
M. Yaqoob Wani
Immunology Section, Izatnagar, Bareilly, Uttar Pradesh, 243122, India
Tapas Kumar Goswami
Immunology Section, Izatnagar, Bareilly, Uttar Pradesh, 243122, India
Raies Ahmad Mir
Division of Bacteriology and Mycology, Izatnagar, Bareilly, Uttar Pradesh, 243122, India
Pallab Chaudhuri
Division of Bacteriology and Mycology, Izatnagar, Bareilly, Uttar Pradesh, 243122, India
Kuldeep Dhama
Division of Pathology, Indian Veterinary Research Institute, Izatnagar, Bareilly, Uttar Pradesh, 243122, India
International Journal of Pharmacology
Year: 2013 | Volume: 9 | Issue: 8 | Page No.: 513-523







ABSTRACT
Gram negative sepsis and septic shock are among the leading causes of death, both in humans and animals. Statins, 3-hydroxy-3-methylglutaryl coenzyme (HMG-CoA) reductase inhibitors are said to have immune modulating effects. In the present study, it was hypothesized that amelioration of hyper immune activation by pravastatin can improve the immunoapthological status of acute sepsis. Pasteurella multocida Pm52 strain was used as a source of Lipopolysaccharide (LPS) and the pathogenic organism for induction of septicaemia in mice. In vitro trials showed that LPS extracted from P. multocida stimulated Nitric Oxide (NO) production in time and dose dependent manner in Mouse Embryonic Fibroblast (MEF) cultures. Addition of pravastatin to MEF culture supernatant significantly reduced Pm52 LPS induced NO production (p<0.05). In vivo studies showed that administration of pravastatin in combination with cefotaxime to P. multocida induced septicaemic mice significantly increased both mean survival time and survivability percentage compared to antibiotic and pravastatin treatments regimes. Furthermore, the serum TNF-α : IL-10 levels were significantly improved and near to normal healthy ratios in septicaemic mice treated with pravastatin+cefotaxime combination at 24 h post infection. Gross and histopathological findings revealed moderate lesion in pravastatin treated mice as compared to untreated and cefotaxime alone treated groups. The findings conclude that pravastatin stabilizes the immune compromised status of the septicaemic animals during early septic stages by stabilising the NO production, regulating the TNF-α: IL-10 ratio and reducing histopathological lesions. Although the mortality was not prevented, the immunopathological signs were ameliorated to a greater extent by this new treatment combination, further investigations are suggested to explore its possible therapeutic utility against sepsis and for septicaemic patients.
PDF Abstract XML References Citation
Received: January 06, 2014;
Accepted: February 19, 2014;
Published: May 13, 2014
How to cite this article
M. Yaqoob Wani, Tapas Kumar Goswami, Raies Ahmad Mir, Pallab Chaudhuri and Kuldeep Dhama, 2013. Moderation of Immunopathological Parameters by Pravastatin in Pasteurella multocida (Pm52) Induced Septicaemic Mice. International Journal of Pharmacology, 9: 513-523.
DOI: 10.3923/ijp.2013.513.523
URL: https://scialert.net/abstract/?doi=ijp.2013.513.523
DOI: 10.3923/ijp.2013.513.523
URL: https://scialert.net/abstract/?doi=ijp.2013.513.523
INTRODUCTION
Septicaemia is a systemic disease involving the presence of bacteria (bacteraemia) or their toxins like Lipopolysaccharide (LPS) (endotoxaemia) in the blood (Cohen, 2002; Van Amersfoort et al., 2003). The pathological process of sepsis is a disease continuum from infection, signs and symptoms of Infection/Systemic Inflammatory Response Syndrome (SIRS), sepsis, severe sepsis and Multiple Organ Dysfunction and Injury (MODI) (Liu and Malik, 2006). Human hospital mortality rate due to fatal sepsis remains very high between 30 and 50% worldwide even under intensive health care management (Cohen, 2002; Gao et al., 2008). Even in animals, sepsis and septic shock remained a leading cause of death from neonatal and immunocompromised animals, horses with colic and pleuropneumonia, bovines with displacement of abomasum, abomasal volvulus to uterine rupture during dystocia by causing acute peritonitis. Moreover, several bacterial diseases like haemorrhagic septicaemia, secondary bacterial complications to non bacterial epidemics, haemorrhage, trauma, burn or abdominal surgery all terminate with sepsis. The widespread activation of cells responsive to bacteria or bacterial components during sepsis results in the release of an array of inflammatory mediators, such as cytokines, chemokines, prostaglandins, lipid mediators and reactive oxygen species (Van Amersfoort et al., 2003; Remick, 2007). The key determinant of gram negative sepsis is LPS which is the fundamental structural component of Gram negative bacteria. Also, it has been found that LPS has been detected in majority of patients with sepsis, even independent of nature of infecting organism (Opal and Cohen, 1999). The standard model for the gram negative septic shock study is Caecal Ligation and Perforation model (CLP) (Wichterman et al., 1980; Buras et al., 2005). However, septic condition during CLP is influenced by many factors like intestinal contents at the time of perforation, size of perforating needle, surgical technique and the associated surgical strain and other complication. In this study it was hypothesized that a standard and constant number of septicaemia producing bacteria like Pasteurella multocida, when injected by intraperitoneal route can be used to produce consistent and well controlled acute septicaemic condition in mice. P. multocida is the causative agent of numerous economically important animal diseases like avian fowl cholera, bovine haemorrhagic septicaemia, enzootic pneumonia, swine atrophic rhinitis and snuffles’ (Manning et al., 1989; De Alwis, 1992; Ranjan et al., 2011; Shivachandra et al., 2011). Even, human infections have been observed in a range of sites by P. multocida, commonly following cat or dog bites (Weber et al., 1984). The key virulence factors of P. multocida identified till date include capsule and LPS (Harper et al., 2006; 2011). Being a Gram negative bacterium, LPS is the major structural constituent of cell wall of P. multocida and constitutes 1-10% of bacterial dry weight (Hodgson, 2006). Furthermore, it was found that intravenous inoculation of P. multocida LPS from serotype B:2 strain reproduce clinical signs of haemorrhagic septicaemia in buffalo (Horadagoda et al., 2002).
