
A. Das
Department of Animal Nutrition, College of Veterinary Science and Animal Husbandry, Orissa University of Agriculture and Technology, Bhubaneswar, Odisha, 751003, India
S.K. Mishra
Department of Animal Nutrition, College of Veterinary Science and Animal Husbandry, Orissa University of Agriculture and Technology, Bhubaneswar, Odisha, 751003, India
R.K. Swain
Department of Animal Nutrition, College of Veterinary Science and Animal Husbandry, Orissa University of Agriculture and Technology, Bhubaneswar, Odisha, 751003, India
G. Sahoo
Department of Veterinary Biochemistry, College of Veterinary Science and Animal Husbandry, Orissa University of Agriculture and Technology, Bhubaneswar, Odisha, 751003, India
N.C. Behura
Post Graduate Department of Poultry Science, College of Veterinary Science and Animal Husbandry, Orissa University of Agriculture and Technology, Bhubaneswar, Odisha, 751003, India
K. Sethi
Department of Animal Nutrition, College of Veterinary Science and Animal Husbandry, Orissa University of Agriculture and Technology, Bhubaneswar, Odisha, 751003, India
B. Chichilichi
Department of Livestock Production Management, College of Veterinary Science and Animal Husbandry, Orissa University of Agriculture and Technology, Bhubaneswar, Odisha, 751003, India
S.R. Mishra
Department of Animal Nutrition, College of Veterinary Science and Animal Husbandry, Orissa University of Agriculture and Technology, Bhubaneswar, Odisha, 751003, India
T. Behera
Fish Health Management Division, Central Institute of Freshwater Aquaculture, Bhubaneswar, Odisha, 751002, India
K. Dhama
Division of Pathology, Indian Veterinary Research Institute, Izatnagar, Bareilly (Uttar Pradesh), 243122, India
P. Swain
Fish Health Management Division, Central Institute of Freshwater Aquaculture, Bhubaneswar, Odisha, 751002, India
International Journal of Pharmacology
Year: 2014 | Volume: 10 | Issue: 5 | Page No.: 237-247







ABSTRACT
A comparative study on effect of replacement of inorganic minerals viz., Zinc (Zn), Copper (Cu) and Manganese (Mn) with their corresponding organic minerals (methionine) on growth, bioavailability and immunity in layer chicks was undertaken till 8th week post feeding. At 8th week, the body weight of birds were found to be significantly (p<0.05) higher in 100% organic Zn group, 100 and 50% organic Zn, Cu and Mn supplemented groups. The cumulative feed consumption and Feed Conversion Ratio (FCR) of all the treated groups showed no significant (p>0.05) difference. The serum glucose, cholesterol and ALP levels showed significant (p<0.05) differences. Significantly (p<0.05) higher titer levels were observed in 100% organic Zn group, 100% and 50% organic Zn, Cu and Mn supplementation groups. The CBH response of all the treated groups showed no significant (p>0.05) difference. At 8 weeks of age, the relative weight of spleen of layer chicks in 100% and 50% organic Zn, Cu and Mn supplementation groups were significantly (p<0.05) higher than all the other treatments. Tibia bone weight (g), tibia calcium (%) and tibia phosphorus (%) varied insignificantly (p>0.05). But significant (p<0.05) difference was observed with respect to the tibia ash content. Except Zn and Mn levels of tibia, all other studied mineral levels of serum and liver did not differ significantly (p>0.05). Faecal excretions of minerals were significantly lower in organic mineral fed groups. Replacement of inorganic Zn, Cu and Mn with corresponding organic minerals improved the body weight, immunity and lower faecal excretion of minerals in chicks.
PDF Abstract XML References Citation
Received: March 18, 2014;
Accepted: June 15, 2014;
Published: August 21, 2014
How to cite this article
A. Das, S.K. Mishra, R.K. Swain, G. Sahoo, N.C. Behura, K. Sethi, B. Chichilichi, S.R. Mishra, T. Behera, K. Dhama and P. Swain, 2014. Effects of Organic Minerals Supplementation on Growth, Bioavailability and Immunity in Layer Chicks. International Journal of Pharmacology, 10: 237-247.
DOI: 10.3923/ijp.2014.237.247
URL: https://scialert.net/abstract/?doi=ijp.2014.237.247
DOI: 10.3923/ijp.2014.237.247
URL: https://scialert.net/abstract/?doi=ijp.2014.237.247
INTRODUCTION
To meet the mineral requirements, Zinc (Zn), Copper (Cu) and Manganese (Mn) have been supplemented in poultry diets using inorganic salts such as oxides and sulphates. But the trace minerals tend to dissociate in the low pH environment of upper gastrointestinal tract, leaving the minerals susceptible to various nutrients and ingredient antagonisms that impair absorption (Underwood and Shuttle, 1999). Chelated minerals can be utilized at a much lower concentration in the diet than inorganic minerals, without a negative impact on production performance. Higher bioavailability of proteinates and amino acid chelates (Wedekind and Baker, 1990; Wedekind et al., 1992; Cao et al., 2000) along with lowered mineral load in manure has resulted in increased use of these forms of trace minerals in feeding of livestock and poultry. Abdallah et al. (2009) observed that chicks fed diets containing 100% organic minerals (Zn, Cu, Mn and Fe) had significantly higher body weight and better feed conversion on comparison with those of inorganic minerals. The effects of organic minerals are not limited to the growth, production or increased ash content of bone in layer birds; rather, they have some beneficial impact on immune system as well as in reduction of stress in the birds (Wei et al., 2001; Aksu et al., 2010). The variability of the results observed by the researchers posted questions on the validity of chelated mineral application in animal production. Also most of the studies, as reported, were carried out in either broilers or layer birds during laying phase and the output of those reported research work cannot be taken as a guideline for feeding of organic minerals in starter period of layers. Hence, the present study entitled has been planned with the objective of studying the effects of supplementation of organic minerals in on the performance of layer chicks.
