
Kil-Nam Kim
Department of Marine Bio Research, Korea Basic Science Institute (KBSI), Jeju, 690-140, Republic of Korea
Jihyeon Kim
Jeju Biodiversity Research Institute, Jeju Technopark, Jeju, 699-943, Republic of Korea
Weon-Jong Yoon
Jeju Biodiversity Research Institute, Jeju Technopark, Jeju, 699-943, Republic of Korea
Hye-Mi Yang
Department of Marine Bio Research, Korea Basic Science Institute (KBSI), Jeju, 690-140, Republic of Korea
Soo Yeon Heo
Department of Marine Bio Research, Korea Basic Science Institute (KBSI), Jeju, 690-140, Republic of Korea
Ji-Yeon Ko
Department of Marine Bio Research, Korea Basic Science Institute (KBSI), Jeju, 690-140, Republic of Korea
Seong Woon Roh
Department of Marine Bio Research, Korea Basic Science Institute (KBSI), Jeju, 690-140, Republic of Korea
You-Jin Jeon
School of Marine Biomedical Sciences, Jeju National University, Jeju, 690-756, Republic of Korea
Sung-Myung Kang
School of Marine Biomedical Sciences, Jeju National University, Jeju, 690-756, Republic of Korea
Soo-Jin Heo
Global Bioresources Research Center, Korea Institute of Ocean Science and Technology, Ansan, 426-744, Republic of Korea
Kalpa W. Samarakoon
School of Marine Biomedical Sciences, Jeju National University, Jeju, 690-756, Republic of Korea
Dong-Sam Kim
Research and Education Institution of World Organic Agriculture and Fisheries, Samcheok, 245-802, Republic of Korea
Daekyung Kim
Department of Marine Bio Research, Korea Basic Science Institute (KBSI), Jeju, 690-140, Republic of Korea
International Journal of Pharmacology
Year: 2013 | Volume: 9 | Issue: 8 | Page No.: 524-532







ABSTRACT
Many seaweed species are used as traditional medicine in different parts of the world. However, data on the anti-inflammatory and anti-melanogenic effects of seaweeds are scarce. This study investigated the inhibitory activity of ethyl acetate extracts of Sargassum patens (ESP) on the production of inflammatory cytokines and melanin synthesis in cultured cell lines in vitro. ESP significantly inhibited the production of Nitric Oxide (NO), prostaglandin E2 (PGE2), interleukin (IL)-6 and tumor necrosis factor (TNF-α) on the lipopolysaccharide (LPS)-stimulated RAW264.7 cells. In addition, ESP suppressed the protein expressions of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) in LPS-stimulated RAW264.7 cells. ESP markedly inhibited α-melanocyte stimulating hormone (α-MSH)-induced melanin synthesis and tyrosinase activity and also decreased α-MSH-induced expression of microphthalmia-associated transcription factor (MITF), tyrosinase, tyrosinase-related protein (TRP)-1 and TRP-2 in B16 F10 cells. Furthermore, High Performance Liquid Chromatography (HPLC) fingerprinting of ESP revealed that fucoxanthin was the major component of these extracts.
PDF Abstract XML References Citation
Received: February 18, 2014;
Accepted: April 08, 2014;
Published: May 13, 2014
How to cite this article
Kil-Nam Kim, Jihyeon Kim, Weon-Jong Yoon, Hye-Mi Yang, Soo Yeon Heo, Ji-Yeon Ko, Seong Woon Roh, You-Jin Jeon, Sung-Myung Kang, Soo-Jin Heo, Kalpa W. Samarakoon, Dong-Sam Kim and Daekyung Kim, 2013. Inhibitory Effect of Sargassum patens on Inflammation and Melanogenesis. International Journal of Pharmacology, 9: 524-532.
DOI: 10.3923/ijp.2013.524.532
URL: https://scialert.net/abstract/?doi=ijp.2013.524.532
DOI: 10.3923/ijp.2013.524.532
URL: https://scialert.net/abstract/?doi=ijp.2013.524.532
INTRODUCTION
Seaweeds or their extracts have long been used as novel sources of various biologically acitive compounds having potential medicinal value (Balboa et al., 2013). Recently, considerable attention has been paid to the anti-tumor, anticholesterolemic and antioxidant activities of seaweed constituents. Seaweeds are commonly classified into 3 groups on the basis of their pigmentation: Brown (Phaeophyceae), red (Rhodophyceae) and green (Chlorophyceae) seaweeds.