Since the discovery of penicillin in the mid twentieth century, antibiotics have been selectively used for bacterial disease prevention and control. Their wide spread use both at prophylactic and metaphylactic level has created a selective pressure towards emergence of resistant strains due to influences on biochemical and genetic pathways which in turn led to new pathologies (Ferber, 2000; Tiwari et al., 2013). The immunopathogenesis of sepsis, on another hand, indicates that the disease progresses through early immune hyper stimulation due to ‘cytokine storm’ which progresses towards immune paralytic or depression stages during terminal phases (Xiao et al., 2006; Remick, 2007). Therefore, a fine balance in the immune system can help to a great extent in averting the clinico-pathology of the septic disease consortium. Recent treatment approaches includes combination of different dug combinations like anti-inflammatory cytokine antibodies, Toll-like receptor (TLR)-4 antagonists, immunomodulatory drugs and others (Mekontso-Dessap and Brun-Buisson, 2006; Leon et al., 2008; Dhama et al., 2013a, b).
Statins are among the most effective drugs for lowering cardiovascular risk associated with elevation of Low Density Lipid (LDL) cholesterol (Brown and Goldstein, 1986; Jamroz-Wisniewska and Jerzy, 2005; Gao et al., 2008). Several lines of evidence suggest that the beneficial effects of statins are at least in part mediated by LDL independent (pleiotropic) mechanisms. These pleiotropic effects include anti-inflammatory actions, improvement of endothelial and microvascular functions and modulation of endothelial nitric oxide synthase (eNOS) (Lefer, 2002; Walter et al., 2004; Greenwood et al., 2006). Furthermore, both in vitro and in vivo studies have confirmed that statins decrease the acute inflammatory substances during LPS induced septic condition (Ando et al., 2000; Merx et al., 2005; McGown et al., 2010; Yeo et al., 2013). Therefore, in this study it was hypothesized that administration of an immunomodulatory drug in combination with an antibacterial can lead to a balance in the LPS induced immune stimulation thereby increasing the survivability of septicaemic mice. It was found that both the survivability percentage and survival duration of the septic mice was increased on administration of pravastatin and cefotaxime combination treatment. Although, the mortality was not prevented but the present study demonstrated that the immunopathological signs were ameliorated to a great extent by this new treatment combination; further investigations are suggested to explore its therapeutic potential against sepsis.
MATERIALS AND METHODS
Bacteria: Freeze dried ampoule of P. multocida strain 52 (Pm 52) of serotype B:2 used in the present study was procured from the Division of Bacteriology and Mycology, Indian Veterinary Research Institute (IVRI), Izatnagar. Culture was revived by reconstitution in 0.5 mL of Brain Heart Infusion (BHI) broth followed by streaking on sheep blood agar. Identification was done on the basis of morphology, biochemical characters and confirmed by amplification of serotype specific regions by Polymerase Chain Reaction (PCR) as described previously (Mir et al., 2012).
Experimental animals: Swiss albino mice of 6-8 weeks old weighing about 25-30 g were obtained from Laboratory Animal Resource (LAR) Section of IVRI, Izatnagar. Animals were used after acclimatization period of 4-5 days. All the experimental procedures on the animals were carried out according to the recommendations and approval of the Institute Animal Ethics Committee (IAEC) under the guidelines set forth by the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA).
Pathogenicity testing in mice: A single colony of Pm52 bacteria was inoculated into 5 mL of Brain Heart Infusion (BHI) broth and incubated for 18 h at 37°C as a shake culture. After overnight incubation, 0.2 mL of Pm52 culture was injected intraperitoneally into five healthy adult mice and observed for development of septicemia and mortality. The Pm52 bacteria were re-isolated from heart blood of the dead animals and were used for further studies.
Isolation, characterization and quantification of Pm52 LPS: Lipopolysaccharide (LPS) from the Pm52 bacterial cell wall was isolated by hot water/phenol procedure, characterized by silver staining method and quantified indirectly by measuring the carbohydrate content of the LPS as described previously (Wani et al., 2011).
Mouse Embryonic Fibroblasts (MEF) culture: The MEF cells were cultured as per the method described earlier (Lavnikova and Laskin, 1995) with some modifications. Embryos were collected from pregnant Swiss albino mice at 17-20 days of gestation after sacrificing the animal by standard ethical method. The extremities of embryos were removed, minced into small pieces and washed by Phosphate Buffered Saline (PBS), followed by washing with pre-warmed (37°C) 0.5% trypsin. The embryonic pieces were treated with 0.5% trypsin for 15 min to obtain single cell suspension. The resulting cell suspension was sieved, washed twice (400xg, 10 min) with PBS and finally with DMEM growth media containing 10% FCS. The mouse embryonic cells so obtained were incubated into tissue culture flasks in complete DMEM growth media (containing 10% FCS, 100 IU mL-1 penicillin, 100 μg mL-1 streptomycin) at 37°C and 5% CO2 for 72 h. Under these standard conditions only fibroblasts were expected to survive in the culture. The cells were subcultured 2-3 times to remove the non adherent cells and to keep exponential growth. The cell count and viability was determined by trypan blue dye exclusion method.