MATERIALS AND METHODS
Animals, diet and experimental procedure: Vaccinated 1-day-old sexed commercial (BV 300) layer chicks (n = 120) were randomly allocated to six dietary treatments. Each treatment group had 2 replicates containing 10 chicks in each replicate. The chicks were randomly distributed so as to eliminate any significant difference between treatments with respect to body weight. The chicks were protected against New Castle and Infectious Bursal diseases by routine vaccination. Experimental diets were offered as per the specification of BV 300 from day old up to the end of experimental feeding period (0-8 weeks). The formulation of experimental diet is presented in Table 1. The chicks were provided 24 h free access to clean drinking water. From 0-4 weeks of age, artificial light was provided to chicks to achieve brooding temperature and further the day length was the photoperiod for the birds during chick and grower stage. Samples of the experimental feed was analysed for dry matter, crude protein, ether extract, crude fibre, total ash and acid insoluble ash as per AOAC (1995). Calcium was measured according to the method modified by Talapatra et al. (1940) and available phosphorus was determined as per IS: 1374-1968. The micro minerals content of the feed samples were estimated by using atomic absorption spectrophotometer.
The dietary treatments of the experiment were, T1: Basal diet (without Cu, Zn and Mn minerals) +80 ppm Zn, 15 ppm Cu and 80 ppm Mn supplementation from inorganic source in form of ZnSO4, CuSO4 and MnSO4, respectively, T2: T1 with replacement of 80 ppm inorganic Mn with 80 ppm Mn-methionate, T3: T1 with replacement of 15 ppm inorganic Cu with 15 ppm Cu-methionate, T4: T1 with replacement of 80 ppm inorganic Zn with 80 ppm Zn-methionate, T5: Basal diet+50% Zn (40 ppm), Cu (7.5) and Mn (40 ppm) supplementation from organic source (Zn-methionate, Cu-methionate and Mn-methionate) and T6: Basal diet+100% Zn (80 ppm), Cu (15) and Mn (80 ppm) supplementation from organic source (Zn-methionate, Cu-methionate and Mn-methionate).
Organic minerals source: The organic Zn, organic Cu and organic Mn minerals were obtained from S.A. Pharmaceuticals Pvt. Limited, Vadodara, Gujurat, India. The analytical report revealed the methionine activity in Organic Cu, Organic Zn and Organic Mn were 67.04, 73.13 and 73.60%, respectively. Similarly Zn, Cu and Mn activity were 12.67, 18.20 and 14.85%, respectively.
Growth performance: Initial live weight and weekly body weight of individual chicks were recorded for the calculation of weight gain. Daily feed offered to the birds were recorded group wise. The group average feed consumption was calculated by subtracting the left over feeds at the end of each week from the total feed supplied to the birds during the said weeks. Cumulative feed consumption was calculated by adding the feed consumption from 1st up to the desired week. The FCR was calculated using the following equation:
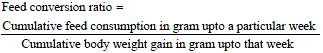 |
Biochemical analysis: The biochemical analysis of serum was carried out at 8 weeks of age.
| Table 1: | Formulation of experimental diet |
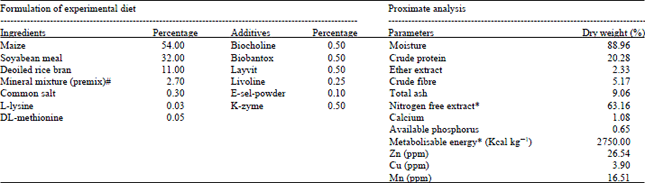 | |
| #: Mineral mixture-without Zn, Cu and Mn minerals, *Calculated value | |
Serum biochemical indices determined were serum glucose, urea, Alkaline phosphate (ALP), Serum Glutamate-Oxalloacetate Transaminase (SGOT or AST), Serum Glutamate-Pyruvate Transaminase (SGPT or ALP) and total protein. The serum Glucose, Cholesterol, Total Protein, Albumin, Globulin, Urea, BUN, Calcium, Phosphorus, Total Alkaline Phosphatase (ALP), SGOT, SGPT were determined by using Crest biosystems Kit (Goa, India).
Processing of immune organs and tibia bone: At the end of 8th week, 3 birds were randomly chosen from each treatment and slaughtered for collection of liver, tibia bone, spleen, bursa of fabricius and thymus. The birds were kept off fed overnight before bleeding and only water was provided. The live weight of the birds was recorded as pre slaughter weight. The broiler birds were bled by modified Kosher’s method (Panda and Mohapatra, 1989).
Tibia bone weight and mineral content: The tibia length (mm), weight (g), ash (%), Ca (%) and phosphorus (%) were determined to study the effect of dietary treatments on bone mineralization. The weight of the dried tibia was recorded with the help of electronic balance. Length and width of tibia was determined using electronic slide calliper. The tibia bones were ashed in a muffle furnace at 600°C for 4 h. The total ash was determined on percent weight basis. The tibia bone calcium was determined according to the method modified by Talapatra et al. (1940) and available phosphorus was determined as per IS: 1374-1968.
Micro mineral content in liver and tibia: The collected liver samples were oven dried at 100°C for 24 h and finely ground. The micro mineral content in the liver samples were determined by collecting 0.5 g samples and digesting at 120°C with 5 mL concentrate HNO3 for 1 h using KEL plus digestion system. The digested samples were cooled and further digested with 30% H2O2 at 200°C. The process continued until the content appeared clear and colourless. The digested samples were filtered into a volumetric flask. The contents of digestion tubes were repeatedly washed with triple distilled water to obtain complete extract of the mineral. For determination of tibia micro mineral content, 0.2 g of ash samples were solubilized in 5 mL of 50% HCl and the mineral extract was filtered into a volumetric flask. The extract was then diluted using triple distilled water to the required volume and the micro mineral concentration was determined by using atomic absorption spectrophotometer.
