventionally used glibenclamide. The antiglycation activities of A. graveolens were attributed in part to their antioxidant activity and its abilities to scavenge reactive carbonyls. Furthermore, AG-C alleviated oxidative stress under diabetic conditions through the inhibition of lipid peroxidation, prevent and/or delay the onset renal, pancreatic and hepatic damage. In addition improved glucose tolerance suggesting a decrease in insulin resistance and helping to maintain blood glucose levels steady which may indicate certain induction of peripheral utilization of glucose.
REFERENCES
- Aybar, M.J., A.N. Sanchez Riera, A. Grau and S.S. Sanchez, 2001. Hypoglycemic effect of the water extract of Smallantus sonchifolius (yacon) leaves in normal and diabetic rats. J. Ethnopharmacol., 74: 125-132.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Brownlee, M., 1995. The pathological implications of protein glycation. Clin. Invest. Med., 18: 275-281.
PubMedDirect Link - Fraga, C.G., B.E. Leibovitz and A.L. Tappel, 1988. Lipid peroxidation measured as thiobarbituric acid-reactive substances in tissue slices: Characterization and comparison with homogenates and microsomes. Free Radical Biol. Med., 4: 155-161.
CrossRefDirect Link - Holst, J.J., 2007. The physiology of glucagon-like peptide 1. Phys. Rev., 87: 1409-1439.
CrossRefPubMedDirect Link - Hosseinzadeh, H., M. Ramezani and A.R. Danaei, 2002. Antihyperglycaemic effect and acute toxicity of Securigera Securidaca L. seed extracts in mice. Phytoter. Res., 16: 745-747.
CrossRefDirect Link - Iyer, D. and U.K. Patil, 2011. Effect of chloroform and aqueous basic fraction of ethanolic extract from Apium graveolens L. in experimentally-induced hyperlipidemia in rats. J. Complementary Integr. Med., Vol. 8.
CrossRefDirect Link - Jung, K. and M. Pergande, 1985. Influence of cyclosporin A on the respiration of isolated rat kidney mitochondria. FEBS Lett., 183: 167-169.
PubMedDirect Link - Jelodar, G., M. Maleki and S. Sirus, 2007. Effect of fumitory, celery and lemon on blood glucose and histopathology of pancreas of alloxan diabetic rats. J. Applied Anim. Res., 31: 101-104.
CrossRef - Khalifah, R.G., J.W. Baynes and B.G. Hudson, 1999. Amadorins: Novel post-Amadori inhibitors of advanced glycation reactions. Biochem. Biophys. Res. Commun., 257: 251-258.
CrossRefPubMedDirect Link - Latha, R.C.R. and P. Daisy, 2011. Insulin-secretagogue, antihyperlipidemic and other protective effects of gallic acid isolated from Terminalia bellerica Roxb. in streptozotocin-induced diabetic rats. Chem. Biol. Interact., 189: 112-118.
CrossRefPubMedDirect Link - Mandal, S., B. Hazra, R. Sarkar, S. Biswas and N. Mandal, 2011. Assessment of the antioxidant and reactive oxygen species scavenging activity of methanolic extract of Caesalpinia crista leaf. Evidence-Based Complementary Alter. Med.
CrossRefDirect Link - Matkovics, B., M. Kotorman, I.S. Varga, D.Q. Hai and C. Varga, 1998. Oxidative stress in experimental diabetes induced by streptozotocin. Acta Physiologica Hungarica, 85: 29-38.
PubMedDirect Link - Matsuura, N., T. Aradate, E. Sasaki, H. Kojima and M. Ohara, J. Hasegawa and M. Ubukata, 2002. Screening system for the Maillard reaction inhibitor from natural product extracts. J. Health Sci., 48: 520-526.
CrossRef - Nakayama, H., T. Mitsuhashi, S. Kuwajima, S. Aoki, Y. Kuroda, T. Itoh and S. Nakagawa, 1993. Immunochemical detection of advanced glycation end products in lens crystallins from streptozocin-induced diabetic rat. Diabetes, 42: 345-350.