Effect of pravastatin on nitric oxide (NO) production by MEF culture: To determine the effect of pravastatin, 100 μL of fibroblast cells (2x106 cells mL-1) were plated in 96 well tissue culture plate (Nunc) in complete media supplemented with 5 μM L-arginine. Additional 100 μL of growth media containing Pm52 LPS was added to each well to achieve final concentration of 4 and 8 μg mL-1 LPS in the cell supernatant either alone or in combination with pravastatin (10 μM). Cells cultured without Pm52 LPS and/or pravastatin were kept as control. Each treatment was given to six wells in the culture plate. Cell culture supernatant was collected from the triplicate wells after 24 and 48 h post LPS treatment and stored at -20°C. The NO production by the LPS stimulated cells was done indirectly by measuring the nitrite (a stable metabolite of NO) levels as previously described (Tsai et al., 1999). Briefly, to 100 μL of the well supernatant, 100 μL of Griess reagent (equal volumes of 1% (w/v) sulphanilamide in 2.5% (v/v) H3PO4 and 0.1% (w/v) naphylethylenediamine HCl in 2.5% (v/v) H3PO4) was added. After incubation for 10 min at room temperature in darkness, absorbance was measures at 540 nm wavelength.
Viable bacterial counting: Viable bacterial count was performed by standard Miles-Misra colony counting method on blood agar plates (Quinn et al., 2001). Tenfold serial dilution of overnight bacterial broth culture was carried out using prewarmed PBS and 0.01 mL of 10-5 to 10-8 dilutions were placed on sectors of agar plate. Average colony forming units (CFU mL-1) were calculated by following Eq:
In vitro effect of drugs on bacterial growth: To determine the antibacterial effect and Minimum Inhibitory Concentration (MIC) of cefotaxime on Pm52, cefotaxime was added at concentrations of 0.5, 1, 2, 4, 8, 16 μg mL-1 as per the microtitre broth dilution method (Wiegand et al., 2008). About 200 μL of 18 h bacterial culture was added to each tube without dilution. Tubes were incubated for a period of 24 h at 37°C to determine the inhibition of Pm52 by antibiotic. The lowest concentration which inhibited the growth to minimum was assigned the MIC for cefotaxime. Effect of pravastatin on bacterial growth was also checked in a same way except that the final concentrations were 0.05, 0.1, 0.2, 0.4, 0.8 and 1.6 μg mL-1 in the culture broth medium.
In vivo experimental trial: Fifty mice were randomly and equally distributed into five different groups (n = 10 in each group). The first four different groups (1-4) were inoculated intraperitoneally with 0.2 mL of overnight bacterial culture inoculums containing 1.6x107 CFU mL-1 while as fifth one (5) was kept as control. Different treatment regimes for septicaemic animals were followed as described in Table 1. Mice in each group were closely observed for development of septicaemia.
| Table 1: | Treatment regime of septicaemic animals |
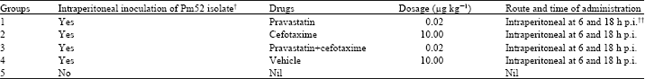 | |
| †Pm52 bacterial culture (0.2 mL of 1.6x107 CFU mL-1); ††p.i: Post inoculation of Pm52 bacteria | |
Animals were watched for their general behaviour (normal, dull or depressed), body condition, feed intake, body temperature and mortality patterns. The mortality/ survivability of experimentally infected mice were done in different treatment regimes. Survivability was checked at 6 h intervals after bacterial infection. Serum samples were collected at 24 h to determine the cytokine concentration and histopathological lesions were recorded in the representative tissues samples.
Serum cytokine assay: Blood from mice was collected from the retro orbital plexus at 24 h after the inoculation of bacteria in different groups. Serum was isolated from clotted blood and stored at -20°C until used. Serum TNF-α and IL-10 concentrations were determined by commercially available mice cytokine detection kits (Genetix Biotech) as per the manufacturers recommended protocol. Each serum sample was dilution (1:10) with PBS and tested in duplicates.
Histopathological evaluation: Animals from each group were randomly selected and euthanized by cervical dislocation at 24 and 48 h after bacterial inoculation. Parts of the lung, liver and spleen from the euthanized animals were preserved in 10% buffered neutral formalin and processed by routine paraffin embedding technique. Tissue section of 4-5 μm thickness were cut and stained with Haematoxylin and Eosin (HE) as per the conventional staining procedure (Bancroft and Cook, 1999). Effects of pravastatin and antibiotic treatments on cellular changes induced by the inflammatory responses to bacterial infection were assessed by assigning non-parametric scoring/severity index lesion. Severity index in the tissue sections was given based on the intensity of cellular changes as extremely severe (5), very severe (4), moderately severe (3), less severe (2) and least severe (1).
Statistical analysis: All data were analyzed with GraphPad Prism v4 software and results are expressed as Mean±SD. One way analyses of variance were used to test the statistical significance of parameters and values with p<0.05 where considered statistically significant.
RESULTS
Pathogenicity of Pm52 isolate in mice: Before the actual study the procured P. multocida Pm52 isolate was confirmed by morphological, biochemical characterization and molecular diagnostic methods. Expected amplicon sizes of P. multocida serotype B: 2 by PM-PCR and strain specific multiplex capsular PCR of ~460 and ~760 bp, respectively, were obtained (Fig. 1).
On inoculation to mice, Pm52 was found to be highly pathogenic and all the five mice died within 24 h by intraperitonial inoculation of 0.2 mL overnight grown culture. The bacteria were reisolated in pure culture from the heart blood of dead animals and were reused for further experimentations.
In vitro effects of antibiotic and pravastatin on bacterial growth: In order to determine the antibacterial effect and Minimum Inhibitory Concentration (MIC) of cefotaxime and pravastatin on Pm52 isolate, microtitre broth dilution method using BHI broth was used. The Pm52 isolate was found highly susceptible to cefotaxime antibiotic. The MIC of cefotaxime was found to be 1 μg mL-1 when the bacteria in BHI broth. Pravastatin was found to have no antibacterial effect even when at 1.6 μg mL-1 concentration in the culture medium.