Brown algae are under-exploited plant resources that are known to produce various secondary metabolites with different carbon skeletons (Blunt et al., 2006). In particular, Sargassum, a genus of brown seaweed in the Sargassaceae family, contains approximately 400 species (Mattio and Payri, 2011). Sargassum is found across all oceans and consumed as food and medicine in many cultures. Currently, about 200 bioactive compounds such as meroterpenoids, carotenoids, phlorotannins, fucoidans, sterols and glycolipids have been identified from this genus (Heo et al., 2011; Liu et al., 2012). These compounds have been reported to possess biological activities, including anticancer, antibacterial, antifungal, antiviral, anti-inflammatory, anticoagulant, antioxidant, hypoglycemic, hypolipidemic, antimelanogenic, anti-bone loss, hepatoprotective and neuroprotective activities (Balboa et al., 2013; Heo et al., 2011; Kim et al., 2010a; Liu et al., 2012). However, the anti-inflammatory and anti-melanogenesis activities of one of the species belonging to this genus, Sargassum patens have not yet been studied. Hence, in the present study, ethyl acetate extracts were obtained from S. patens (ESP) and their anti-inflammatory and anti-melanogenesis properties were assessed.
MATERIALS AND METHODS
Reagents: Dimethylsulfoxide (DMSO), 3-(4,5-dimethylthiazol-2-yl) 2,5-diphenyltetrazolium bromide (MTT), L-DOPA, alpha-melanocyte stimulating hormone (α-MSH) and Phosphate Buffered Saline (PBS) and lipopolysaccharide (LPS) were purchased from Sigma-Aldrich (St. Louis, MO, USA). ELISA kits for PGE2, TNF-α, IL-1β and IL-6 were purchased from R and D Systems, Inc. (St. Louis, MO, USA) and BD Biosciences (San Diego, CA, USA). Dulbecco’s modified Eagle’s medium (DMEM) and Fetal Bovine Serum (FBS) were obtained from Invitrogen-Gibco (Grand Island, NY, USA). Anti-inducible nitric oxide synthase (iNOS) and anti-cyclooxygenase-2 (COX-2) were purchased from Cell Signaling Technology (Beverly, MA, USA). Antibodies against microphthalmia-associated transcription factor (MITF), tyrosinase, tyrosinase-related protein (TRP)-1 and TRP-2 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). All other reagents were purchased from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA).
Preparation of ESP: S. patens was collected in Jeju Island on March 2010 and was then identified by Dong-Sam Kim. The samples were washed three times in tap water to remove the salt, epiphytes and sand attached to its surface, then rinsed carefully in fresh water and then dried at room temperature for 2 weeks. The dried samples were lyophilized and homogenized with a grinder prior to extraction. The powdered S. patens (100 g) was extracted with 80% ethanol (EtOH; 2L) at room temperature for 24 h and then evaporated in a vacuum. The evaporated EtOH extract was then suspended in water (4 L) and partitioned with ethyl acetate (EtOAc; 4L); this portioning was repeated 3 times.
Cell culture: The RAW 264.7 murine macrophage cells and B16F10 mouse melanoma cells were obtained from ATCC (American Type Culture Collection, Manassas, VA, USA). RAW 264.7 and B16F10 cells were cultured in DMEM supplemented with 100 U mL-1 of penicillin, 100 μg mL-1 of streptomycin and 10% FBS. The cells were then incubated in an atmosphere of 5% CO2 at 37°C and were sub-cultured every 3 days.
Lactate dehydrogenase (LDH) cytotoxicity assay: RAW 264.7 cells (1.5x105 cells mL-1) plated on 96-well plates were pre-incubated and subsequently treated with LPS (1 μg mL-1) coupled with aliquots of ESP at 37°C for 24 h. The medium was carefully removed from each well and the LDH activity in the medium was determined using an LDH cytotoxicity detection kit (Promega, Madison, WI, USA). In brief, 100 μL of reaction mixture were added to each well and the reaction was incubated for 30 min at room temperature in darkness. The absorbance of each well was measured at 490 nm with a UV spectrophotometer.
Determination of NO production: After a 24 h pre-incubation of RAW 264.7 cells (1.5x105 cells mL-1) with LPS (1 μg mL-1), the quantity of nitrite accumulated in the culture medium was measured as an indicator of NO production. In brief, 100 μL of cell culture medium was mixed with 100 μL of Griess reagent (1% sulfanilamide and 0.1% naphthylethylenediamine dihydrochloride in 2.5% phosphoric acid), the mixture was incubated at room temperature for 10 min and the absorbance at 540 nm was measured in a microplate reader. Fresh culture medium employed as a blank in every experiment. The quantity of nitrite determined from a sodium nitrite standard curve.
Measurement of PGE2 and pro-inflammatory cytokines (IL-6 and TNF-α) production: The inhibitory effect ESP on prostaglandin E2 (PGE2), interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) production in LPS-treated RAW264.7 cells was determined as previously described (Kim et al., 2013b). Supernatants were then harvested and assayed for PGE2 and cytokines by ELISA.
Measurement of cellular melanin contents: Cellular melanin contents were measured using a previously described method (Yoon et al., 2010). The cells (2x104 cells mL-1) were stimulated with α-MSH (50 nM) and then incubated with ESP (0, 12.5, 25 and 50 μg mL-1) for 72 h; the cells were then washed in ice-cold PBS. Briefly, the cells were incubated at 80°C for 1 h in 1 mL of 1N NaOH/10% DMSO and then vortexed to solubilize the melanin: the absorbance at 450 nm was then measured.
