
Abdullah Al- Taher
Department of Pharmacology, Faculty of Veterinary Medicine and Animal Resources, King Faisal University, Al-Ahsa, Saudi Arabia
Mahmoud Kandeel
Department of Pharmacology, Faculty of Veterinary Medicine and Animal Resources, King Faisal University, Al-Ahsa, Saudi Arabia
Hye-Sook Kim
Faculty of Pharmacutical Sciences, Okayama University, 1-1-1 Tsushim-naka, Kita-ku, Okayama, 700-8530, Japan
Yukio Kitade
United Graduate School of Drug Discovery and Medical Information Sciences, 1-1 Yanagido, Gifu 501-1193, Japan
International Journal of Pharmacology
Year: 2014 | Volume: 10 | Issue: 6 | Page No.: 334-339







ABSTRACT
The identification of extra-functional activities of enzymes is an attractive strategy for development of new antiparasitic drugs. In this study, the nuclease properties of Plasmodium Nucleoside Diphosphate Kinase (PfNDK) were demonstrated. The classical effect of PfNDK is the production of nucleotides for DNA/RNA synthesis. Such new nuclease activity of PfNDK implies the incrimination of the enzyme in functions other than its classical kinase activity. Such functions include apoptosis, regulation of cell cycle, cell division and repair of DNA damage. We have investigated the nuclease activity of Plasmodium falciparum Nucleoside Diphosphate Kinase (PfNDK). The nature of the nucleotide sequence and the length of the DNA substrate were critical factors in PfNDK’s nuclease activity. The PfNDK was unable to bind to oligonucleotides, although it formed aggregates with oligonucleotides that contained repeated pyrimidine nucleotides in their sequence. The enzyme was unable to cleave a supercoiled plasmid, whereas DNA substrates of various lengths were cleaved in a time- and concentration-dependent manner. Furthermore, ATP at a concentration of 1 mM was able to inhibit the nuclease activity. These data is valuable in highlighting the possible role of PfNDK in cellular functions other than its classical nucleotide kinase activity. Specific PfNDK inhibition is expected to modulate parasites cell cycle and induction of apoptosis.
PDF Abstract XML References Citation
Received: April 11, 2014;
Accepted: August 12, 2014;
Published: October 01, 2014
How to cite this article
Abdullah Al- Taher, Mahmoud Kandeel, Hye-Sook Kim and Yukio Kitade, 2014. Cleavage of DNA and Nuclease Properties of Plasmodium Nucleoside Diphosphate Kinase. International Journal of Pharmacology, 10: 334-339.
DOI: 10.3923/ijp.2014.334.339
URL: https://scialert.net/abstract/?doi=ijp.2014.334.339
DOI: 10.3923/ijp.2014.334.339
URL: https://scialert.net/abstract/?doi=ijp.2014.334.339
INTRODUCTION
Nucleoside Diphosphate Kinase B (NDK, nucleoside diphosphate phosphotransferase, PF3D7_1366500, EC 2.7.4.6) helps to maintain the homeostasis of the intracellular nucleotide pool by catalyzing the transfer of high-energy γ-phosphate between nucleotide di-and tri-phosphates (Miranda et al., 2008; Sikarwar et al., 2013).
Plasmodium falciparum Nucleoside Diphosphate Kinase (PfNDK) binds a broad range of ribo-and deoxyribo-nucleotides, thus it can produce all the precursors for DNA and RNA synthesis (Kandeel et al., 2009a; Kandeel and Kitade, 2010). In addition, NDP kinases are thought to have a central role in signal transduction in bacteria, fungi, plants, invertebrates and vertebrates (Hasunuma et al., 2003), DNA damage repair (Jarrett et al., 2012), apoptosis and tumor suppression (Choudhuri et al., 2010; Fancsalszky et al., 2014) and microtubule polymerization (Jeudy et al., 2009).