Measure of cellular immunity: At 8th week, two birds from each replicate in each dietary treatment were injected intra-dermally in the comb with 100 μg of Phytohaemaglutinin-P (PHAP) in 0.1 mL of normal saline to measure the cellular immune response by Cutaneous Basophilic Hypersensitivity (CBH) test (Edelman et al., 1986). The thickness of comb was measured using digital calliper before inoculation and 24 h post inoculation and CBH response was calculated using the equation:
Measure of humoral immunity: The measure of humoral immunity was carried out as per the method described by Abdallah et al. (2009). Sheep Red Blood Cells (SRBC) were used as test antigens to quantitatively analyse specific antibody response as measure of humoral immunity. Antibody production to SRBC was measured using microtitration haemagglutination technique with microtiter plate U shape of 96 wells (8 rowsx12 column) according to Bachman and Mashaly (1986) and Kai et al. (1988). All SRBC antibody titres were expressed as log2 of the reciprocal of the highest serum dilution causing agglutination of SRBC.
Collection of faeces: The faeces of the experimental chicks and grower birds were collected at 8th week of age. Three birds of each group were taken for individual collection of faeces. A polythene sheet was attached under the cages of the birds and light was turn off for 1 h. The faeces were homogeneously mixed replicate wise and representative samples of the faeces were collected in moisture cup and were oven dried at 105°C for 24 h. For determination of micro mineral content in faeces, 2 g of faecal samples were taken in a digestion tube and to it 12 mL of triacid mixture (7 mL HNO3, 3 mL H2SO4 and 2 mL Perchloric acid) was added and digested at 200°C. The process continued until the content appeared clear and colorless. The digested samples were filtered into a volumetric flask. The content of digestion tubes were repeatedly washed with triple distilled water to obtain complete extract of the mineral.
Statistical analysis: The statistical analysis of the experimental data was done according to Snedecor and Cochran (1998).
RESULTS
Body weight: Growth performance and feed utilization of layer chicks with different mineral sources is presented in the Table 2. The average body weights (g) of BV 300 layer chicks at the end of 8th week were 488.85±10.62, 479.80±13.16, 485.90±14.37, 526.55±18.14, 514.50±16.16 and 531.55±13.30 in the treatments T1, T2, T3, T4, T5 and T6, respectively.
| Table 2: | Growth performance and feed utilization of layer chicks with different mineral sources (inorganic and organic) |
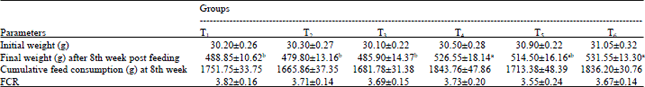 | |
| FCR: Feed conversion ratio, results were presented as Mean±SE of triplicate observations. Values bearing different superscripts in a row differ significantly (p<0.05) | |
| Table 3: | Biochemical profile of layer chicks with different mineral sources (inorganic and organic) |
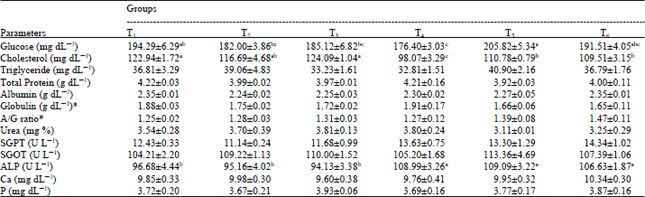 | |
| Results were presented as Mean±SE of triplicate observations. Values bearing different superscripts in a row differ significantly (p<0.05) | |
| Table 4: | Immunity status of layer chicks with different mineral sources (inorganic and organic) |
 | |
| Results were presented as Mean±SE of triplicate observations. Values bearing different superscripts in a row differ significantly (p<0.05) | |
| Table 5: | Lymphoid organ weight (% of live weight) of layer chicks with different mineral sources (inorganic and organic) |
 | |
| Results were presented as Mean±SE of triplicate observations. Values bearing different superscripts in a row differ significantly (p<0.05) | |
At 8th week, group T6 was found to be significantly (p<0.05) higher with respect to average body weight as compared to groups T1, T2 and T3. But the body weight of chicks of T4, T5 and T6 groups did not differ significantly (p>0.05). The cumulative feed consumption and FCR of all the treated groups up to 8th week of age did not differ significantly (p>0.05). However, best FCR of 3.55 was observed in treatment T5 that received 50% of organic minerals followed by T6.
Biochemical profile: Serum biochemical parameters viz., glucose, cholesterol, triglyceric acid, total protein, albumin, globulin, urea, BUN, SGPT, SGOT, ALP, calcium and phosphorus at eight week BV 300 layer chicks are presented in the Table 3. The serum triglyceride, total protein, albumin, globulin, urea, SGPT, SGOT, Ca and P in serum of the layer birds varied insignificantly (p>0.05) between the treatments whereas, serum glucose, cholesterol and ALP levels in the blood serum showed significant (p<0.05) differences.
Immunity status: Antibody titer against SRBC and CBH response against PHA-P and weight of lymphoid organs, were used as measures to study the immunity status of the layer birds under different dietary treatments. The antibody titer against SRBC and CBH response against PHA-P and weight of lymphoid organs are presented in Table 4 and Table 5, respectively. Significantly (p<0.05) higher titer levels were observed in the treatments T6 (7.677±0.49), T4 (7.66±0.60) and T5 (7.17± 0.60) when compared to that of treatments T1 and T2.