Measurement of cellular tyrosinase activity: Cellular tyrosinase activity was measured according to the previously reported method with slight modifications (Tomita et al., 1992). Briefly, the cells were cultured at 2x104 cells mL-1 in 24 well plates. The 16 h after cells seeding, the cells (2x104 cells mL-1) were stimulated with α-MSH (50 nM) and then incubated with ESP (0, 12.5, 25 and 50 μg mL-1) for 72 h. The cells were washed with PBS and lysed in PBS containing 1% Triton X-100 by freezing and thawing. The lysates were clarified by centrifugation at 13000 g for 10 min. After protein quantification and normalization, 90 μL of cell lyate (each sample contained the same amount of protein) was incubated in duplicate with 10 μL of 10 mM L-DOPA at 37°C for 1 h. After incubation, dopachrom was monitored by measuring the absorbance at 405 nm using and ELISA reader. The value of each measurement is expressed as percentage change from the control.
Western blot analysis: The effect of ESP on the expression of pro-inflammatory mediators and melanogenesis-related protein such as iNOS, COX-2, tyrosinase, TRP-1, TRP-2 and MITF were assessed by Western blot analysis. Western blot was performed as described previously (Kim et al., 2012). Briefly, nuclear proteins (40 μg) loaded into SDS-PAGE were electro-transferred onto nitrocellulose transfer membranes. After blocking with 5% nonfat milk for 2 h, blot were incubated with iNOS (1:1000 dilution), COX-2 (1:1000 dilution), tyrosinase (1:300 dilution), TRP1 (1:1000 dilution), TRP2 (1:1000 dilution), MITF (1:1000 dilution), or β-actin (1:3000 dilution) antibodies. β-actin was used as an internal control. Visualization was achieved by using ECL reagents.
HPLC-fingerprint analysis: HPLC analysis were carried out using an ACCELA system (ThermoFisher Scientific, USA), which included pump, auto sampler, column oven and PDA detector, connected to Xcalibur software. A Hypersil-Gold C18 column (100x2.1 mm i.d., 1.9 um) from Thermo was used. The column temperature was maintained at 25°C. The standards and samples were separated using an isocratic mobile phase consisting of 80% methanol. The flow rate was set at 0.2 mL min-1 and the injection volume was 5 μL. The detection wavelength was set at 450 nm. Identification and quantification of fucoxanthin was based on retention time when co-injected with standards.
Statistical analysis: All data are expressed as Means±S.D. Significant differences between the groups were determined using the unpaired Student’s t-test. A value of *p<0.05 was considered to be statistically significant.
RESULTS
Effect of ESP on NO and PGE2 production in LPS-stimulated RAW264.7 cells: The potential anti-inflammatory properties of ESP were investigated using RAW264.7 murine macrophage cells, which can produce NO and PGE2 upon stimulation with LPS. Cells were pre-incubated with ESP for 1 h, following which they were stimulated with 1 μg mL-1 LPS for 24 h. Both LPS and sample were not treated in control cells. LPS alone was able to markedly induce higher NO and PGE2 production in the cells than that noted in control cells. However, ESP inhibited NO and PGE2 production in LPS-stimulated RAW264.7 cells in a concentration-dependent manner (Fig. 1a, b). In addition, ESP did not show cytotoxic effects on RAW 264.7 cells in the concentration range of 12.5-50 μg mL-1 (Fig. 1a). Thus, doses of 12.5-50 μg mL-1 were selected for use in further experiments.
Effect of ESP on expression of iNOS and COX-2 proteins in LPS-stimulated RAW 264.7 cells: Further determine whether inhibition of NO and PGE2 production were associated with decreased levels of iNOS and COX-2 was determined by performing western blot analysis for the detection of protein levels. Untreated cells did not induce iNOS and COX-2 protein expressions, while LPS induced their expressions. However, ESP attenuated LPS-induced expressions of iNOS and COX-2 proteins in the cells (Fig. 2). Thus, the effects of EPS on the expression of the iNOS and COX-2 enzymes, which generate NO and PGE2 as key mediators of inflammation, respectively, were evaluated in greater detail.
Effect of ESP on IL-6 and TNF-α production in LPS-stimulated RAW264.7 cells: Next, whether ESP suppresses the production of pro-inflammatory cytokines, IL-6 and TNF-α in LPS-stimulated RAW264.7 cells was investigated. The cells were incubated with ESP in the presence or absence of LPS for 24 h and cytokine levels in the culture media were measured. IL-6 was produced by treatment of LPS, which was inhibited to around 90% by the addition of ESP at 50 μg mL-1 and LPS-induced IL-6 production decreased in a dose-dependent manner after treatment with ESP (Fig. 3a). Moreover, the results of the TNF-α assay suggested that 50 μg mL-1 ESP significantly suppressed (25%) LPS-induced TNF-α production (Fig. 3b). Although ESP had an inhibitory effect on IL-6 production, the effect was not as strong as that exhibited during the inhibition of TNF-α production.