The discovered functions of NDKs are extended to include important aspects of host-parasite interactions and microbial pathogenicity. The NDK is proved to have a role in persistence of Mycobacterium tuberculosis inside host macrophages by modulation of innate immunity (Sun et al., 2013). Interestingly, NDK is associated with the pathogenicity of some micro-organisms. For instance, NDK is injected from Pseudomonas aeruginosa into the host cells causing cellular toxicity (Neeld et al., 2014).
The NDKs are multifunctional enzymes, although their functions differ between species and need to be experimentally assigned. We report evidence of DNA binding and the nuclease activities of PfNDK. This will be the first report to characterize such unusual activities of a metabolic enzyme (NDK) in P. falciparum. This could be important is discovering potential new therapeutic approaches.
MATERIALS AND METHODS
Expression of recombinant PfNDK: The PfNDK was prepared as previously described (Kandeel et al., 2009a). The recombinant E. coli cells were grown overnight in an LB medium containing ampicillin (50 μg mL-1). The culture was diluted to 1:100 with the same fresh medium and cells were grown at 37°C to mid-log phase (D600 = 0.6). Expression was induced by the addition of IPTG to a final concentration of 1 mM and cell growth was continued at 37°C for 4 h. Cells from 2 L of culture were harvested by centrifugation at 5000xg for 15 min and then stored at -20°C until use. The cells were lysed in an extraction buffer (25 mM Tris-HCl buffer pH 7.2 containing 150 mM NaCl) and then disintegrated by sonication for 40 sec (3 cycles at 3 min intervals) to obtain the cell extract.
Purification of recombinant PfNDK: The lysate was centrifuged at 16,000xg for 15 min at 4°C and the precipitate was discarded. The hexahistidine-tagged PfNDK was purified from the soluble cell extract by using talon metal affinity resin, with a flow rate of 30 mL min-1. After binding and washing with the extraction buffer, the protein was eluted using a linear imidazole gradient (0-150 mM).
Production of nucleic acids substrates: To check the nuclease activity of PfNDK, various substrates were prepared, including supercoiled and linearized plasmids, oligonucleotides and various lengths of DNA substrates obtained by PCR.
Plasmids: Supercoiled pQE30 plasmid (3461 bp) was extracted from E. coli JM109 by using the PureYield Plasmid Miniprep System (Promega, USA). The linearized plasmid was prepared by double digestion of the supercoiled plasmid in a buffer containing the enzymes BamHI and HindIII. The linearized plasmid was gel purified by using a QIAquick gel extraction kit (Qiagen, USA).
Oligonucleotides: The PfNDK-oligonucleotide binding assay was carried out using single-or double-stranded oligonucleotides. The oligonucleotides were fluorescently labeled at their 5’ terminal and their sequences are given in Table 1. To produce the double-stranded substrate, the oligonucleotide and its complementary sequence were annealed by mixing equimolar amounts of each oligonucleotide.
| Table 1: | Sequence of oligonucleotides used in the PfNDK DNA cleavage assay |
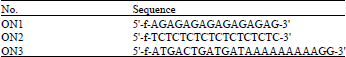 | |
The mixture was heated in boiling water for 10 min and then placed on ice. Three different types of oligonucleotides were used in this assay, ON1, ON2 and ON3. The sequences oligos were designed to check the nucleotide binding preferences of PfNDK. The ON1 is composed of repeats of purine nucleotides (A/G) and used to check the binding PfNDK with purine nucleotides. The ON2 is composed of repeats of pyrimidine nucleotides (T/C) and used to check the binding PfNDK with purine nucleotides. The ON3 is containing internal repeats of adenosine nucleotide (A) and used to check the preferential binding of PfNDK with A/T repeats of nucleotides.
Variable-length DNA substrates: To examine the effect of the length of DNA substrates on PfNDK’s nuclease properties, PCR was used to amplify DNA substrates of variable length. The gene encoding P. falciparum thymidylate kinase was used as a template to obtain the required DNA substrates. Fluorescently labeled forward and reverse primers were used to obtain the full-length gene and then several truncated forms were amplified by PCR using variable internal primers.