| Table 6: | Tibia bone parameters of layer chicks with different mineral sources (inorganic and organic) |
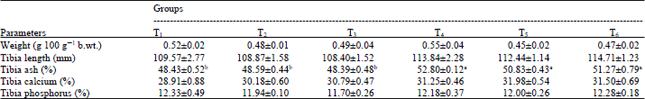 | |
| Results were presented as Mean±SE of triplicate observations. Values bearing different superscripts in a row differ significantly (p<0.05) | |
| Table 7: | Average minerals concentration (ppm) in different organs, serum and faeces of layer chicks with different mineral sources (inorganic and organic) |
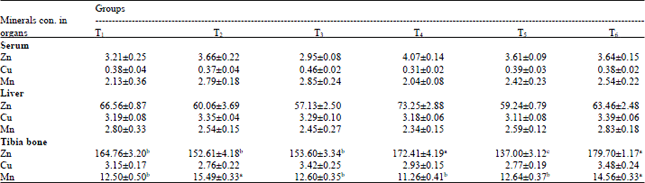 | |
| Results were presented as Mean±SE of triplicate observations. Values bearing different superscripts in a row differ significantly (p<0.05) | |
| Table 8: | Mineral content (ppm) in the faeces of layer chicks |
 | |
| Results were presented as Mean±SE of triplicate observations. Values bearing different superscripts in a row differ significantly (p<0.05) | |
The CBH response of all the treated groups ranged from 112.51±2.70 (T2) to 152.84±11.44 (T4) without any significant (p>0.05) difference between the treated groups. The relative weight of spleen of layers in T6 was significantly (p<0.05) higher than all the other treatments except T5. The percentage weight of bursa was found to be the highest in the birds of treatment T5 which varied insignificantly (p>0.05). Significantly higher (p<0.05) relative weight of thymus was observed in the layers of T6 when compared to that of T1, T2 and T3.
Tibia bone weight and mineral content: The tibia bone weight (g), tibia ash (%), tibia calcium (%) and tibia phosphorus (%) of 8 weeks old layer chicks are presented in the Table 6. Tibia bone weight (g), tibia calcium (%) and tibia phosphorus (%) varied insignificantly (p>0.05). But significant (p<0.05) difference was observed with respect to the tibia ash content.
Bioavailability: Measuring the deposition or storage of minerals in selected tissues (tibia or plasma Zn, liver Cu and tibia Mn) is the most common output in trace mineral relative bioavailability experiments (Underwood and Shuttle, 1999). The average mineral concentration (ppm) in different organs and serum of BV 300 layer chicks under different dietary treatments is presented in Table 7. The mineral levels in liver and serum of the experimental birds of all the treated groups did not differ significantly (p>0.05). The tibia Zn level of T5 was observed to be significantly (p<0.05) lower than that of other treated groups. The tibia Mn level of T2 was observed to be significantly (p<0.05) higher than that of all treated groups except T6. At 8 weeks of age, except Zn and Mn levels of tibia, all other mineral levels differed non-significantly (p>0.05).
Faecal excretion: The average mineral concentration (ppm) in faeces of BV 300 layer chicks under different dietary treatments is presented in Table 8. At 8 weeks of age, the faecal Zn (ppm) excretion was significantly (p<0.05) higher in T1 (273.52±10.59), T2 (262.17±6.74) and T3 (260.04±10.39) than that of other treated groups. The faecal excretion of Cu was found to be significantly (p<0.05) higher in T1 (63.74±2.88), T2 (62.27±1.54) and T4 (70.82±4.35) than that of other treated groups. Similarly, faecal excretion of Mn was found to be significantly (p<0.05) higher in T1 (307.69±8.62) and T3 (290.08±10.00) than that of other treated groups.
DISCUSSION
The earlier studies on organic mineral feeding in poultry are mostly reported in broiler birds and during the laying stage of layers and not many references are available for layer chicks. So the discussion of this experimental finding is mainly based on the available literature on broiler and layers. From 5-8 weeks of age, the body weight of organic Zn supplemented group (T4) was significantly higher than inorganic Zn supplemented group (T1). Though the sources of Zn in both T4 and T1 were different but the Zn level in both the groups were same. In spite of same level of Zn feeding, in the present study, significantly (p<0.05) higher body weight was recorded in T4 than that of T1. This corroborated with the findings of Abdallah et al. (2009) who reported that the body of weight chicks at 35th day fed with 100% organic Zn were significantly higher than the group fed with 50% organic and 50% inorganic Zn and group fed with 100% inorganic Zn. Similar findings were also reported by Koreleski et al. (2001). The significantly higher body weight observed in T4 might be due to improved bioavailability of Zn amino acid chelates (Spears, 1989; Wedekind et al., 1992). Organic minerals are good vehicles to supply broilers with more trace minerals without increasing the dietary minerals levels (Abdallah et al., 2009). The 8th week body weight of layer chicks of T1, T2 and T3 did not differ significantly which implied that replacement of inorganic Mn and Cu with respective organic minerals did not have any significant (p>0.05) effect on body weight. These results are in accordance with (Banks et al., 2004; Zhao et al., 2010), who reported no significant difference in body weight in poultry by replacing inorganic Cu at 50 and 100% level.
However, the results contradicted the findings of Paik et al. (1999) who reported that broilers fed Cu-Met (organic) had increased daily gain and feed intake. The body weight of layer chicks of T4 and T6 groups recorded significantly (p<0.05) higher body weight at 8th week of age from T1, T2 and T3. The body weight of layer chicks of T5 group did not differ significantly from other treated groups. This was in agreement with the findings of Abdallah et al. (2009). They reported lower body weight in 100% inorganic mineral fed groups as compared to feeding of organic minerals to broiler birds. The lower body weight in T1 might be due to the reason that use of organic mineral sources can improve intestinal absorption of trace elements as they reduce interference from agents that form insoluble complexes with the ionic trace elements (Van der Klis and Kemme, 2002). On comparison of the body weight of T5 (50% Zn, Cu and Mn organic mineral) with T6 (100% Zn, Cu and Mn organic minerals), the body weight of both the groups did not differ significantly at 8th week. This is in agreement with the findings of Bao et al. (2007) who reported that medium level of organic minerals supplementation had higher body weight gain than that of high or low level of inorganic mineral supplementation. Cumulative feed consumption throughout the study period did not show any significant (p>0.05) difference among the treatment groups. Similarly, the FCR, of all the groups from 1st to 8th weeks of age did not differ significantly. On summarization, it was observed that, supplementation of organic minerals either alone or combination at 50 or 100% level in layer chick diet had no significant effect on Feed Consumption and FCR. Similar findings was recorded by Aksu et al. (2011) who reported that the feed consumption and feed conversion were not influenced by replacement of 1/3, 2/3 and 3/3 levels of inorganic Zn, Cu and Mn with organic sources in broiler ration. This finding is contrast to the findings as reported by Abdallah et al. (2009). The FCR of 50% level of inclusion of organic trace minerals (Zn, Cu and Mn) (T5) did not differ significantly (p>0.05) from their respective mineral inclusion at 100% level (T6) in the diet of experimental layer chicks. This is in agreement with the findings of Zhao et al. (2010).