Effect of pravastatin on NO production by fibroblasts: An incubation period of 2 h was given to cells for acclimatisation in the 96 well culture plates before stimulation with Pm52 extracted LPS. The MEF cultured cell (2 x105 cells in each well) were stimulated with 4 and 8 μg mL-1 of LPS either in presence or absence of pravastatin (10 μM). The NO release in the culture supernatant was measured indirectly by measuring the nitrite levels. The LPS was found to increase NO production in time and dose dependent manner (p<0.05). Average value of nitrite levels in the culture supernatant were 0.3±0.08 μM at 24 h and 1.46±0.33 μM at 48 h in 4 μg mL-1 stimulated cells. The levels were 1.58±0.24 μM at 24 h and 3.26±0.71 μM at 24 h in 8 μg mL-1 stimulated cells. These levels were significantly higher than the levels in respective control cells. Pravastatin was found to inhibit the NO production from LPS stimulated cells in a significant manner (p<0.05).
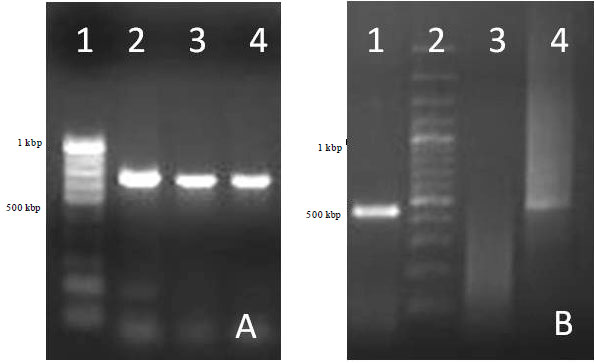 | |
| Fig. 1(a-d): | Confirmation of Pasteurella multocida serotype B: 2 (Pm52 isolate) by using strain B specific PCR and PM-PCR specific PCR |
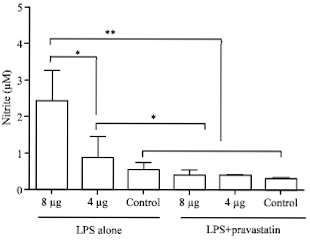 | |
| Fig. 2: | Effect of Pm52 extracted LPS on nitric oxide production by MEF (2x105 cells). The cells were stimulated with different concentrations of LPS either alone or in presence of pravastatin (10 μM) and NO production was estimated indirectly by measuring nitrite levels in culture supernatant (μM/2x105 cells). Values are represented as Mean±SD, **p< 0.01, *p< 0.05 |
Average nitrite levels in cell culture supernatant of 4 μg mL-1 LPs+pravastatin stimulated cells were 0.4±0.1 μM at 24 h and 0.39±0.14 μM at 48 h of incubation. The levels for 8 μg mL-1 LPs+pravastatin were 0.22±0.04 μM at 24 h and 0.54±0.26 μM at 48 h post stimulation (Fig. 2).
Effect of pravastatin on animal survivability: In the first step, a randomized experimental trial was carried out to determine the number of bacteria to induce a consistent and acute clinical septicaemic condition. Tenfold serially diluted bacterial samples of 0.2 mL from 1.6x109 to 105 CFU mL-1 were inoculated into five groups with 5 mice in each group. It was found that inoculation of mice with of 0.2 mL of bacterial culture containing 1.6x107 CFU mL-1 induces consistent septicaemic condition. Most of the septicaemic mice died during acute phase and does not survived beyond 36 h post bacterial inoculation. In the second step, pravastatin was used either alone or in combination with cefotaxime to determine its effect on mortality/survivability in septicaemic mice. The mortality patterns were different under different treatment regimes. Mean survival time was longer in antibiotic treated group (42 h) and cefotaxime+pravastatin treated group (48 h) as compared to pravastatin alone treated group (20 h) and untreated septicaemic group (20 h), respectively. The survivability percentages were significantly different and were highest cefotaxime+pravastatin treated group (3/10; 30%) and antibiotic treated groups (2/10; 20%). None of the animals survived in untreated and pravastatin alone treated groups at the end of the experimental study (72 h) while all remained healthy in the non-infected control group (Fig. 3).
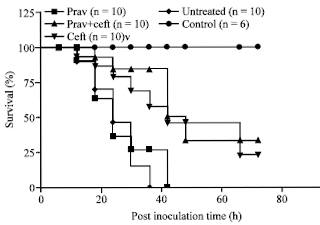 | |
| Fig. 3: | Effect of different treatment regimes on the survivability/mortality patterns of septicaemic mice |
Serum cytokine levels during septicaemia: Effect of the pravastatin on serum TNF-α and IL-10 levels was assessed at 24 h post infection in septicaemic mice treated with different treatment regimes. The normal serum TNF-α and IL-10 levels in mice were found to be 324.02±13 and 658.52±109.29 pg mL-1. At 24 h post Pm52 inoculation, serum TNF-α level were undetectable in untreated septicaemic mice and significantly higher in pravastatin alone treated group (1057.10±447.73 pg mL-1). On the other hand, serum IL-10 levels were significantly higher in untreated septicaemic mice (1929.92± 704.45 pg mL-1) and lower in antibiotic alone treated group (124.96±101.95) as compared to control values. Mice treated with either pravastatin alone or in combination with antibiotic were having serum IL-10 levels within the normal range. The ratio of serum TNF-α: IL-10 cytokine levels at 24 h post Pm52 inoculation are also presented (Fig. 4). The normal TNF-α: IL-10 ration in healthy mice was found to be 0.49 and was significantly different from that of pravastatin alone treated mice (2.09). Mice treated with cefotaxime were having ratios in their normal range.
Histopathology: Histopathology of lung, liver and spleen samples collected from the septicaemic mice was carried out at 24 post infection to determine the effect of antibiotic and pravastatin on immune mediated cellular changes. Slight to moderate differences with respect to inflammatory changes were observed in different groups. Histopathological changes with regard to infiltration of polymorphonuclear cells, presence of inflammatory exudate and haemorrhages, haemosiderosis, hydropic degeneration and fatty changes in hepatocytes, thickening of alveolar septa and disruption of alveolar lumen in lungs, atrophy of red and white pulp in spleen were recorded (Fig. 5).