Nuclease activity: Variable concentrations of plasmidic linear, supercoiled, or genomic DNA substrates were incubated at 37°C in Tris (pH 7.2, 50 mM), MgCl2 (10 mM) with variable concentrations of PfNDK. The assays were done in the presence or absence of ATP (1 mM) and the incubation time was varied from several minutes to 4 h. The reaction products were resolved in either agarose or polyacrylamide gels.
RESULTS AND DISCUSSION
PfNDK showed different interaction profiles with different oligonucleotides (Fig. 1a). No interaction was observed with oligonucleotides ON1 and ON2, whereas an aggregate was formed with ON3 (Fig. 1b). This interaction was observed with both single-and double-stranded oligonucleotides and the degree of aggregation was related to the enzyme concentration. The bands were checked with non-denaturing polyacrylamide gel electrophoresis which showed no shift in oligonucleotide migration, although a continuous smear and aggregates in the loading wells indicated the presence of enzyme-oligonucleotide aggregates.
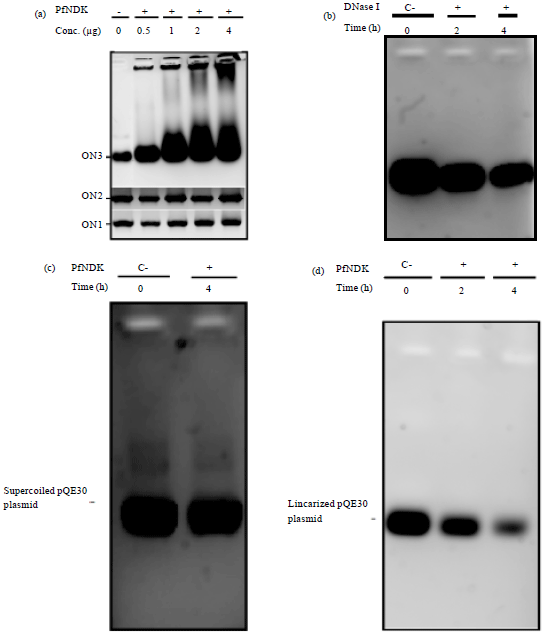 | |
| Fig. 1(a-d): | (a) Agarose gel showing the interaction of different oligonucleotides with several concentrations of PfNDK. The 500 ng of each oligonucleotide was incubated with 0, 0.5, 1, 2 or 4 μg of PfNDK for 2 h. The ON1, ON2 and ON3 refer to the used oligonucleotide. The sequences of ON1, ON2 and ON3 are shown in Table 1. -: No addition of PfNDK, +: Adding PfNDK at the indicated amounts, (b) Control experiment was made to illustrate the control positive nuclease activity. The 1 μg of ON3 was incubated with 0.05 U of DNase I for 0, 2 or 4 h. C: No addition of DNase I, +: Addition of 0.05 U of DNase I, (c) Agarose gel showing the interaction of PfNDK with supercoiled pQE30 plasmid. The 1 μg of pQE30 plasmid was incubated with 4 μg of PfNDK for 0 or 4 h. C: No addition of PfNDK, +: Adding 4 μg of PfNDK and (d) Agarose gel showing the interaction of PfNDK with linearized pQE30 plasmid. The 1 μg of linearized pQE30 plasmid was incubated with 4 μg of PfNDK for 0, 2 or 4 h. C: No addition of PfNDK, +: Adding 4 μg of PfNDK. Prior to assay, supercoiled pQE30 plasmid was linearized by incubation with BamHI and HindIII enzymes at 37°C for 1 h followed by purification from agarose gel electrophoresis bands |
This suggests the preferential binding of PfNDK to repeated sequences of A or T nucleotides. Based on this finding, the cleavage assay was conducted in the presence of variable lengths of a DNA fragment of the Plasmodium gene encoding for thymidylate kinase. Genes rich in A/T nucleotides from P. falciparum can be used to detect the binding of PfNDK to larger substrates.