The serum triglyceride, total protein, albumin, globulin, albumin and globulin ratio, urea, SGOT and SGPT, calcium and phosphorus levels of all the treated groups of layer chicks did not differ significantly (p>0.05). Feng et al. (2010) in their experiments in broiler on feeding of organic Zn reported non-significant difference in serum albumin level. Non-significant levels of serum calcium and phosphorus was reported by Parak and Strakova (2011) while comparing feeding of inorganic with organic Zn in breeding cocks which corroborated with the present finding. In contrast to this, Al-Daraji and Amen (2011) reported significantly higher levels of serum calcium and phosphorus on increasing Zn concentration in the diet by addition of 100 mg of Zn per kg of diet than that of control (basal diet) in broiler breeders from 58-66 weeks of age. The significant increase of serum calcium and phosphorus as reported by Al-Daraji and Amen (2011) might be due to higher level of Zn in the diet. Osman and Ragab (2007) also observed significant (p<0.05) effects of Zn-Met supplemented at 30, 40 and 50 ppm on serum SGPT level and Idowu et al. (2011) observed significant (p<0.05) difference in serum protein and uric acid concentrations between control and Zn-proteinate groups with higher levels in Zn-proteinate group which is in contrast to our findings. The serum glucose, cholesterol and ALP levels of all the treated groups of layer chicks differed significantly (p<0.05). The serum glucose and serum cholesterol levels were significantly (p<0.05) lower in T4 (Organic Zn fed group) than T1 (inorganic mineral fed group) and T5 (50% organic mineral fed group). This observed cholesterol level is in agreement with the finding of Aksu et al. (2010); Also, Cu, Zn and Mn are related to lipid metabolism. Lu and Combs (1988) reported that inorganic Zn did not affect the serum cholesterol level. On contrary (Boukaiba et al., 1993; Uyanik et al., 2001) indicated that inorganic Zn decreased the serum cholesterol level. In our study, a significant variation in the levels of serum cholesterol of layer chicks was found in case of Zn supplemented groups than the control. Herzig et al. (2009) proved that there was a significant decrease of plasma cholesterol when broilers were fed with high amounts of Zn in diet. It was also observed by Parak and Strakova (2011) while comparing feeding of inorganic with organic Zn in breeding cocks. It revealed that concentrations of plasma cholesterol are influenced by Zn (ZnO) supplementation either alone or in combination with Cu or vitamins (Age-Related Eye Disease Study Research Group, 2002). Our results along with the different research findings confirmed the positive impact of Zn on lipid metabolism. Non- significant levels of serum glucose was reported by Parak and Strakova (2011) while comparing feeding of inorganic with organic Zn in breeding cocks which is in contradiction to our finding. The ALP level in the serum of organic Zn supplemented groups (T4, T5 and T6) was found to be significantly (p<0.05) higher from the inorganic Zn supplemented group. Idowu et al. (2011) also observed significant difference in the levels of serum ALP and serum Zn concentrations between control and Zn proteinate groups with higher levels in Zn proteinate and opined that due to Zn binding capacity of serum, alkaline phosphatase acts as good indicator of Zn status. Al-Daraji and Amen (2011) reported significantly higher levels of serum ALP level on increasing Zn concentration in the diet by addition of 100 mg pure Zn to a kg of diet than that of control (basal diet) in broiler breeders from 58-66 weeks of age. The increase in ALP level on Zn supplementation might be due to increase in corticosteroid hormone, epinephrine and nor-epinephrine secretion (Al-Daraji and Amen, 2011). In contrast to this, non significant level of serum ALP in organic Zn fed groups was reported by Aksu et al. (2010) on feeding different levels of organic Zn, Cu and Mn in broiler ration.
Higher antibody titers against SRBC were observed in layer chicks in organic mineral supplementation in form of Zn alone or combination of Zn, Cu and Mn when compared to the control T1. Chicks fed on 100% organic-Zn were recorded the highest value of average antibody titer and the control group fed inorganic mineral recorded the lowest value. Increasing the levels of organic minerals from 50-100% had no significant effect on the antibodies titer. Beach et al. (1980) reported that diets supplemented with Zn tended to improve the ability of birds to produce antibodies. Stahl et al. (1989) reported that Zn as Zn-methionine supplementation (100 mg kg-1 Zn to a basal diet containing 36.8 mg kg-1 Zn) had better effect on primary immune response to SRBC relative to control. Richards et al. (2006) tested the antibody response to vaccination against coccidiosis. They reported that birds supplemented with organic Zn or organic Cu exhibited a significantly improvement in immune response and intestinal health benefits of broiler. The better immune response as observed in T5 and T6 might be due to the fact that antagonism between Zn and Cu can be avoided through using chelated forms of these minerals (Abdallah et al., 2009). The layer chicks provided diets supplemented with Zn from Zn amino acid might have increased thymulin activity; therefore, enhancing immune response through increased maturation of T-lymphocyte and activation of B lymphocytes by T-helper cells (Hudson et al., 2005). Though the CBH response of all the treated groups did not differ significantly but numerically higher CBH response was observed in the treatment T4. Sunder et al. (2008) reported that the birds supplemented with diets without any Zn elicited the lowest CBH response. Hudson et al. (2005) reported that immune response to PHA-P injection was enhanced when dietary Zn supplementation was solely from Zn-AA. Cellular immune response of the birds to PHA-P injection was enhanced and transferred to their progenies when the diets were supplemented with Zn-Met in place of dietary addition of Zn in the form of Zn sulphate (Kidd et al., 1992, 2000).