Effect of ESP on tyrosinase activity and melanin synthesis in α-MSH-stimulated B16F10 cells: The hypopigmentary effect of ESP was investigated by performing melanin content and tyrosinase activity assays in α-MSH-stimulated B16F10 cells. ESP significantly inhibited both melanin production and tyrosinase activity in a concentration-dependent manner (Fig. 4). In addition, MTT assay results suggested that there was no cytotoxicity effect of ESP on B16F10 cells at the tested concentration (data not shown). These results suggested that ESP inhibits α-MSH-induced melanin synthesis with down-regulation of tyrosinase activity in B16F10 cells.
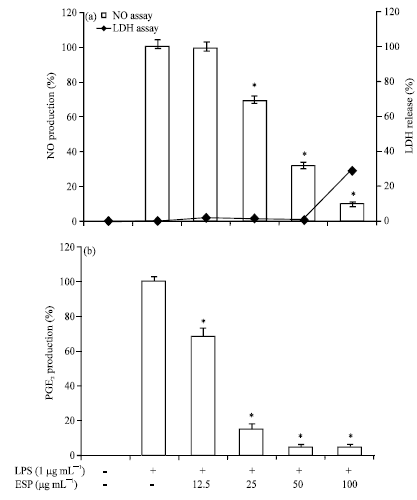 | |
| Fig. 1(a-b): | Effects of ethyl acetate extracts of S. patens (ESP) on (a) Nitric oxide (NO) and (b) prostaglandin E2 (PGE2) production in lipopolysaccharide (LPS)-induced RAW264.7 cells. Cells were pretreated for 1h with different concentrations (12.5, 25, 50 and 100 μg mL-1) of ESP and then LPS (1 μg mL-1) was added and incubated for 24 h, (a) Cytotoxicity was determined using the Lactate dehydrogenase (LDH) method, values are expressed as Means±S.D of triplicate experiments, *p<0.05 indicate significant differences from the LPS-stimulated group. Parameters on x-axis denote as follows, LPS (-): Without LPS-treatment; LPS (+): LPS-treated cells; ESP (-): Without EPS treatment; ESP 12.5, 25, 50 and 100: 12.5, 25, 50 and 100 μg mL-1 ESP-treated cells, respectively |
Effect of ESP on the expression of tyrosinase, TRP-1, TRP-2 and MITF proteins in α-MSH-stimulated B16F10 cells: Next whether ESP could modulate melanogenesis-related protein expression under α-MSH stimulation was investigated. B16F10 cells were exposed to ESP in the presence of α-MSH and the changes in the levels of tyrosinase, TRP-1, TRP-2 and MITF expression were analyzed by western blot analysis. B16F10 cells treated with ESP showed significantly decreased tyrosinase, TRP-1, TRP-2 and MITF protein expression levels compared with those of the α-MSH group (Fig. 5). These results indicated that ESP decreases melanin synthesis via the down-regulation of TRP and MITF expression.
HPLC fingerprint of ESP: According to traditional oriental medicine, the therapeutic actions of herbal medicines are based on integral interaction of many ingredients combined. With the development of analytical technology, chromatographic methods are highly recommended for developing fingerprints of traditional oriental medicine and their raw materials. Thus, interest in HPLC fingerprint analysis has increased in not only Asia but also other countries worldwide (Kim et al., 2009; Yoon et al., 2009; Zhang et al., 2007). Therefore, simple HPLC fingerprints of the algae were explored in this study. Since fucoxanthin has been reported to be an ingredient in the Sargassum species that has effective anti-inflammatory and anti-melanogenesis effects, this compound was used as a standard substance.
 | |
| Fig. 2: | Effects of ethyl acetate extracts of S. patens (ESP) on lipopolysaccharide (LPS)-induced inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) protein expressions in RAW 264.7 cells. Cells were treated for 1h with different concentrations (12.5, 25 and 50 μg mL-1) of ESP, LPS (1 μg mL-1) was then added and cells were incubated for 24 h. iNOS and COX-2 protein level were determined via Western blotting and β-actin was used as an internal control |
The conditions described in the methods section led to better resolution of fucoxanthin from ESP, with excellent peak shapes. The contents in the ESP were 11.2 mg g-1 (Fig. 6).
DISCCUSION
Sargassum species extracted using various solvents showed anti-inflammatory, anti-allergic, antimicrobial, antiviral, antiplasmodial, anticancer, liver protective, gastric protective, bone protective, skin-whitening, anti-alzheimer and antioxidant activities (Liu et al., 2012). This is the first report of evaluation of the anti-inflammatory and anti-melanogenesis activities of S. patens in RAW264.7 cells and B16F10 cells, respectively.
Inflammatory action is an essential aspect of host response to bacterial and viral infections and injury and is required for the maintenance of good health. However, chronic and uncontrolled inflammation has been implicated as the pathophysiological mechanism underlying many chronic diseases, including diabetes, cardiovascular disease, cancer, arthritis and neurodegenerative diseases (Lawrence et al., 2002; Libby, 2007).