After a 4 h incubation, no change was observed for the supercoiled plasmid (Fig. 1c), whereas the amount of linearized plasmid is gradually lowered (Fig. 1d). This indicates that PfNDK was unable to nick the supercoiled plasmid, although it was able to bind to linearized forms of DNA.
PfNDK was able to cleave either single-or double-stranded DNA (Fig. 2a) in a time-dependent manner (Fig. 2b and c); the DNA cleavage increased linearly with incubation time. The substrates were produced by PCR under previously described conditions (Kandeel and Kitade, 2008, 2011; Kandeel et al., 2009b). The full length PCR fragment was ~633 bp and this was used as a template to obtain shorter substrates, with previously reported primers (Kandeel et al., 2009b). Interestingly, PfNDK showed nuclease activity for different substrate lengths (Fig. 2d).
ATP is a potent inhibitor of NDK’s nuclease activity (Hammargren et al., 2007). To examine the inhibitory effect of ATP on PfNDK’s nuclease activity, the cleavage assay Was performed in the presence of 1 mM ATP and ATP was found to inhibit the cleavage of ssDNA and dsDNA (Fig. 2e). After evaluation of bands density using ImageQuant 5.2 software, there was about 2-3 folds decrease in nuclease activity of PfNDK in the presence of 1 mM ATP.
This study reports a function of PfNDK which is different from its classical kinase activity. The DNA cleavage activity of PfNDK indicates that the enzyme may participate in other biological functions such as apoptosis and the regulation of cell growth. The nuclease activity is inhibited by ATP at low millimolar concentrations which suggests that the nuclease activity of PfNDK is inhibited under normal cell conditions where ATP is present in similar quantities. However, the level of ATP drops significantly during the early stages of programmed cell death (Valenti et al., 2007). Such new findings can be further investigated for development of new antiparasitic drugs.
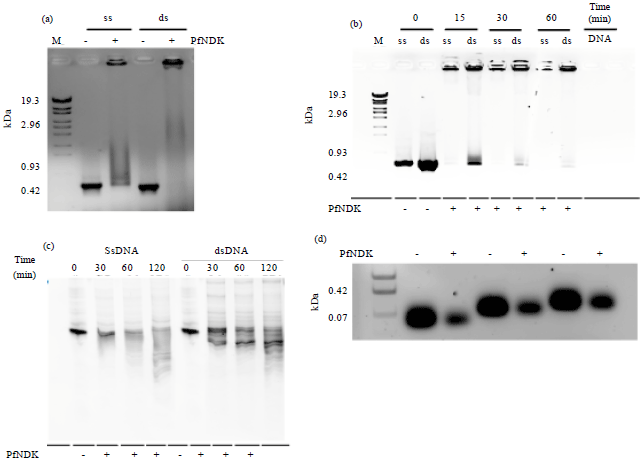  | |
| Fig. 2(a-e): | (a) Agarose gel showing the interaction of PfNDK with ssDNA or dsDNA. The 1 μg of ssDNA or dsDNA was incubated with 0 or 4 μg of PfNDK for 2 h. -: No addition of PfNDK, +: Adding 4 μg of PfNDK. M: DNA size marker, (b) Agarose gel showing the interaction of PfNDK with ssDNA or dsDNA. The 1 μg of ssDNA or dsDNA was incubated with 0 or 4 μg of PfNDK for 0, 15, 30 or 60 min. -: No addition of PfNDK, +: Adding 4 μg of PfNDK, (c) Polyacrylamide gel showing the interaction of PfNDK with ssDNA or dsDNA. The 1 μg of ssDNA or dsDNA was incubated with 0 or 4 μg of PfNDK for 0, 30 or 60 min. -: No addition of PfNDK, +: Adding 4 μg of PfNDK, (d) Agarose gel showing the interaction of PfNDK with different lengths of DNA substrate. The 2 μg of ssDNA was incubated with 0 or 4 μg of PfNDK for 2 h. -: No addition of PfNDK, +: Adding 4 μg of PfNDK and (e) Nuclease assay in the presence of 1 mM ATP. The 2 μg of PfNDK were incubated with 500 ng of ssDNA or dsDNA for 2 h in the presence or absence of 1 mM ATP. The density of bands was evaluated with ImageQuant 5.2. The relative peak area represents the size of each band compared with the control bands (absence of PfNDK). M: Molecular weight marker, ss: ssDNA and ds: dsDNA |
ABBREVIATIONS
| NDP | = | Nucleoside diphosphate |
| PfNDPK | = | Plasmodium falciparum nucleoside diphosphate kinase |
| NDPK | = | Nucleoside diphosphate kinase |
REFERENCES
- Choudhuri, T., M. Murakami, R. Kaul, S.K. Sahu and S. Mohanty et al., 2010. Nm23-h1 can induce cell cycle arrest and apoptosis in b cells. Cancer Biol. Ther., 9: 1065-1078.