Except bursa, significantly higher weights of lymphoid organs (percentage of live weight) were observed in the treatment T4, T5 and T6 groups. Low levels of supplemental Zn showed a relative reduction in the size of lymphoid organs with the possible decrease in T-cell function (Kidd et al., 1996). Idowu et al. (2011) observed significant higher weight of spleen in Zn-proteanate group than the control group. Earlier researchers in this context reported that, a diet deficient in Zn leads to atrophy of the thymus (Prasad and Oberleas, 1971) and a reduction in spleen weight of rats (Mengheri et al., 1988). Similar results were also reported when hens were provided diets with Zn from amino acid complexes or chelates that had increased relative thymus weights when compared to their inorganic counterparts (Virden et al., 2004; Feng et al., 2010; Moghaddam and Jahanian, 2009). In contrast to the present results, Moghaddam and Jahanian (2009) and Feng et al. (2010) reported non- significant effect on the weight of spleen on organic mineral supplementation. But they reported non-significant effect on bursa of Fabricius which corroborated to present finding. Dietary supplementation with a more bioavailable Zn source like Zn-Met caused a significant increase in relative weight of thymus. In contrast to this, Bartlett and Smith (2003) reported that none of these studied organs (thymus, bursa and spleen) were significantly affected by the level of Zn in the diet. Even if insignificant, birds on a high Zn diet showed a slight increase in relative weight of thymus when compared with lower levels of Zn supplementation. Moghaddam and Jahanian (2009) reported that dietary addition of organic Zn in place of inorganic source in broiler diet increased the relative weight of thymus as a proportion of live body weight but no significant effect on the weight of spleen and bursa of fabricius. They opined that more dietary Zn or a more bio-available Zn source is needed to assure functionality of the immune system, particularly cell- mediated immune response. The lower weight of lymphoid organs observed in treatment T3 might be due to antagonism between Zn and Cu as Cu was supplemented in organic and Zn was in organic form (Abdallah et al., 2009).
The tibia wt (g), tibia length (mm), tibia Ca (%) and tibia P (%) of all the treatments did not differ significantly. But significant (p<0.05) difference was observed with respect to the tibia ash content. The tibia ash content was observed to be significantly higher in organic Zn (T4) and 100% organic Zn, Cu and Mn (T6) mineral supplemented group than inorganic mineral group (T1). Similar results were also reported by Abdallah et al. (2009) who reported that supplementation of organic Zn, Cu and Mn at 50 and 100% levels in the diet of broilers did not have any significant effect on the tibia wt (g), tibia length (mm), tibia Ca (%) and tibia P (%) levels but they observed significantly (p<0.05) higher levels of tibia ash content in organic mineral supplemented groups. Zhao et al. (2010) and Britanico et al. (2012) had also the similar findings as regard to tibia mineral concentration. Swiatkiewicz and Koreleski (2008) in their experiment reported that organic Zn supplementation had no significant effect on tibia length, relative weight of tibia which corroborated with our findings but they observed non-significant effect on tibia ash content and of the laying birds which contradicts to our finding.
The tibia bone from chicks fed the diet containing 100% organic Zn had significantly higher Zn content than those from chicks fed the diet containing inorganic Zn. These results are consistent with published data by Ledoux et al. (1991), Miles et al. (1998), Ao et al. (2006), Lim and Paik (2006) and Shelton and Southern (2006). Organic Zn source caused more Zn accumulation in tibia than inorganic Zn source (Ao et al., 2009). Trace mineral concentrations in the liver of birds were almost similar inspite of organic mineral supplementation. This was because chickens give priorities to their mineral requirements for vital functions in compromise of body growth as indicated by the normal concentrations of the minerals in the plasma of the control birds (Bao et al., 2007). Bao et al. (2007) reported that adding Cu to the control diet had no effect on total Cu, Fe, Mn and Zn contents of tibia bone which is in agreement with the present finding. With Zn, the results are consistent with the model for laboratory animals (King, 1990) which showed that if dietary deficiency of Zn was mild (as in case of T5), the animal usually reduced their rate of growth and excretion to maintain normal tissue concentrations.
As regard to excretion of trace minerals concerned, the excretion of Zn, Cu and Mn were significantly higher in inorganic mineral fed groups than organic mineral fed groups even at same level of feeding. This was because organically complexed trace minerals provide alternative pathways for absorption, thus leading to a reduction in the excretion of minerals (Leeson, 2003). Burrell et al. (2004) reported that the cumulative Zn excretion of organic Zn fed groups were significantly lower than inorganic Zn fed groups. Sacranie (2003) reported that the total amount of inorganic minerals in a broiler premix could be totally replaced by 20% organic minerals without affecting growth performance and at the same time reducing environmental pollution. The mineral excretion of group T5 (50% organic mineral supplemented group) was significantly (p<0.05) lower than that of other treated groups. This implied that at 50% level of organic mineral supplementation, the bio-availability of minerals to cater the need of the body was higher than feeding at 100% level. This result is also in agreement with human Zn research in which there appears to be an adaptation to low-Zn intake associated with a reduction in Zn excretion to conserve this micro-nutrient and maintain normal plasma Zn concentration (Sullivan et al., 1998). Abdallah et al. (2009) also reported the similar result.
CONCLUSION
Replacement of inorganic Zn with organic Zn at 100% level or replacement inorganic Zn, Cu and Mn with organic Zn, Cu and Mn either at 50 or 100% level improved the body weight and immunity of layer chicks. Bio-availability, on the basis of tibia, liver and serum concentration, was same for Cu and Mn. But tibia Zn concentration was better in 100% organic Zn and 100% organic minerals supplementation. Faecal excretions of organic minerals were lower than inorganic minerals.
REFERENCES
- Abdallah, A.G., O.M. El-Husseiny and K.O. Abdel-Latif, 2009. Influence of some dietary organic mineral supplementations on broiler performance. Int. J. Poult. Sci., 8: 291-298.
CrossRefDirect Link - Aksu, D.S., T. Aksu and B. Ozsoy, 2010. The effects of lower supplementation levels of organically complexed minerals (zinc, copper and manganese) versus inorganic forms on hematological and biochemical parameters in broilers. Kafkas Universitesi Veteriner Fakultesi Dergisi, 16: 553-559.