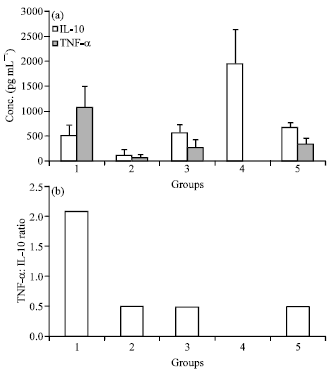 | |
| Fig. 4(a-b): | Effect of different treatment regimes on the (a) Serum TNFα and IL-10 cytokine levels and (b) Ratio of TNFα: IL-10 in the serum. The mice were treated either with pravastatin alone (Group 1), cefotaxime alone (Group 2) or combination of cefotaxime+pravastatin (Group 3). The positive control as untreated septicaemic mice (Group 4) and negative control as healthy mice (Group 5) were kept for comparison |
The intensities of pathological changes were significantly different in different treatment groups. The lesions were more pronounced in untreated septicaemic mice and were least in cefotaxime+pravastatin treated mice (Fig. 6).
DISCUSSION
Conventional models used for the study of immune-pathogenesis of sepsis assume that microorganism or their products are necessarily injurious to health. However, an evolutionary perspective suggests that host-microbial interactions are symbiotic in nature and disease results only after the disruption of mutually beneficial homeostatic state (Marshall, 2005). Due to its intimate association with sepsis, LPS is the target for development of novel and emerging therapies (Leaver et al., 2007; Leon et al., 2008). In this regard, the present study was proposed on the hypothesis that amelioration of the exaggerated immune responses during early stages of LPS induced septic shock by pravastatin can help the host in a better way.
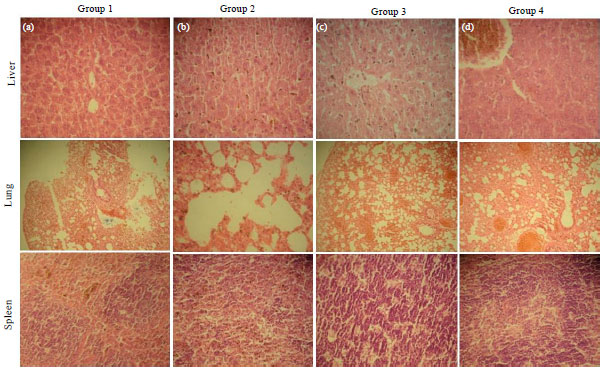 | |
| Fig. 5(a-b): | Histopathological changes in various tissues of the mice inoculated with P. multocida Pm52 isolate to induce septicemia. The effect of pravastatin (Group 1), cefotaxime (Group 2) and their combination (Group 3) was compared with infected control (Group 4) at 24 h. Mice were randomly selected and sacrificed before the death in each group. The vascular changes and degeneration of parenchyma cells in different organs were compared |
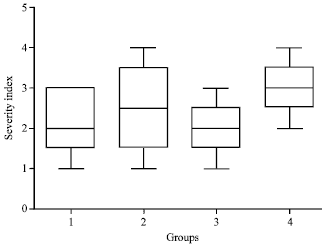 | |
| Fig. 6: | Box plots representation of histopathological lesions recorded as severity index in pravastatin (Group 1), cefotaxime (Group 2) cefotaxime+pravastatin (Group 3) and untreated group (Group 4) at 24 h post infection. Lesion scoring was done as extreme severe (5), very severe (4), moderately severe (3), less severe (2) and least severe (1) |
Nitric oxide is one of the central molecules in the immunopathogenesis of sepsis (Parratt, 1998). At lower concentrations, NO exhibits wider range of physiological functional from inhibition of platelet aggregation, neurotransmission, vasodilatation to regulation of cell death while higher concentration it leads to endothelial cell damage, disseminated intravascular coagulation and cell cytotoxicity (Tripathi, 2007). Although, macrophages produce higher levels of NO during inflammation, fibroblasts are of particular interest because they outnumber the tissue macrophages (Pinkerton et al., 1982; Nathan, 1992). In the present study, Pm52 derived LPS was found to increase NO production both in fibroblast cultures both in a time and concentration dependent manner (p<0.05). Additionally, previous reports have shown that that LPS has synergistic effects with various cytokines (IFNγ, IL-1, TNF-α and IL-1β) on NO production by MEF and rat lung fibroblasts (Lavnikova and Laskin, 1995). Therefore, the actual amount of NO produced by fibroblasts during LPS stimulation would be much higher in the host during acute inflammatory conditions.
Addition of pravastatin was found to inhibit LPS induced NO production from MEF significantly when used at the concentration of 10 μM. This concentration (10 μM) was selected to make it comparable with the plasma levels which are achieved at therapeutic dosage during treatment (Desager and Horsmans, 1996). The results of the present study were in agreement with the previous reports indicating inhibitory effect of NO production by simvastatin and pravastatin on cardiac embryonic myoblasts and RAW 264 cell lines, respectively (Madonna et al., 2005; Abe et al., 2008). One of the interesting observation during present study was that pravastatin inhibited NO production only to basal levels, indicating its inhibitory effects on exaggeratedly stimulated fibroblasts. The possible explanation for this low production of NO by LPS stimulated cells in presence of pravastatin may be due to selective inhibition of Inducible Nitric Oxide Synthase (iNOS) (Wagner et al., 2002).