Macrophages actively participate in inflammatory responses by releasing pro-inflammatory cytokines (TNF-α and IL-6) and inflammatory factors (NO and PGE2) that recruit additional immune cells to sites of infection or tissue injury (Bosca et al., 2005; Kim et al., 2010a).
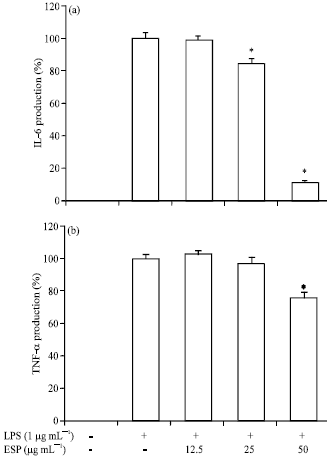 | |
| Fig. 3(a-b): | Inhibitory effect of ethyl acetate extracts of S. patens (ESP) on the pro-inflammatory cytokine production in RAW264.7 cells. The production of (a) IL-6 and (b) TNF-α were assayed in the culture medium of cells stimulated with lipopolysaccharide (LPS) (1 μg mL-1) for 24 h in the presence of ESP (12.5, 25 and 50 μg mL-1). Supernatants were collected and the tumor necrosis factor (TNF-α) and interleukin (IL)-6 concentration in the supernatants were determined by ELISA, values are expressed as Means±S.D of triplicate experiments. *p<0.05 indicate significant differences from the LPS: Stimulated group, LPS (-): Without LPS-treatment; LPS (+): LPS-treated cells, ESP (-): Without EPS treatment; ESP 12.5, 25 and 50: 12.5, 25 and 50 μg mL-1 ESP-treated cells, respectively |
The production of these inflammatory molecules by macrophages can be induced in response to LPS stimulation.
NO is an important regulatory molecule in a range of physiological and pathological processes(Ajizian et al., 1999; Kim et al., 2013b).
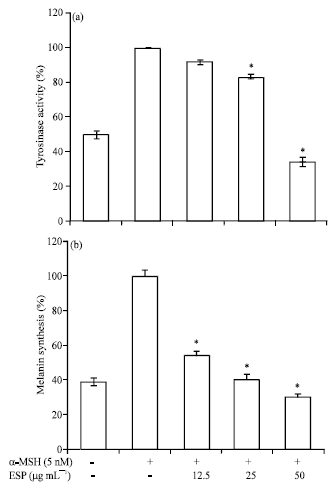 | |
| Fig. 4(a-b): | Effect of ethyl acetate extracts of S. patens (ESP) on cellular (a) Tyrosinase activity and (b) Melanin synthesis in B16F10 cells, cells were exposed to 50 nM α-melanocyte stimulating hormone (α-MSH) in the presence of 12.5-50 μg mL-1 ESP. The percentage values of the treated cells are expressed relatively compared to that in the control cells. Data are reported as the Mean±S.D of three independent experiments carried out in triplicate. *p<0.05 indicate significant differences from the the α-MSH treated one. α-MSH (-): Without α-MSH-treatment, α-MSH (+): α-MSH-treated cells; ESP (-): Without EPS treatment; ESP 12.5, 25 and 50: 12.5, 25 and 50 μg mL-1 ESP-treated cells, respectively |
It is generated by 3 isoforms of NO Synthase (NOS): endothelial NOS (eNOS), neuronal NOS (nNOS) and iNOS (Nathan and Xie, 1994). iNOS is expressed in response to various inflammatory stimuli and generates high levels of NO in macrophages during the inflammatory process (Laskin and Pendino, 1995).
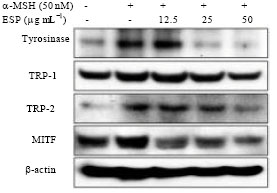 | |
| Fig. 5: | Effect of ethyl acetate extracts of S. patens (ESP) on the expression of melanogenesis-related proteins in B16F10 cells. Cells were exposed to 50 nM α-melanocyte stimulating hormone (α-MSH) in the presence of 12.5-50 μM ESP for 72 h. The expression levels of the microphthalmia-associated transcription factor (MITF), tyrosinase, tyrosinase-related protein (TRP)-1 and TRP-2 proteins were examined by Western blot and β-actin was used as an internal control |
PGE2 is also a well-known inflammatory mediator derived from arachidonic acid via the action of cyclooxygenases. Growth factors, cytokines and pro-inflammatory molecules-mediated overproduction of PGE2 is known to be associated with the up-regulation of COX-2 (Minghetti, 2004; St-Onge et al., 2007). The results obtained in this study suggested that ESP inhibits NO and PGE2 production via the suppression of iNOS and COX-2 expression, respectively.