Direct Link - Aksu, T., M.I. Aksu, M.A. Yoruk and M. Karaoglu, 2011. Effects of organically-complexed minerals on meat quality in chickens. Br. Poult. Sci., 52: 558-563.
CrossRefDirect Link - Al-Daraji, H.J. and M.H.M. Amen, 2011. Effect of dietary zinc on certain blood traits of broiler breeder chickens. Int. J. Poult. Sci., 10: 807-813.
CrossRefDirect Link - Ao, T., J.L. Pierce, R. Power, K.A. Dawson, A.J. Pescatore, A.H. Cantor and M.J. Ford, 2006. Evaluation of bioplex Zn® as an organic zinc source for chicks. Int. J. Poult. Sci., 5: 808-811.
CrossRefDirect Link - Ao, T., J.L. Pierce, R. Power, A.J. Pescatore, A.H. Cantor, K.A. Dawson and M.J. Ford, 2009. Effects of feeding different forms of zinc and copper on the performance and tissue mineral content of chicks. Poult. Sci., 88: 2171-2175.
CrossRefPubMedDirect Link - AOAC., 1995. Official Methods of Analysis. 16th Edn., Association of Official Analytical Chemists, Washington, DC., USA.
Direct Link - Banks, K.M., K.L. Thompson, J.K. Rush and T.J. Applegate, 2004. Effects of copper source on phosphorus retention in broiler chicks and laying hens. Poult. Sci., 83: 990-996.
CrossRefDirect Link - Bachman, S.E. and M.M. Mashaly, 1986. Relationship between circulating thyroid hormones and humoral immunity in immature male chickens. Dev. Comp. Immunol., 10: 395-403.
CrossRefPubMedDirect Link - Bao, Y.M., M. Choct, P.A. Iji and K. Bruerton, 2007. Effect of organically complexed copper, iron, manganese, and zinc on broiler performance, mineral excretion, and accumulation in tissues. J. Appl. Poult. Res., 16: 448-455.
CrossRefDirect Link - Bartlett, J.R. and M.O. Smith, 2003. Effects of different levels of zinc on the performance and immunocompetence of broilers under heat stress. Poult. Sci., 82: 1580-1588.
CrossRefDirect Link - Beach, R.H., M.E. Gershwin, R.K. Makishima and L.S. Hurley, 1980. Impaired immunologic ontogeny in postnatal zinc deprivation. J. Nutr., 110: 805-810.
Direct Link - Boukaiba, N., C. Flament, S. Acher, P. Chappuis and A. Piau et al., 1993. A physiological amount of zinc supplementation: Effects on nutritional, lipid and thymic status in an elderly population. Am. J. Clin. Nutr., 57: 566-572.
Direct Link - Britanico, E.B., F.E. Merca, A.A. Angeles, S.P. Acda and E.S. Luis, 2012. Effects of supplementing diets with amino acid chelates of copper, zinc, manganese and iron on the performance of broilers. Philipp. J. Vet. Anim. Sci., 38: 1-10.
Direct Link - Burrell, A.L., W.A. Dozier, A.J. Davis, M.M. Compton, M.E. Freeman, P.F. Vendrell and T.L. Ward, 2004. Responses of broilers to dietary zinc concentrations and sources in relation to environmental implications. Br. Poult. Sci., 45: 255-263.
CrossRefDirect Link - Cao, J., P.R. Henry, R. Guo, R.A. Holwerda and J.P. Toth et al., 2000. Chemical characteristics and relative bioavailability of supplemental organic zinc sources for poultry and ruminants. J. Anim. Sci., 78: 2039-2054.
Direct Link - Edelman, A.S., P.L. Sanchez, M.E. Robinson, G.M. Hochwald and G.J. Thorbecke, 1986. Primary and secondary wattle swelling response to phytohemagglutinin as a measure of immunocompetence in chickens. Avian Dis., 30: 105-111.
Direct Link - Feng, J., W.Q. Ma, H.H. Niu, X.M. Wu, Y. Wang and J. Feng, 2010. Effects of zinc glycine chelate on growth, hematological and immunological characteristics in broilers. Biol. Trace Elem. Res., 133: 203-211.
CrossRefDirect Link - Age-Related Eye Disease Study Research Group, 2002. The effect of five-year zinc supplementation on serum zinc, serum cholesterol and hematocrit in persons randomly assigned to treatment group in the age-related eye disease study: AREDS Report No. 7. J. Nutr., 132: 697-702.
PubMedDirect Link - Herzig, I., M. Navratilova, J. Totusek, P. Suchy, V. Vecerek, J. Blahova and Z. Zraly, 2009. The effect of humic acid on zinc accumulation in chicken broiler tissues. Czech J. Anim. Sci., 54: 121-127.
Direct Link - Hudson, B.P., W.A. Dozier III and J.L. Wilson, 2005. Broiler live performance response to dietary zinc source and the influence of zinc supplementation in broiler breeder diets. Anim. Feed Sci. Technol., 118: 329-335.
CrossRefDirect Link - Idowu, O.M.O., R.O. Ajuwon, A.O. Oso and O.A. Akinloye, 2011. Effects of zinc supplementation on laying performance, serum chemistry and Zn residue in tibia bone, liver, excreta and egg shell of laying hens. Int. J. Poult. Sci., 10: 225-230.
CrossRefDirect Link - Wei, J.P.J., C. Srinivasan, H. Han, J.S. Valentine and E.B. Gralla, 2001. Evidence for a novel role of copper-zinc superoxide dismutase in zinc metabolism. J. Biol. Chem., 276: 44798-44803.
CrossRefDirect Link - Kidd, M.T., N.B. Anthony and S.R. Lee, 1992. Progeny performance when dams and chicks are fed supplemental zinc. Poult. Sci., 71: 1201-1206.
CrossRefDirect Link - Kidd, M.T., P.R. Ferket and M.A. Qureshi, 1996. Zinc metabolism with special reference to its role in immunity. World's Poult. Sci., 52: 309-323.