In the present study, P. multocida type B (Pm52) was inoculated through intra-peritoneal route in Swiss albino mice to induce septicaemia. This combination would provide an ideal tool to study septicaemia due to cost and suitable animal models. It was found that 0.2 mL bacterial culture containing 1.6x107 CFU mL-1 induces acute sepsis in mice and all the mice died within 36 h post bacterial inoculation. In order to determine the effect of pravastatin during septicaemic condition, different treatment regimes were followed along with using cefotaxime, a third generation cephalosporin. The therapeutic dose for cefotaxime was selected as 10 times the in vitro MIC value. This is due to the fact that in general clinical practices doses are selected to achieve 4-10 times higher MIC concentration in the serum (Periti and Mazzei, 1999). Treatment of septicaemic mice with cefotaxime either alone or in combination with pravastatin was found to significantly increase the mean survival time and survivability (p< 0.05). Mean survival time was longer in cefotaxime+pravastatin treated group (48 h) and cefotaxime treated group (42 h) as compared to pravastatin treated and untreated septicaemic mice (20 h), respectively. The survivability percentages were significantly different and were highest for cefotaxime+pravastatin treated group (3/ 10; 30%) and antibiotic treated groups (2/10; 20%) compared to untreated and pravastatin alone treated groups.
Antibacterial are generally used during sepsis and septicaemic conditions. However, antibiotics do not prevent the overall mortality even when appropriate care is provided to septic patients. In the present study, cefotaxime and pravastatin were used for treating septic mice. During in vitro studies it was found that addition of pravastatin to the Pm52 bacterial culture does not have any effect antibacterial effect even when added at therapeutic doses. Recent reports have also indicated that statins does not have antibacterial effects (Farmer et al., 2013). Further, earlier reports indicated that cefotaxime bind to PBP3 leading to formation of long filamentous and multiseptate bacterial biomasses which act as endotoxin accumulators (Buijs et al., 2008). The endotoxin is non toxic when incorporated into the Gram negative bacterial outer membrane, but its release due to bacterial death and lysis caused by complement activation, bactericidal proteins or exposure to bactericidal antibiotics (Crosby et al., 1994; Smedsrod et al., 1994; Hellman et al., 2000). Release of endotoxin from bacteria causes its toxic moiety, lipid A to be exposed to immune cells thereby evoking acute inflammatory responses.
The endotoxic shock, a consequence of LPS mediated hyper immune activation, is known to be mediated by TNF-α and alleviated physiologically by IL-10 (Howard et al., 1993; Durez et al., 1999). Therefore, animal models and experimental designs used to study TNF-α: IL-10 ratio may be suitable for the screening of compounds used for the treatment during septic shock and septicaemic conditions. In present experimental design the effect of pravastatin on serum levels of TNF-α and IL-10 were estimated at 24 h post infection to determine the immune status of the septicaemic mice. It was found that serum TNF-α levels in untreated septicaemic mice were undetectable and that of IL-10 were highest (1929.917±704.452) at 24 h post infection. Decrease in TNF-α levels and increase in IL-10 serum concentrations indicates the immune paralysed status of septic mice. Immune paralysed status usually occurs towards the terminal stages of septic conditions (Xiao et al., 2006). Mice treated with either pravastatin alone or in combination with cefotaxime were having serum IL-10 levels within the normal range. Analysis of data further indicated that the ratio of TNF-α: IL-10 cytokines was highest for pravastatin treated group (2.09) as compared to untreated group (not detected) which may be due to its delay in cytokine release. This delay in the cytokine release would have beneficial effect on the host by balancing the immune status and is the possible explanation of increased mean survival time and survivability of septicaemic condition.
Histopathology of lungs, liver and spleen indicated almost similar changes at 24 h post infection in all the septicaemic mice treated with different regimes. This may be due to trapping of bacteria and the active inflammatory reaction that have occurred in these organs. In visceral organs especially in the liver, macrophages (Kupffer cells) constitutively transcribe TNF-α leading to its rapid release during inflammation (Smedsrod et al., 1994). Also, alveolar macrophages and lung fibroblasts are regarded as the active sources of NO production during inflammation (Lavnikova et al., 1993). All these factors are responsible for increased severity in visceral organs during sepsis and septicaemic conditions. However, on the basis of intensity of histopathological lesion, measures as severity index in each group indicated that mice treated with pravastatin either alone or in combination with cefotaxime showed less severe lesions compared to untreated septicaemic mice. The possible explanation for minimal cellular changes in pravastatin treated groups may be due to inhibited of both TNF-α and iNOS release and cefotaxime may have synergistically prevented infection before the exaggerated tissue damage. Due to this only the mice from groups treated with cefotaxime and pravastatin+cefotaxime crossed the 48 h post infection intervals.
CONCLUSION
Taken together, the present study demonstrates for the first time the effect of pravastatin and cefotaxime combinations on the immunopathological parameters during P. multocida induced sepsis. It was found that pravastatin stabilized the immunocompromised status of the septicaemic animals by stabilising the NO production and regulating the TNF-α: IL-10 ratios. Significant improvements in the mean survival time, survivability percentage and less severe histopathological signs were found by this new treatment regime. Treatment regimes based on the immunomodulation during early stages of sepsis and septicaemic conditions would thus provide an increased time window for the treatment of septicaemic patients. However, further investigations are required in this direction to provide clarifications regarding the molecular mechanisms, drug interactions and the long term effects of such regimes.
ACKNOWLEDGMENT
The authors are thankful to the Director, Joint Director (Academic) and Scientific Coordinator of IVRI, Izatnagar, for monetary and material support.
REFERENCES
- Ando, H., T. Takamura, T. Ota, Y. Nagai and K.I. Kobayashi, 2000. Cerivastatin improves survival of mice with lipopolysaccharide-induced sepsis. J. Pharmacol. Exp. Ther., 294: 1043-1046.
Direct Link - Bancroft, J.D. and H.C. Cook, 1999. Manual of Histological Techniques and their Diagnostic Applications. Churchill Livingstone, New York.
Direct Link - Brown, M.S. and J.L. Goldstein, 1986. A receptor-mediated pathway for cholesterol homeostasis. Science, 232: 34-47.