PubMed - Fancsalszky, L., E. Monostori, Z. Farkas, E. Pourkarimi and N. Masoudi et al., 2014. Ndk-1, the homolog of nm23-h1/h2 regulates cell migration and apoptotic engulfment in C. elegans. PLoS One, Vol. 9.
CrossRef - Hammargren, J., T. Salinas, L. Marechal-Drouard and C. Knorpp, 2007. The pea mitochondrial nucleoside diphosphate kinase cleaves DNA and rna. FEBS Lett., 581: 3507-3511.
CrossRefPubMedDirect Link - Hasunuma, K., N. Yabe, Y. Yoshida, Y. Ogura and T. Hamada, 2003. Putative functions of nucleoside diphosphate kinase in plants and fungi. J. Bioenerg. Biomembr., 35: 57-65.
PubMed - Kandeel, M. and Y. Kitade, 2010. Substrate specificity and nucleotides binding properties of NM23H2/nucleoside diphosphate kinase homolog from Plasmodium falciparum. J. Bioenergetics Biomembr., 42: 361-369.
CrossRefDirect Link - Kandeel, M. and Y. Kitade, 2011. The substrate binding preferences of Plasmodium thymidylate kinase. Biol. Pharm. Bull., 34: 173-176.
CrossRefPubMedDirect Link - Miranda, M.R., G.E. Canepa, L.A. Bouvier and C.A. Pereira, 2008. Trypanosoma cruzi: Multiple nucleoside diphosphate kinase isoforms in a single cell. Exp. Parasitol., 120: 103-107.
CrossRefPubMedDirect Link - Neeld, D., Y. Jin, C. Bichsel, J. Jia and J. Guo et al., 2014. Pseudomonas aeruginosa injects NDK into host cells through a type III secretion system. Microbiology, 160: 1417-1426.
CrossRefDirect Link - Sikarwar, J., S. Kaushik, M. Sinha, P. Kaur, S. Sharma and T.P. Singh, 2013. Cloning, expression and purification of nucleoside diphosphate kinase from Acinetobacter baumannii. Enzyme Res., Vol. 2013.
CrossRefDirect Link - Sun, J., V. Singh, A. Lau, R.W. Stokes and A. Obregon-Henao et al., 2013. Mycobacterium tuberculosis nucleoside diphosphate kinase inactivates small gtpases leading to evasion of innate immunity. PLoS Pathog., Vol. 9.
CrossRefDirect Link - Valenti, D., R.A. Vacca, M.C. de Pinto, L. de Gara, E. Marra and S. Passarella, 2007. In the early phase of programmed cell death in tobacco bright yellow 2 cells the mitochondrial adenine nucleotide translocator, adenylate kinase and nucleoside diphosphate kinase are impaired in a reactive oxygen species-dependent manner. Biochim. Biophys. Acta, 1767: 66-78.
CrossRefPubMedDirect Link