PubMed - Pari, L. and M.A. Satheesh, 2004. Antidiabetic activity of Boerhaavia diffusa L.: Effect on hepatic key enzymes in experimental diabetes. J. Ethnopharmacol., 91: 109-113.
CrossRefDirect Link - Patel, D.K., R. Kumar, S.K. Prasad, K. Sairam and S. Hemalatha, 2011. Antidiabetic and in vitro antioxidant potential of Hybanthus enneaspermus (Linn) F. Muell in streptozotocin-induced diabetic rats. Asian Pacific J. Trop. Biomed., 1: 316-322.
CrossRefDirect Link - Pepato, M.T., R.H. Migliorini, A.L. Goldberg and I.C. Kettelhut, 1996. Role of different proteolytic pathways in degradation of muscle protein from streptozotocin-diabetic rats. Am. J. Physiol., 271: E340-E347.
PubMedDirect Link - Pociot, F., J.I. Reimers and H.U. Andersen, 1993. Nicotinamide-biological actions and therapeutic potential in diabetes prevention. Diabetologia, 36: 574-576.
CrossRefPubMedDirect Link - Rahbar, S. and J.L. Figarola, 2003. Novel inhibitors of advanced glycation endproducts. Arch. Biochem. Biophys., 419: 63-79.
CrossRefPubMedDirect Link - Kasetti, R.B., M.D. Rajasekhar, V.K. Kondeti, S.S. Fatima and E.G.T. Kumar et al., 2010. Antihyperglycemic and antihyperlipidemic activities of methanol: Water (4:1) fraction isolated from aqueous extract of Syzygium alternifolium seeds in streptozotocin induced diabetic rats. Food Chem. Toxicol., 48: 1078-1084.
CrossRefPubMedDirect Link - Park, S.E., M.H. Cho, J.K. Lim, J.S. Kim, J.H. Kim, D.Y. Kwon and C.S. Park, 2007. A new colorimetric method for determining the isomerization activity of sucrose isomerase. Biosci. Biotechnol. Biochem., 71: 583-586.
CrossRefPubMedDirect Link - Sharma, M., M.W. Siddique, A.M. Shamim, S. Gyanesh and K.K. Pillai, 2011. Evaluation of antidiabetic and antioxidant effects of seabuckthorn (Hippophaerhamnoides L.) in streptozotocin-nicotinamide induced diabetic rats. Open Conf. Proceed. J., 2: 53-58.
Direct Link - Sowbhagya, H.B., 2014. Chemistry, technology and nutraceutical functions of celery (Apium graveolens L.): An overview. Crit. Rev. Food Sci. Nutr., 54: 389-398.
CrossRefDirect Link - Uchiyama, M. and M. Mihara, 1978. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem., 86: 271-278.
CrossRefDirect Link - Voziyan, P.A., T.O. Metz, J.W. Baynes and B.G. Hudson, 2002. A post-Amadori inhibitor pyridoxamine also inhibits chemical modification of proteins by scavenging carbonyl intermediates of carbohydrate and lipid degradation. J. Biol. Chem., 277: 3397-3403.
CrossRefDirect Link - Whitehouse, M.W., D.E. Butters, M.L. Clark and K.D. Ranisford, 2001. NSAID gastropathy: Prevention by celery seed extracts in disease-stressed rats. Inflammopharamacology, 9: 201-209.
CrossRefDirect Link - Zhou, K., F. Zhao, Z. Liu, Y. Zhuang, L. Chen and F. Qiu, 2009. Triterpenoids and flavonoids from celery (Apium graveolens). J. Nat. Prod., 72: 1563-1567.
CrossRefPubMedDirect Link - Zhou, J.Y., S.W. Zhou, S.Y. Zeng, J.Y. Zhou, M.J. Jiang and Y. He, 2012. Hypoglycemic and hypolipidemic effects of ethanolic extract of Mirabilis jalapa L. root on normal and diabetic mice. Evid.-Based Complementary Altern. Med., Vol. 2012.
CrossRefDirect Link