CrossRefDirect Link - Buijs, J., A.S. Dofferhoff, J.W. Mouton, J.H. Wagenvoort and J.W. van der Meer, 2008. Concentration-dependency of β-lactam-induced filament formation in Gram-negative bacteria. Clin. Microbiol. Infect., 14: 344-349.
CrossRef - Buras, J.A., B. Holzmann and M. Sitkovsky, 2005. Animal models of sepsis: Setting the stage. Nat. Rev. Drug Discovery, 4: 854-865.
CrossRef - Crosby, H.A., J.F. Bion, C.W. Penn and T.S.J. Elliott, 1994. Antibiotic-induced release of endotoxin from bacteria in vitro. J. Med. Microbiol., 40: 23-30.
CrossRef - De Alwis, M.C.L., 1992. Pasteurellosis in Production Animals: A Review. In: Pasteurellosis in Production Animals: An International Workshop Sponsored, Patten, B.E., T.L. Spencer, R.B. Johnson, D. Hoffman and L. Lehane (Eds.). Australian Centre for International Agricultural Research, Bali, Indonesia, ISBN: 9781863200813, pp: 11-18.
- Smedsrod, B., P.J. de Bleser, F. Braet, P. Lovisetti, K. Vanderkerken, E. Wisse and A. Geerts, 1994. Cell biology of liver endothelial and Kupffer cells. Gut, 35: 1509-1516.
Direct Link - Desager, J.P. and Y. Horsmans, 1996. Clinical pharmacokinetics of 3-hydroxy-3-methylglutaryl-coenzyme A reductase inhibitors. Clin. Pharmacokinetics, 31: 348-371.
CrossRef - Dhama, K., S. Chakraborty, Mahima, M.Y. Wani and A.K. Verma et al., 2013. Novel and emerging therapies safeguarding health of humans and their companion animals: A review. Pak. J. Biol. Sci., 16: 101-111.
CrossRefDirect Link - Dhama, K., S. Chakraborty, M.Y. Wani, R. Tiwari and R. Barathidasan, 2013. Cytokine therapy for combating animal and human diseases-A review. Res. Opin. Anim. Vet. Sci., 3: 195-208.
Direct Link - Durez, P., T. Appelboom, C. Pira, P. Stordeur, B. Vray and M. Goldman, 1999. Antiinflammatory properties of mycophenolate mofetil in murine endotoxemia: Inhibition of TNF-α and upregulation of IL-10 release. Int. J. Immunopharmacol., 21: 581-587.
CrossRefDirect Link - Farmer, A.R., C.K. Murray, K. Mende, K.S. Akers, W.C. Zera, M.L. Beckius and H.C. Yun, 2013. Effect of HMG-CoA reductase inhibitors on antimicrobial susceptibilities for Gram‐Negative rods. J. Basic Microbiol., 53: 336-339.
CrossRef - Gao, F., L. Linhartova, A.M. Johnston and D.R. Thickett, 2008. Statins and sepsis. Br. J. Anaesthesia, 100: 288-298.
CrossRefDirect Link - Greenwood, J., L. Steinman and S.S. Zamvil, 2006. Statin therapy and autoimmune disease: From protein prenylation to immunomodulation. Nat. Rev. Immunol., 6: 358-370.
CrossRefDirect Link - Harper, M., A.D. Cox, B. Adler and J.D. Boyce, 2011. Pasteurella multocida lipopolysaccharide: The long and the short of it. Vet. Microbiol., 153: 109-115.
CrossRef - Harper, M., J.D. Boyce and B. Adler, 2006. Pasteurella multocida pathogenesis: 125 years after Pasteur. F.E.M.S. Microbiol. Lett., 265: 1-10.
CrossRefDirect Link - Hellman, J., P.M. Loiselle, M.M. Tehan, J.E. Allaire and L.A. Boyle et al., 2000. Outer membrane protein A, peptidoglycan-associated lipoprotein and murein lipoprotein are released by Escherichia coli bacteria into serum. Infect. Immun., 68: 2566-2572.
CrossRef - Hodgson, J.C., 2006. Endotoxin and mammalian host responses during experimental disease. J. Comp. Pathol., 135: 157-175.
CrossRefDirect Link - Horadagoda, N.U., J.C. Hodgson, G.M. Moon, T.G. Wijewardana and P.D. Eckersall, 2002. Development of a clinical syndrome resembling haemorrhagic septicaemia in the buffalo following intravenous inoculation of Pasteurella multocida serotype B:2 endotoxin and the role of tumour necrosis factor-α. Res. Vet. Sci., 72: 194-200.
CrossRefPubMed - Howard, M., T. Muchamuel, S. Andrade and S. Menon, 1993. Interleukin 10 protects mice from lethal endotoxemia. J. Exp. Med., 177: 1205-1208.
CrossRefPubMedDirect Link - Jamroz-Wisniewska, A. and J. Bettowski, 2005. Adverse effects of statins. Int. J. Pharmacol., 1: 210-225.
CrossRefDirect Link - Lavnikova, N., J.C. Drapier and D.L. Laskin, 1993. A single exogenous stimulus activates resident rat macrophages for nitric oxide production and tumor cytotoxicity. J. Leukocyte Biol., 54: 322-328.
Direct Link - Lavnikova, N. and D.L. Laskin, 1995. Unique patterns of regulation of nitric oxide production in fibroblasts. J. Leukocyte Biol., 58: 451-458.
Direct Link - Leaver, S.K., S.J. Finney, A. Burke-Gaffney and T.W. Evans, 2007. Sepsis since the discovery of Toll-like receptors: Disease concepts and therapeutic opportunities. Crit. Care Med., 35: 1404-1410.
CrossRef - Leon, C.G., R. Tory, J. Jia, O. Sivak and K.M. Wasan, 2008. Discovery and development of Toll-Like Receptor 4 (TLR4) antagonists: A new paradigm for treating sepsis and other diseases. Pharmaceut. Res., 25: 1751-1761.