Furthermore, ESP also reduced the LPS-induced production of TNF-α and IL-6. Pro-inflammatory cytokines such as TNF-α, IL-1β and IL-6 are known to contribute to tissue damage and multiple organ failure. They are considered to be important initiators of inflammatory response and mediators of the development of various inflammatory diseases (Glauser, 1996; Mannel and Echtenacher, 2000). Moreover, the production of TNF-α is crucially required for the synergistic induction of NO synthesis in LPS-stimulated macrophages (Aggarwal and Natarajan, 1996). IL-6 is a cytokine released by LPS-activated monocytes and plays a crucial role in the immune response (Ghosh and Karin, 2002). Therefore, anti-inflammatory drugs are developed using inhibitors of these inflammatory molecules (Andreakos et al., 2004) to alleviate various disorders caused by the activation of macrophages.
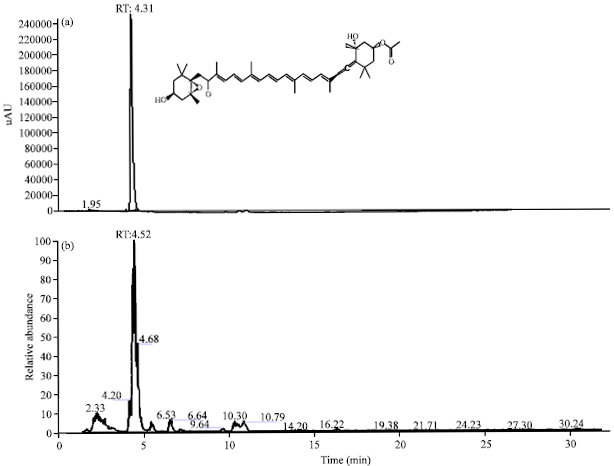 | |
| Fig. 6(a-b): | High-performance liquid chromatography (HPLC) fingerprinting analysis of the ethyl acetate extracts of S. patens (ESP), HPLC profiles of (a) Authentic fucoxanthin and (b) ESP, respectively, the wavelength of fucoxanthin is 440 nm |
Melanogenesis is a physiological process resulting in the production of melanin pigment, which plays an important role in the prevention of sun-induced skin injury and contributes to skin and hair color (Gilchrest and Eller, 1999; Sturm et al., 1998). However, the accumulation of excess epidermal melanin can induce cosmetic problems, such as melasma, lentigo and age spots and cause various hyperpigmentation disorders such as malignant melanoma (Gupta et al., 2006; Narayanan et al., 2010). Therefore, many plant parts have been investigated to determine their potential for use as anti-melanogenesis agents.
Melanogenesis is directly regulated by melanogenic enzymes such as tyrosinase, TRP-1 and TRP-2. Tyrosinase is a rate-limiting enzyme of melanogenesis, which first catalyzes the hydroxylation of tyrosine to 3,4-dihydroxyphenylalanine (DOPA) and then catalyzes the oxidation of DOPA to DOPA-quinone. TRP-1 oxidizes 5,6-dihydroxyindole-2-carboxylic acid (DHICA) to a carboxylated indole-quinone (Hearing and Jimenez, 1987). Tyrosinase and TRP-1 complex is important for tyrosinase activation and stability (Kobayashi et al., 1998). TRP-2, which functions as a DOPA-chrome tautomerase, catalyzes the rearrangement of DOPA-chrome to DHICA (Park et al., 2009). These 3 enzymes have been recognized as the most critical regulators of melanin biosynthesis. The tyrosinase family genes tyrosinase, TRP-1 and TRP-2 responsible for pigmentation are transcriptionally regulated by microphthalmia-associated transcription factor (MITF) (Levy et al., 2006; Tachibana, 2001; Ye et al., 2010). In the present study, ESP was found to markedly inhibit α-MSH-induced melanin synthesis and tyrosinase activity in a dose-dependent manner and these doses were not cytotoxic to B16F10 cells. Further, ESP significantly suppressed the protein levels of tyrosinase, TRP-1, TRP-2 and MITF in α-MSH-stimulated B16F10 cells.
Sargassum is widely distributed in tropical areas and is reported to possess diverse compounds, including triterpenoids, carotenoids, phlorotannins, fucoidans, sterols and glycolipids, with various biological activities (Heo et al., 2011; Liu et al., 2012). Among the carotenoids, fucoxanthin-one of the major xanthophylls in brown alga-has been identified in several Sargassum species. Recent pharmacological research indicates that fucoxanthin has anticancer, anti-pigmentary, anti-inflammatory, anti-oxidant, anti-obesity and antidiabetic activities (Kim et al., 2013a; Kim et al., 2010a; Kim et al., 2010b; Liu et al., 2012; Shimoda et al., 2010). Sargassum contains 1-8 mg of fucoxanthin per gram dry weight, with seasonal variation (Terasaki et al., 2009). In the present study, ESP showed high fucoxanthin contents (11.2 mg g-1). These facts suggest the anti-inflammatory and anti-melanogenesis effects of ESP might primarily be attributed to fucoxanthin.
CONCLUSION
In conclusion, ESP was found to significantly attenuate the release of inflammatory mediators (NO, PGE2, TNF-α and IL-6) in LPS-stimulated RAW264.7 cells. Additionally, ESP inhibited cellular melanin synthesis and tyrosinase activity by suppressing MITF, tyrosinase, TRP-1 and TRP-2 in α-MSH-stimulated B16F10 cells. Further studies are necessary to isolate the bioactive compounds from ESP in order to elucidate the mechanism of melanogenesis and inflammation inhibition.