CrossRefDirect Link - Kidd, M.T., M.A. Qureshi, P.R. Ferket and L.N. Thomas, 2000. Turkey hen zinc source affects progeny immunity and disease resistance. J. Applied Poult. Res., 9: 414-423.
CrossRefDirect Link - Ledoux, D.R., P.R. Henry, C.B. Ammerman, P.V. Rao and R.D. Miles, 1991. Estimation of the relative bioavailability of inorganic copper sources for chicks using tissue uptake of copper. J. Anim. Sci., 69: 215-222.
Direct Link - Leeson, S., 2003. A New Look at Trace Mineral Nutrition of Poultry, Can We Reduce Environmental Burden of Poultry Manure? In: Nutritional Biotechnology in the Feed and Food Industries: Proceedings of Alltech's Twentieth Annual Symposium, Lyons T.P. and K.A. Jacques (Eds.). Nottingham University Press, UK., ISBN: 9781904761273, pp: 125-129.
- Lim, H.S. and I.K. Paik, 2006. Effects of dietary supplementation of copper chelates in the form of methionine, chitosan and yeast in laying hens. Asian-Aust. J. Anim. Sci., 19: 1174-1179.
Direct Link - Lu, J. and G.F. Combs Jr., 1988. Effect of excess dietary zinc on pancreatic exocrine function in the chick. J. Nutr., 118: 681-689.
PubMedDirect Link - Mengheri, E., G. Bises and S. Gaetani, 1988. Differentiated cell-mediated immune response in zinc deficiency and in protein malnutrition. Nutr. Res., 8: 801-812.
CrossRefDirect Link - Miles, R.D., S.F. O'Keefe, P.R. Henry, C.B. Ammerman and X.G. Luo, 1998. The effect of dietary supplementation with copper sulfate or tribasic copper chloride on broiler performance, relative copper bioavailability and dietary prooxidant activity. Poult. Sci., 77: 416-425.
CrossRefDirect Link - Moghaddam, H.N. and R. Jahanian, 2009. Immunological responses of broiler chicks can be modulated by dietary supplementation of zinc-methionine in place of inorganic zinc sources. Asian-Aust. J. Anim. Sci., 22: 396-403.
CrossRefDirect Link - Paik, I.K., S.H. Seo, J.S. Um, M.B. Chang and B.H. Lee, 1999. Effects of supplementary copper-chelate on the performance and cholesterol level in plasma and breast muscle of broiler chickens. Asian-Aust. J. Anim. Sci., 12: 794-798.
Direct Link - Parak, T. and E. Strakova, 2011. Zinc as a feed supplement and its impact on plasma cholesterol concentrations in breeding cocks. Acta Veterinaria Brno, 80: 281-285.
CrossRefDirect Link - Prasad, A.S. and D. Oberleas, 1971. Changes in activities of zinc-dependent enzymes in zinc-deficient tissues of rats. J. Applied Physiol., 31: 842-846.
Direct Link - Richards, J., T. Hampton, C. Wuelling, M. Wehmeyer and J.J. Dibner, 2006. Mintrex Zn and mintrex Cu organic trace minerals improve intestinal strength and immune responses to coccidiosis infection and/or vaccination in broilers. Proceedings of the International Poultry Scientific Forum, January 23-24, 2006, Atlanta, GA., pp: 257-259.
- Shelton, J.L. and L.L. Southern, 2006. Effects of phytase addition with or without a trace mineral premix on growth performance, bone response variables and tissue mineral concentrations in commercial broilers. J. Applied Poult. Res., 15: 94-102.
CrossRefDirect Link - Spears, J.W., 1989. Zinc methionine for ruminants: Relative bioavailability of zinc in lambs and effects of growth and performance of growing heifers. J. Anim. Sci., 67: 835-843.
Direct Link - Stahl, J.L., J.L. Greger and M.E. Cook, 1989. Zinc, copper and iron utilisation by chicks fed various concentrations of zinc. Br. Poult. Sci., 30: 123-134.
CrossRefPubMedDirect Link - Sullivan, V.K., F.R. Burnett and R.J. Cousins, 1998. Metallothionein expression is increased in monocytes and erythrocytes of young men during zinc supplementation. J. Nutr., 128: 707-713.
Direct Link - Sunder, G.S., A.K. Panda, N.C.S. Gopinath, S.V.R. Rao, M.V.L.N. Raju, M.R. Reddy and C.V. Kumar, 2008. Effects of higher levels of zinc supplementation on performance, mineral availability and immune competence in broiler chickens. J. Applied Poult. Res., 17: 79-86.
CrossRefDirect Link - Swiatkiewicz, S. and J. Koreleski, 2008. The effect of zinc and manganese source in the diet for laying hens on eggshell and bones quality. Vet. Med., 53: 555-563.
Direct Link - Uyanik, F., M. Eren and G. Tuncoku, 2001. Effects of supplemental zinc on growth, serum glucose, cholesterol, enzymes and minerals in broilers. Pak. J. Biol. Sci., 4: 745-747.
CrossRefDirect Link - Virden, W.S., J.B. Yeatman, S.J. Barber, K.O. Willeford and T.L. Ward et al., 2004. Immune system and cardiac functions of progeny chicks from dams fed diets differing in zinc and manganese level and source. Poult. Sci., 83: 344-351.
CrossRefDirect Link - Wedekind, K.J. and D.H. Baker, 1990. Zinc bioavailability in feed-grade sources of zinc. J. Anim. Sci., 68: 684-689.
Direct Link - Wedekind, K.J., A.E. Hortin and D.H. Baker, 1992. Methodology for assessing zinc bioavailability: Efficacy estimates for zinc-methionine, zinc sulfate and zinc oxide. J. Anim. Sci., 70: 178-187.
PubMedDirect Link - Zhao, J., R.B. Shirley, M. Vazquez-Anon, J.J. Dibner and J.D. Richards et al., 2010. Effects of chelated trace minerals on growth performance, breast meat yield, and footpad health in commercial meat broilers. J. Appl. Poult. Res., 19: 365-372.
CrossRefDirect Link