CrossRefDirect Link - Liu, S.F. and A.B. Malik, 2006. NF-κB activation as a pathological mechanism of septic shock and inflammation. Am. J. Physiol. Lung Cell. Mol. Physiol., 290: L622-L645.
Direct Link - Madonna, R., P. di Napoli, M. Massaro, A. Grilli and M. Felaco et al., 2005. Simvastatin attenuates expression of cytokine-inducible nitric-oxide synthase in embryonic cardiac myoblasts. J. Biol. Chem., 280: 13503-13511.
CrossRefPubMedDirect Link - Marshall, J.C., 2005. Lipopolysaccharide: An endotoxin or an exogenous hormone? Clin. Infect. Dis., 41: S470-S480.
CrossRefDirect Link - McGown, C.C., N.J. Brown, P.G. Hellewell, C.S. Reilly and Z.L.S. Brookes, 2010. Beneficial microvascular and anti-inflammatory effects of pravastatin during sepsis involve nitric oxide synthase III. Br. J. Anaesthesia, 104: 183-190.
CrossRefDirect Link - Mekontso-Dessap, A. and C. Brun-Buisson, 2006. Statins: The next step in adjuvant therapy for sepsis? Intensive Care Med., 32: 11-14.
CrossRef - Merx, M.W., E.A. Liehn, J. Graf, A. van de Sandt and M. Schaltenbrand et al., 2005. Statin treatment after onset of sepsis in a murine model improves survival. Circulation, 112: 117-124.
CrossRef - Nathan, C., 1992. Nitric oxide as a secretory product of mammalian cells. FASEB J., 6: 3051-3064.
Direct Link - Opal, S.M. and J. Cohen, 1999. Clinical gram-positive sepsis: Does it fundamentally differ from gram-negative bacterial sepsis? Crit. Care Med., 27: 1608-1616.
Direct Link - Parratt, J.R., 1998. Nitric oxide in sepsis and endotoxaemia. J. Antimicrob. Chemother., 41: 31-39.
CrossRef - Periti, P. and T. Mazzei, 1999. New criteria for selecting the proper antimicrobial chemotherapy for severe sepsis and septic shock. Int. J. Antimicrob. Agents, 12: 97-105.
CrossRef - Pinkerton, K.E., B.E. Barry, J.J. O'Neil, J.A. Raub, P.C. Pratt and J.D. Crapo, 1982. Morphologic changes in the lung during the lifespan of Fischer 344 rats. Am. J. Anatomy, 164: 155-174.
CrossRef - Ranjan, R., S.K. Panda, A.P. Acharya, A.P. Singh and M.K. Gupta, 2011. Molecular diagnosis of haemorrhagic septicaemia-A review. Vet. World, 4: 189-192.
Direct Link - Shivachandra, S.B., K.N. Viswas and A.A. Kumar, 2011. A review of hemorrhagic septicemia in cattle and buffalo. Anim. Health Res. Rev., 12: 67-82.
CrossRefDirect Link - Tiwari, R., S. Chakraborty, K. Dhama, S. Rajagunalan and S.V. Singh, 2013. Antibiotic resistance-an emerging health problem: Causes, worries, challenges and solutions: A review. Int. J. Curr. Res., 5: 1880-1892.
Direct Link - Tripathi, P., 2007. Nitric oxide and immune response. Indian J. Biochem. Biophys., 44: 310-319.
Direct Link - Tsai, S.H., S.Y. Lin-Shiau and J.K. Lin, 1999. Suppression of nitric oxide synthase and the down-regulation of the activation of NFκB in macrophages by resveratrol. Br. J. Pharmacol., 126: 673-680.
CrossRefDirect Link - van Amersfoort, E.S., T.J.C. van Berkel and J. Kuiper, 2003. Receptors, mediators and mechanisms involved in bacterial sepsis and septic shock. Clin. Microbiol. Rev., 16: 379-414.
CrossRefDirect Link - Wagner, A.H., O. Schwabe and M. Hecker, 2002. Atorvastatin inhibition of cytokine-inducible nitric oxide synthase expression in native endothelial cells in situ. Br. J. Pharmacol., 136: 143-149.
CrossRef - Walter, D.H., S. Dimmeler and A.M. Zeiher, 2004. Effects of statins on endothelium and endothelial progenitor cell recruitment. Semin. Vascular Med., 4: 385-393.
CrossRef - Wani, M.Y., T.K. Goswami, R.A. Mir, D. Chaudhary and G.C. Ram, 2011. Isolation, quantification and characterization of Pasteurella multocida Pm52 derived lipopolysaccharaide and its effect on nitric oxide production in mice. J. Immunol. Immunopathol., 13: 75-82.
Direct Link - Weber, D.J., J.S. Wolfson, M.N. Swartz and D.C. Hooper, 1984. Pasteurella multocida infections: Report of 34 cases and review of the literature. Medicine, 63: 133-151.
Direct Link - Wichterman, K.A., A.E. Baue and I.H. Chandry, 1980. Sepsis and septic shock-A review of laboratory models and a proposal. J. Surg. Res., 29: 189-201.
CrossRefPubMedDirect Link - Wiegand, I., K. Hilpert and R.E.W. Hancock, 2008. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc., 3: 163-175.
CrossRefDirect Link - Xiao, H., J. Siddiqui and D.G. Remick, 2006. Mechanisms of mortality in early and late sepsis. Infect. Immun., 74: 5227-5235.
CrossRefDirect Link - Yeo, C.D., C.K. Rhee, I.K. Kim, H.H. Kang and S.H. Lee et al., 2013. Protective effect of pravastatin on lipopolysaccharide-induced acute lung injury during neutropenia recovery in mice. Exp. Lung Res., 39: 99-106.
CrossRef