ACKNOWLEDGMENT
This study was supported by the project fund (C34290) to D. Kim from Korea Basic Science Institute (KBSI).
REFERENCES
- Aggarwal, B.B. and K. Natarajan, 1996. Tumor necrosis factors: Developments during the last decade. Eur. Cytokine Network, 7: 93-124.
PubMed - Ajizian, S.J., B.K. English and E.A. Meals, 1999. Specific inhibitors of p38 and extracellular Signal-regulated kinase Mitogen-activated protein kinase pathways block inducible nitric oxide synthase and tumor necrosis factor accumulation in murine macrophages stimulated with lipopolysaccharide and Interferon-γ. J. Infect. Dis., 179: 939-944.
CrossRefPubMedDirect Link - Andreakos, E., B. Foxwell and M. Feldmann, 2004. Is targeting Toll-like receptors and their signaling pathway a useful therapeutic approach to modulating Cytokine-driven inflammation? Immunol. Rev., 202: 250-265.
CrossRef - Balboa, E.M., E. Conde, A. Moure, E. Falque and H. Dominguez, 2013. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem., 138: 1764-1785.
CrossRef - Blunt, J.W., B.R. Copp, M.H.G. Munro, P.T. Northcote and M.R. Prinsep, 2006. Marine natural products. Nat. Prod. Rep., 23: 26-78.
CrossRefDirect Link - Bosca, L., M. Zeini, P.G. Traves and S. Hortelano, 2005. Nitric oxide and cell viability in inflammatory cells: A role for NO in macrophage function and fate. Toxicology, 208: 249-258.
CrossRefDirect Link - Ghosh, S. and M. Karin, 2002. Missing pieces in the NF-κB puzzle. Cell, 109: S81-S96.
CrossRefPubMedDirect Link - Gilchrest, B.A. and M.S. Eller, 1999. DNA photodamage stimulates melanogenesis and other photoprotective responses. J. Invest. Dermatol. Symp. Proc., 4: 35-40.
PubMed - Glauser, M.P., 1996. The inflammatory cytokines. New developments in the pathophysiology and treatment of septic shock. Drugs, 52: 9-17.
PubMed - Gupta, A.K., M.D. Gover, K. Nouri and S. Taylor, 2006. The treatment of melasma: A review of clinical trials. J. Am. Acad. Dermatol., 55: 1048-1065.
CrossRef - Hearing, V.J. and M. Jimenez, 1987. Mammalian tyrosinase-the critical regulatory control point in melanocyte pigmentation. Int. J. Biochem., 19: 1141-1147.
CrossRefDirect Link - Heo, S.J., K.N. Kim, W.J. Yoon, C. Oh and Y.U. Choi et al., 2011. Chromene induces apoptosis via caspase-3 activation in human leukemia HL-60 cells. Food Chem. Toxicol., 49: 1998-2004.
CrossRef - Kim, K.N., G. Ahn, S.J. Heo, S.M. Kang and M.C. Kang et al., 2013. Inhibition of tumor growth in vitro and in vivo by fucoxanthin against melanoma B16F10 cells. Environ. Toxicol. Pharmacol., 35: 39-46.
CrossRef - Kim, K.N., Y.M. Ham, J.Y. Moon, M.J. Kim and Y.H. Jung et al., 2012. Acanthoic acid induces cell apoptosis through activation of the p38 MAPK pathway in HL-60 human promyelocytic leukaemia. Food Chem., 135: 2112-2117.
CrossRef - Kim, K.N., Y.M. Ham, J.Y. Moon, M.J. Kim and D.S. Kim et al., 2009. In vitro cytotoxic activity of Sargassum thunbergii and Dictyopteris divaricata (Jeju Seaweeds) on the HL-60 tumour cell line. Int. J. Pharmacol., 9: 298-306.
CrossRefDirect Link - Kim, K.N., S.J. Heo, S.M. Kang, G. Ahn and Y.J. Jeon, 2010. Fucoxanthin induces apoptosis in human leukemia HL-60 cells through a ROS-mediated Bcl-xL pathway. Toxicol. In vitro, 24: 1648-1654.
CrossRefPubMedDirect Link - Kim, K.N., S.J. Heo, W.J. Yoon, S.M. Kang, G. Ahn, T.H. Yi and Y.J. Jeon, 2010. Fucoxanthin inhibits the inflammatory response by suppressing the activation of NF-κB and MAPKs in lipopolysaccharide-induced RAW 264.7 macrophages. Eur. J. Pharmacol., 649: 369-375.
CrossRefPubMedDirect Link - Kim, K.N., Y.J. Ko, M.C. Kang, H.M. Yang and S.W. Roh et al., 2013. Anti-inflammatory effects of trans-1,3-diphenyl-2,3-epoxypropane-1-one mediated by suppression of inflammatory mediators in LPS-stimulated RAW 264.7 macrophages. Food Chem. Toxicol., 53: 371-375.
CrossRef - Kobayashi, T., G. Imokawa, D.C. Bennett and V.J. Hearing, 1998. Tyrosinase stabilization by Tyrp1 (the brown locus protein). J. Biol. Chem., 273: 31801-31805.
PubMed - Laskin, D.L. and K.J. Pendino, 1995. Macrophages and inflammatory mediators in tissue injury. Annu. Rev. Pharmacol. Toxicol., 35: 655-677.
CrossRef - Lawrence, T., D.A. Willoughby and D.W. Gilroy, 2002. Anti-inflammatory lipid mediators and insights into the resolution of inflammation. Nat. Rev. Immunol., 2: 787-795.
CrossRefDirect Link - Levy, C., M. Khaled and D.E. Fisher, 2006. MITF: Master regulator of melanocyte development and melanoma oncogene. Trends Mol. Med., 12: 406-414.
CrossRefDirect Link - Libby, P., 2007. Inflammatory mechanisms: The molecular basis of inflammation and disease. Nutr. Rev., 65: S140-S146.
CrossRefDirect Link - Liu, L., M. Heinrich, S. Myers and S.A. Dworjanyn, 2012. Towards a better understanding of medicinal uses of the brown seaweed Sargassum in traditional Chinese medicine: A phytochemical and pharmacological review. J. Ethnopharmacol., 142: 591-619.
CrossRef - Mannel, D.N. and B. Echtenacher, 2000. TNF in the inflammatory response. Chem. Immunol., 74: 141-161.
PubMed - Mattio, L. and C.E. Payri, 2011. 190 years of Sargassum taxonomy, facing the advent of DNA phylogenies. Bot. Rev., 77: 31-70.
CrossRef - Minghetti, L., 2004. Cyclooxygenase-2 (COX-2) in inflammatory and degenerative brain diseases. J. Neuropathol. Exp. Neural., 63: 901-910.
PubMed - Narayanan, D.L., R.N. Saladi and J.L. Fox, 2010. Review: Ultraviolet radiation and skin cancer. Int. J. Dermatol., 49: 978-986.
CrossRefDirect Link - Nathan, C. and Q.W. Xie, 1994. Nitric oxide synthases: Roles, tolls and controls. Cell, 78: 915-918.
CrossRefDirect Link - Shimoda, H., J. Tanaka, S.J. Shan and T. Maoka, 2010. Anti-pigmentary activity of fucoxanthin and its influence on skin mRNA expression of melanogenic molecules. J. Pharm. Pharmacol., 62: 1137-1145.
CrossRef - St-Onge, M., N. Flamand, J. Biarc, S. Picard and L. Bouchard et al., 2007. Characterization of prostaglandin E2 generation through the cyclooxygenase (COX)-2 pathway in human neutrophils. Biochimica Biophys. Acta, 1771: 1235-1245.
CrossRefPubMedDirect Link - Tachibana, M., 2001. Cochlear melanocytes and MITF signaling. J. Invest. Dermatol. Symp. Proc., 6: 95-98.
CrossRefPubMedDirect Link - Terasaki, M., A. Hirose, B. Narayan, Y. Baba and C. Kawagoe et al., 2009. Evaluation of recoverable functional lipid components of several brown seaweeds (Phaeophyta) from Japan with special reference to fucoxanthin and fucosterol contents. J. Phycol., 45: 974-980.
CrossRef - Tomita, Y., K. Maeda and H. Tagami, 1992. Melanocyte-stimulating properties of arachidonic acid metabolites: Possible role in postinflammatory pigmentation. Pigment Cell Res., 5: 357-361.
CrossRefDirect Link - Ye, Y., J.H. Chu, H. Wang, H. Xu and G.X. Chou et al., 2010. Involvement of p38 MAPK signaling pathway in the anti-melanogenic effect of San-bai-tang, a Chinese herbal formula, in B16 cells. J. Ethnopharmacol., 132: 533-535.
CrossRef - Yoon, W.J., Y.M. Ham, B.S. Yoo, J.Y. Moon, J. Koh and C.G. Hyun, 2009. Oenothera laciniata inhibits lipopolysaccharide induced production of nitric oxide, prostaglandin E2 and proinflammatory cytokines in RAW264.7 macrophages. J. Biosci. Bioeng., 107: 429-438.
CrossRefPubMedDirect Link - Yoon, W.J., M.J. Kim, H.B. Koh, W.J. Lee, N.H. Lee and C.G. Hyun, 2010. Effect of korean red sea cucumber (Stichopus japonicus) on melanogenic protein expression in murine B16 melanoma. Int. J. Pharmacol., 6: 37-42.
CrossRefDirect Link - Zhang, X.Y., J.Y. Tao, L. Zhao, Z.J. Huang and F.L. Xiong et al., 2007. In vitro anti-inflammatory effects of different solution fractions of ethanol extract from Melilotus suaveolens Ledeb. Chinese Med. J., 120: 1992-1998.
PubMed